
Figure 1.
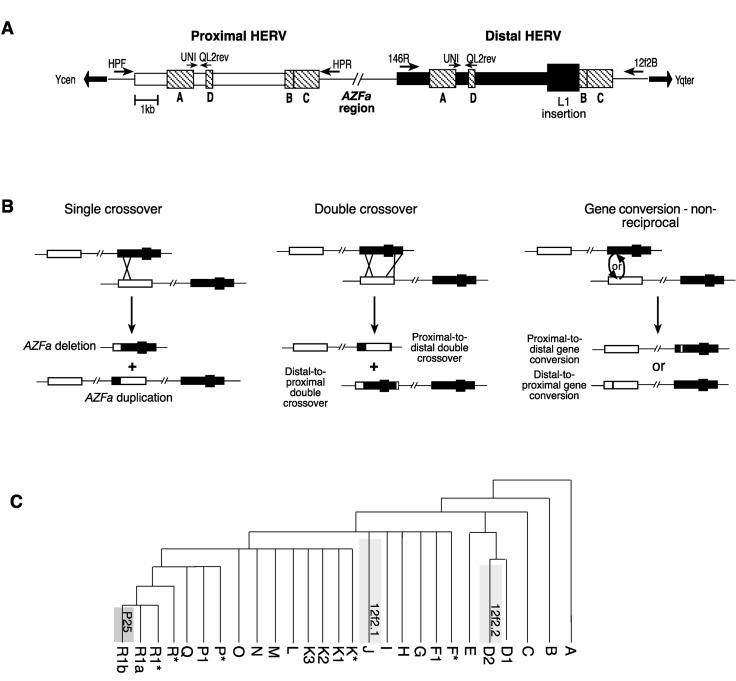
HERV structures, HERV-sponsored rearrangements, and the phylogenetic positions of known double-crossover and gene conversion events. (A) Structures of the proximal and distal AZFa HERVs, showing blocks of identity >200 bp (hatched boxes) and the L1 fragment insertion in the distal HERV. Identity blocks A through D are named according to Bosch and Jobling (2003). Flanking specific primers (large arrows) and primers used for the amplification of the region between identity blocks A and D (small arrows) are shown. (B) Possible sequence rearrangements sponsored by the HERVs: Single meiotic crossover between misaligned sister chromatids (in the same orientation) causes deletion or duplication of the AZFa region, both of which have been observed; double crossover causes the transfer of a large segment of one HERV to the other, observed in the proximal-to-distal case only; and gene conversion causes the nonreciprocal transfer of small segments, observed in both polarities. (C) Phylogeny of the Y chromosome showing known double-crossover and gene conversion events. Two examples of proximal-to-distal transfer through double-crossover are known: The concomitant loss of the L1 segment from the distal HERV is typed as a binary marker (Casanova et al. 1985) and defines haplogroup J (12f2.1). The second occurrence (12f2.2) is phylogenetically equivalent to binary markers defining haplogroup D2 (Blanco et al. 2000). A distal-to-proximal conversion of between 36 and 72 bp is phylogenetically equivalent to the marker P25, defining haplogroup R1b; one example of a chromosome with a proximal-to-distal conversion within this haplogroup is also known (Bosch and Jobling 2003).