
Figure 1. Localization of 3‐phosphoinositides in the cell, and their relevant kinases and phosphatases.
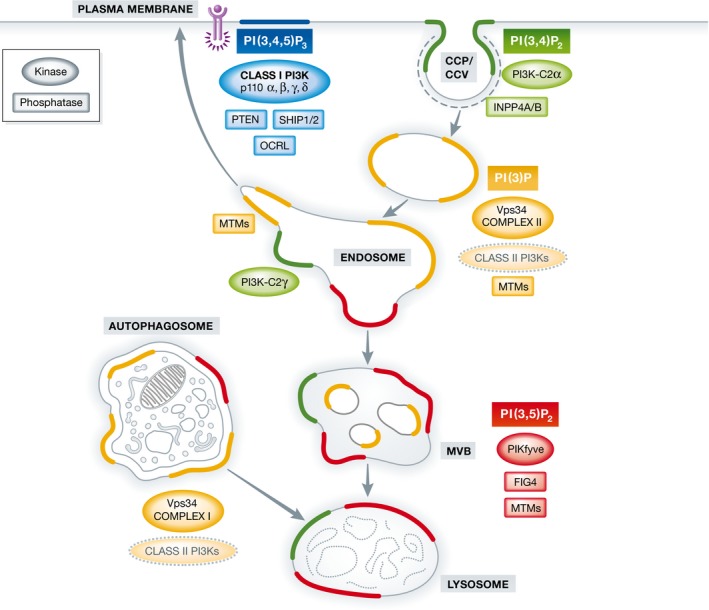
At the plasma membrane, receptor tyrosine kinase (purple) activation results in the production of PI(3,4,5)P3 via class I PI3Ks (with α, β, γ, and δ isoforms). PI(3,4,5)P3 is a substrate for the PI 3‐phosphatase PTEN and for the PI 5‐phosphatases SHIP1/2 and OCRL. Class II PI3Kα generates a plasma membrane pool of PI(3,4)P2 necessary for clathrin‐coated pit (CCP) maturation and formation of free clathrin‐coated vesicles (CCV) during endocytosis. PI(3)P is a defining and essential feature of early endosomes and is generated primarily by the class III PI3K Vps34 complex II with a possible contribution of class II PI3Ks (encircled by dashed line), either by direct PI(3)P synthesis or indirectly via PI(3,4)P2 hydrolysis by the PI 4‐phosphatases INPP4A/B. As endosomes mature into late endosomes/multivesicular bodies (MVBs), PI(3)P is converted into PI(3,5)P2 via the PI(3)P 5‐kinase PIKfyve. Endosomes and lysosomes also harbour a poorly understood pool of PI(3,4)P2. In the liver, this endosomal PI(3,4)P2 pool is synthesized by class II PI3Kγ. Autophagosomes contain PI(3)P produced by the autophagosome‐specific Vps34 complex I, and possibly by the class II PI3Ks (dashed line). Endosomal recycling to the cell surface requires PI(3)P hydrolysis by myotubularin phosphatases (MTMs) such as MTM1 and the concomitant generation of PI(4)P (not shown) to enable exocytosis. PI(3,5)P2 turnover at MVBs and/or lysosomes is mediated by MTMs together with the PI(3,5)P2 5‐phosphatase Fig4.