

ABSTRACT
The putative transfer and gene regulatory activities of diet-derived miRNAs in ingesting animals are still debated. Importantly, no study to date has fully examined the role of dietary uptake of miRNA in the honey bee, a critical pollinator in both agricultural and natural ecosystems. After controlled pollen feeding experiments in adult honey bees, we observed that midguts demonstrated robust increases in plant miRNAs after pollen ingestion. However, we found no evidence of biologically relevant delivery of these molecules to proximal or distal tissues of recipient honey bees. Our results, therefore, support the premise that pollen miRNAs ingested as part of a typical diet are not robustly transferred across barrier epithelia of adult honey bees under normal conditions. Key future questions include whether other small RNA species in honey bee diets behave similarly and whether more specialized and specific delivery mechanisms exist for more efficient transport, particularly in the context of stressed barrier epithelia.
KEYWORDS: Biotechnology, cross-kingdom delivery, ecology, genetically engineered, honey bee, microRNA, pollen, pollinator
Abbreviations
- dsRNA
double-stranded RNA
- GE
genetically engineered
- miRNA
microRNA
- siRNA
short-interfering RNA
Introduction
MicroRNAs (miRNA) are highly conserved, non-protein coding, RNA molecules of approximately 22 nucleotides that negatively regulate gene expression throughout the plant and animal kingdoms. They bind specific sequences in the 3′UTR of target mRNA transcripts and regulate post-transcriptional gene expression through the repression of translation and/or degradation of the targeted mRNA (reviewed in1). Interestingly, miRNAs can be specially processed2 for secretion to regulate gene expression in a non-cell-autonomous manner.1,3-12 These specialized miRNAs are remarkably stable and are resistant to degradation even in harsh environmental conditions,2,13,14 opening up the possibility for organismal and even inter-organismal level regulation by miRNA. In fact, recent evidence suggests that organisms connected in a wide array of ecological interactions use transfer of small RNA molecules to communicate.15-17 Such interactions appear possible within and across kingdoms and the resulting control of gene expression mediated by the RNA transfer can result in both beneficial and detrimental effects on recipient organisms.
Natural cross species transfer of diet-derived small RNA to ingesting organisms has been described in both vertebrates18 and invertebrates.19,20 The first description of such a phenomenon, in the report by C-Y Zhang and colleagues,18 showed that dietary plant miRNAs could enter the mammalian bloodstream and regulate cholesterol metabolism in the ingesting animal. This study stimulated much interest and further investigation21-24 since its initial publication. Since then, however, multiple groups have been unable to reproduce these findings. In fact, independent follow-up studies suggest that systemic uptake of orally ingested foreign miRNAs, as a class of molecules, is negligible25-27 and significantly below levels required to be biologically relevant when acting through canonical sequence-specific miRNA-mediated mechanisms. More recently, it has been suggested that dietary uptake could be enhanced via intestinal or renal injury, but only with artificially high levels of oral ingestion.28,29 In total, such work may suggest that horizontal miRNA transfer is possible, but severely limited under naturally occurring conditions in multiple organisms, including mice, humans, and honey bees. Nonetheless, controversy remains.30-45
The possibility of cross-kingdom regulation of gene expression via ingested miRNAs could be of particular interest in the mutualistic relationships of plants and their pollinators. The relationship involves an exchange of nutrients for the reproduction-enabling service of pollination, but is complicated by the antagonistic relationship of the two players, which is characterized by a fundamental conflict of interest.46 Demonstrating that plants can control gene expression in pollinating insects via small RNA transfer would reveal a new and exciting level of complexity in these plant-pollinator relationships. As one of the most highly studied pollinators, honey bees provide an excellent model for exploring the role of dietary transfer of small RNA in communication between plants and pollinators. In addition, RNAi-based technologies which take advantage of cross-kingdom small RNA (miRNA, siRNA, dsRNA) transfer, including genetically engineered (GE) plants and topical sprays, will be among the cutting edge developments with the potential to transform agriculture in the coming years.47,48 When used as pesticides, these technologies require rigorous research to assess their safety, especially with regards to effects on off-target organisms, such as honey bees. The efficacy of strategies to use GE plant and topical spray technology in combatting the diseases of beneficial insects also requires a solid understanding of uptake potential in target species, such as the honey bee. To date, no study has fully examined the role of dietary uptake of miRNA in the honey bee.
