
Figure 4. Euglycemic hyperinsulinemic clamp.
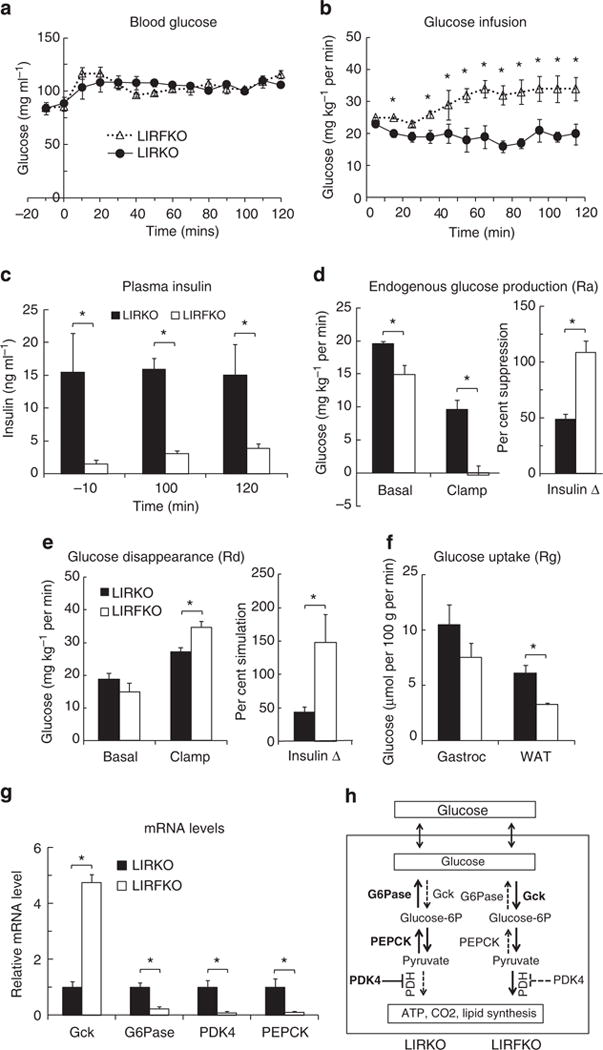
Circulating glucose levels (a), glucose infusion rates (b), insulin levels (c), endogenous glucose production (Ra) (d), total body glucose turnover rate (Rd) (e), glucose uptake (Rg) by gastrocnemius (gastroc) and white adipose tissue (WAT)(f), and hepatic gene expression at the end of the insulin clamp (g) in LIRKO (N = 7) and LIRFKO (N = 6) mice are shown as the mean and s.e.m. *P<0.05 LIRKO versus LIRFKO by two-tailed unpaired Student’s t-test. (h) Hepatic glucose metabolism in LIRKO versus LIRFKO mice. Glucose is readily transported in and out of hepatocytes. In LIRFKO mice, glucose is phosphorylated by glucokinase (Gck) to form glucose-6 phosphate, which can be metabolized to form pyruvate, and then utilized for energy metabolism through the effect of pyruvate dehydrogense (PDH). Glucose utilization is limited in LIRKO liver due to reduced expression of Gck, increased expression of glucose-6 phosphatase (G6Pase), which dephosphorylates glucose-6 phosphate and promotes release of glucose by the liver, and increased expression of PDK4, which inhibits the ability to use pyruvate for energy metabolism. Pyruvate also can serve as a substrate for gluconeogenesis, and increased expression of PEPCK and the availability of pyruvate can both promote increased glucose production in LIRKO mice. In LIRFKO mice, glucose production is limited by reduced PEPCK expression and because pyruvate is able to be used for other purposes.