
Fig. 2.
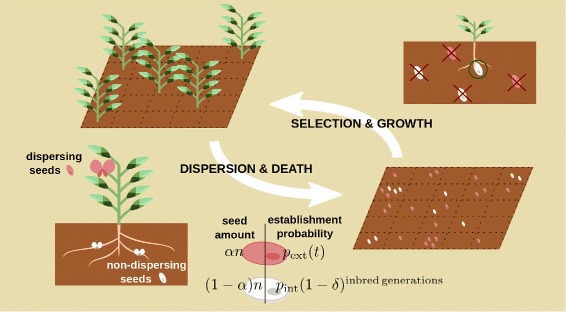
Community of plants with mixed dispersal phenotypes. Each plant is located at a cell of a square lattice of size L×L. Each individual plant produces the same fixed number of seeds, n; seeds can be of two different types: “dispersing” and “non-dispersing”, marked in red and white respectively in the sketch. Each produced seed is external/dispersing with probability α, or internal/non-dispersing with probability, 1−α, where α is the dispersal propensity parameter. After reproduction, all adult plants are removed from the community. Regardless of their origin, dispersing seeds can randomly arrive to any cell in the lattice where they get established with probability p ext(t) which is environment-dependent. On the other hand, non-dispersing seeds can only establish themselves at the maternal location or in its adjacent lattice cells. Then, for cells with more than one established seed, one of these is chosen at random and the rest die. The establishment probability p int of non-dispersing seeds is assumed to be independent from environmental variability (and thus, it does not depends on time). In the simplest case, dispersing seeds are produced by outcrossing, whereas non-dispersing seeds are the product of selfing. Thus, their quality, q, is reduced after each inbreeding event by a penalization factor q→(1−δ)q. In our formulation, inbreeding depression is approximated in a manner that can be assimilated to the interaction among many slightly deleterious alleles that affect the trait independently (e.g., [36]). The number of alleles determining inbreeding is assumed to be n→∞. Therefore, it is always proportional to homozigosity and accumulates multiplicatively with inbreeding events. Outcrossing events are expected to eliminate homozigosity and to reset δ=0 and q=1, so dispersing seeds are always assumed to not have any inbreeding depression. We also study a more general scenario in which selfing is not restricted to a specific dispersal syndrome, but in which inbreeding affects the quality of seeds produced by individuals that mate with relatives (see Additional file 1: S1). In this case, inbreeding is proportional to the kinship between mating partners. This generalized model is less restrictive in that there is no assumption of a perfect association between dispersal and mating, but leads to qualitatively similar conclusions