We previously demonstrated the absence of biologically relevant levels of plant-derived miRNAs in recipient honey bees in the course of normal behavior in managed colonies.25 However, these results were not fully conclusive in the absence of experiments examining diet-derived uptake under conditions in which we possessed the ability to exert direct control over the diet of individual honey bees. Here, in the context of a specifically controlled pollen diet, we demonstrate that ingested miRNAs are not efficiently taken up by the digestive tract or dispersed to distal tissues. In addition, expanded experiments examining midgut and abdominal levels of pollen-derived miRNA in honey bees of different life-stages taken directly from the colony support the conclusions of our feeding experiments. These results therefore reinforce the idea that pollen-derived miRNAs ingested as part of a typical diet are not effectively taken up by cells of the honey bee under normal conditions.
Results
Honey bee–collected pollen from multiple species contains high levels of plant miRNAs
We first wished to determine the levels of a highly conserved plant miRNA that is robustly expressed in fresh honey bee-collected pollen of different plant species. Based on available data in model plants49-51 and our previous data, we selected the highly conserved plant miRNA, MIR156a, which we have found to be highly expressed in pollen-derived bee bread and honey.25 Using qPCR with a Taqman probe specific for MIR156a, we measured the level of this miRNA in thirty-two different samples of honey-bee pollen collected over 2 seasons. Copy number was calculated by using a standard curve for a synthetic MIR156a mimic (a representative standard curve is shown in Fig. S1A) and represented as copy number per 10 pg of total RNA tested (10 pg of RNA representing a conservative estimate of the total RNA content of a single eukaryotic cell). We found that MIR156a was expressed at high levels in all samples of honey bee-collected pollen, albeit at variable levels (Fig. 1A, B), with an average of 47.5 copies per 10 pg of RNA and 1.63 × 106 copies per mg of fresh weight. Using ITS2 sequences, we identified the genus (and highest ranked species) of select pollen samples (Table 1) and found that pollen samples from different genera contain high, but variable levels of MIR156a.
Figure 1.
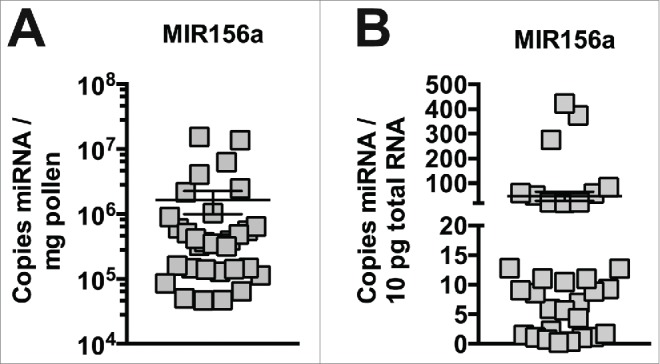
High levels of miRNA in pollen from different species. Plant MIR156a levels in fresh pollen collected by honey bee foragers (n = 32). Copy number is expressed per mg (A) and per 10pg total RNA (B).
Table 1.
Species Identification and MIR156a Levels of Honey Bee Collected Pollen. List of miRNA MIR156a levels in fresh pollen collected by honey bee foragers and the genus (and highest ranked species) of the plant of origin. Copy number is expressed per mg and per 10 pg total RNA.
| Genus, highest ranked species | copy number | copy number |
|---|---|---|
| (Date Collected) |
×105/ mg |
10 pg |
| Rosa, rubus (4/15) | 1.13 | 9.3 |
| Sophora, japonica (4/15) | 1.29 | 51.3 |
| Ulmus, minor (4/15) | 5.5 | 23.4 |
| Oxygraphis, glacialis (4/15) | 3.89 | 21.6 |
| Salix, polaris (4/15) | 6.01 | 6.9 |
| Ulmus, minor (4/15) | 153 | 275.2 |
| Pinus, contorta (4/15) | 8.99 | 8.86 |
| Celastrus, orbiculatus (5/15) | 3.68 | 8.6 |
| Rubus, rosa (5/15) | 136 | 83.2 |
| Maclura, pomifera (5/15) | 40.5 | 374 |
| Rubus, rosa (5/15) | 5.1 | 23.2 |
| Ligustrum, ovalifolium (6/15) | 6.45 | 12.7 |
| Hydrangea, xanthoneura (6/15) | 4.17 | 11.0 |
| Ligustrum, ovalifolium (6/15) | 62.1 | 422.8 |
| Trifolium, repens (6/15) | 10.37 | 60.6 |
| Securigera, varia (7/15) | 1.48 | 1.2 |
| Trifolium, repens (7/15) | 0.5 | 1.1 |
| Campsis, radicans (7/14) | 21.6 | 5.93 |
| Plantago, rugelii (7/14) | 3.25 | 12.8 |
| Trifolium, repens (7/14) | 24.7 | 59 |
Conserved plant miRNA, MIR156a, levels in fresh pollen collected by honey bee foragers. Copy number is expressed per mg and per 10pg total RNA.
Newly eclosed, nurse, and forager honey bees have insignificant levels of plant miRNAs in abdominal tissue
In our previous study, we found that both nurse and forager adult honey bees had only minimal levels of MIR156a in abdominal tissue. However, in that study we did not examine miRNAs in the midgut and therefore could not be sure of recent pollen consumption. Many factors are known to impact honey bee pollen consumption. For example, honey bees of different life stages are known to consume variable amounts of pollen dependent on stage (Fig. 2A). The variable levels MIR156a in pollen of different species observed above, as well as the likelihood of rapid clearance of pollen-derived miRNAs even if uptake occurs, further underscore the critical significance of assuring the amount and timing of ingested miRNA levels.
Figure 2.
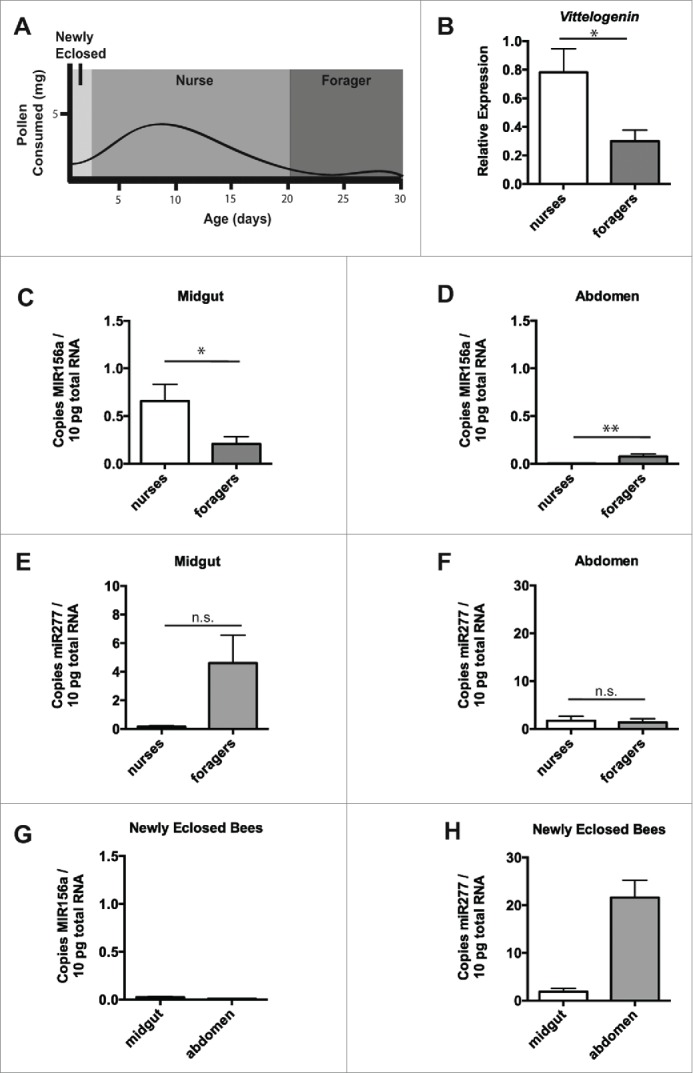
Nurse and forager bees exhibit minimal levels of plant miRNA in abdominal tissue. Schematic of pollen consumption during honey bee life stages (based on55,57) (A). Transcript levels of Vitellogenin relative to β-actin in midgut and abdominal tissue from nurse and forager bees taken directly from the hive (B). Plant MIR156a levels in midgut (C) or abdominal tissue (D) from nurse (n =8 ) or forager (n = 8) honey bees taken directly from the hive. Levels of endogenous mir277 in midgut (E) or abdominal tissue (F) from nurse and forager bees above. Plant MIR156a levels in midgut or abdominal tissue from newly eclosed honey bees from frames taken directly from the hive (G). Levels of endogenous mir277 in midgut or abdominal tissue from newly eclosed bees above (H). Copy number is expressed per 10pg total RNA. Data is expressed as Mean ± SEM, * signifies p < 0.05; ** signifies p < 0.01, n.s. signifies not significant, p > 0.05.
To extend our previous research, we first collected honey bee nurses and forager as before52 and examined the levels of MIR156a in both midgut and abdominal tissues. To verify our selection of nurses and foragers, we examined the levels of Vitellogenin (Vg) mRNA in the abdominal tissue of the collected bees and found decreased Vg expression in the abdomens of foragers (Fig. 2B), confirming the membership of bees in the nurse and forager castes. Importantly, near equivalence was observed in midgut and abdominal expression of the housekeeping gene β-actin between these two groups (Fig. S1B). We also observed modest levels of MIR156a in the midgut of both nurse and forager bees (Fig. 2C). Despite the levels of MIR156a in the digestive tract, we observed very low levels of MIR156a in abdominal tissue (less than 0.1 copy per 10 pg of total RNA) (Fig. 2D). For reference, miR277, a miRNA endogenous to bees53 and active in regulation of amino acid metabolism,54 was expressed at levels ∼50-fold more than the ingested MIR156a but at near equivalent levels in the midgut tissue (Fig. 2E) and abdominal tissue (Fig. 2F) when comparing nurse and forager bees.
Newly eclosed bees are known to have different behavioral and physiological characteristics than nurse and forager bees.55 To determine whether this youngest group of bees differed in miRNA uptake, we collected newly eclosed honey bees from frames incubated at colony temperature overnight and examined the levels of MIR156a in both the midgut and abdominal tissues. We observed very low levels of MIR156a in the midgut and in the abdominal tissue (less than 0.3 copy per 10 pg of total RNA) of these bees (Fig. 2G). Endogenous miR277 was expressed at high levels in both midgut and abdomen (Fig. 2H)). β-actin (Fig. S1C) was expressed at higher levels compared to that observed for older bees.
Negligible transfer of diet-derived miRNA to internal tissue after pollen ingestion
Previous findings demonstrating the absence of biologically relevant levels of plant-derived miRNAs in recipient honey bees remained indecisive in the absence of controlled feeding experiments. To further investigate dietary uptake of miRNA in honey bees, we examined uptake in honey bees fed a diet containing sugar syrup with polyfloral pollen or sugar syrup alone. To provide a more accurate baseline, we attempted to remove any residual pollen from the system of recipient bees. A previous study found that feeding a pollen-free diet for 24 hours results in digestion of previously consumed pollen.56 In agreement, we have found that bees fed a pollen-free diet for 24 hours have negligible numbers of intact pollen grains in their midgut contents (data not shown) and very low levels of pollen-derived miRNA (Fig. S2A). However, we chose to feed bees a pollen-free diet for 48 hours prior to our investigations of diet-derived miRNA transfer to reduce the pre-experiment pollen levels in the digestive tract even further. Subsequently, a pollen dose of 50 mg/ml was given to ensure that individual bees would ingest ∼5 mg of total pollen
We collected bees returning to the hive without pollen loads, fed them a diet of sugar syrup alone for 48 hours, starved them for 1 hour to encourage feeding, and then fed them 30% sucrose with or without polyfloral pollen provided at a concentration of 50 mg/ml for 24 hours. Levels of MIR156a in the polyfloral pollen mixture were comparable to that observed in pollen from single species above (data not shown). Only minimal levels of MIR156a (less than 0.01 copy per 10 pg of total RNA) were detected in the midgut tissue of bees fed sugar water alone for 72 hours. We observed a 32-fold increase in MIR156a levels in the midgut tissue of bees fed sugar water containing pollen compared to those receiving sugar water alone (Fig. 3A), to ∼0.5 copies per 10 pg of total RNA (p < 0.05). This agreed very closely with the amount of MIR156a found in the midguts of nurse bees taken directly from the hive (Fig. 2C), which are known to have a diet high in pollen. Despite the high levels of MIR156a in pollen and robust increase of pollen-derived MIR156a in the midgut tissue, only very low levels of MIR156a in abdominal tissue were observed (less than 0.1 copy per 10 pg of total RNA) and no significant difference in levels between abdominal tissue from bees fed sugar water with pollen compared to those receiving sugar water alone (Fig. 3B). We observed equivalent levels of miR277 in the midgut tissue (Fig. 3C) and abdominal tissue (Fig. 3D) of bees fed sugar water with pollen compared to those bees receiving sugar water alone. We also found no difference in the levels of β-Actin transcript as assessed by Ct values (Fig. S2B) between honey bees in the different treatment groups.
Figure 3.
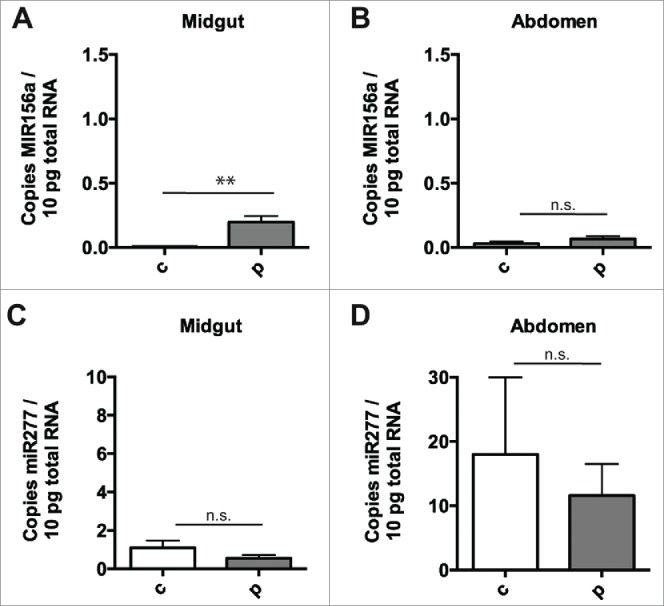
Negligible transfer of diet-derived miRNA to internal tissue after pollen ingestion. Plant miRNA MIR156a levels, expressed per 10pg total RNA, in honey bee midgut (A) and abdomen (B) after 24 hours of ingestion of sugar solution alone (n = 7) or sugar solution containing polyfloral pollen (n = 7) (after 48 hours of pollen-free diet). Endogenous honey bee miRNA mir277 levels in the honey bees above, expressed per 10pg total RNA, in honey bee midgut (C) and abdomen (D) after 24 hours of ingestion of sugar solution alone or sugar solution containing polyfloral pollen (after 48 hours of pollen-free diet). Copy number is expressed per 10pg total RNA. Data is expressed as Mean ± SEM, * signifies p < 0.05; ** signifies p < 0.01, n.s. signifies not significant, p > 0.05.
The MIR156 observed in the midgut was likely from ingested pollen and not actual delivery to cells of the digestive tract. However, we wished to rule out the possibility that midgut epithelial cells were in fact taking up the MIR156a. To do so, we performed a similar experiment as above and tested the effect of removing the midgut food bolus, which can be removed in 1-2 discrete sacs coated by the peritrophic matrix. Plant MIR156a levels in whole honey bee midguts were elevated in honey bees ingesting pollen as above (Fig. 4A). However, we observed no increase in the MIR156a levels in midguts with food boli removed (Fig. 4A)Boli, which contained visible pollen (Fig. 4B), were confirmed to carry the expected substantial levels of MIR156a (Fig. 4C), similar to a midgut from a pollen-fed bee without bolus removed.
Figure 4.
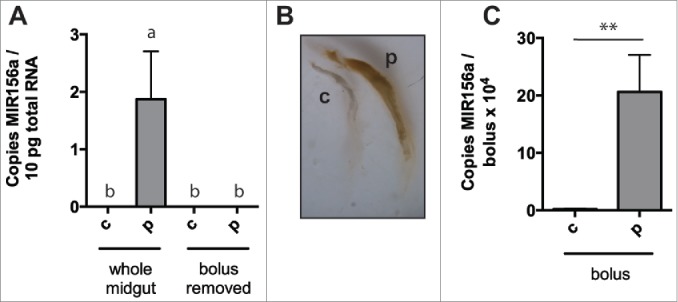
Negligible uptake of diet-derived miRNA by digestive tract epithelia after pollen ingestion. Plant miRNA MIR156a levels, expressed per 10pg total RNA, in whole honey bee midguts or midguts with food bolus removed after 24 hours of ingestion of sugar solution alone (n = 3) or sugar solution containing polyfloral pollen (n = 3) (after 48 hours of pollen-free diet) (A). Brightfield microscopic image of food boluses from midguts above (B). Plant miRNA MIR156a levels, expressed per bolus, in food boli removed from midguts above (n = 3) (C). Data is expressed as Mean ± SEM. In (A) a ≠ b, p < 0.05, the differences between b are not significant and in (B) * signifies p < 0.05; ** signifies p < 0.01.
Discussion
The controlled feeding trials here demonstrated the absence of substantial uptake and systemic delivery of MIR156a despite documented oral ingestion of a typical diet in adult honey bees. We found midgut MIR156a at similar levels to that observed for the endogenous miR277. However, as evident from experiments where the food bolus was removed, such levels likely originate from ingested pollen itself and not actual delivery. We also did not find evidence of significant transfer of dietary miRNA from the digestive tract to distal tissue. Consistent with our previous pilot data25, the simplest explanation is that transfer of miRNA ingested in the diet to proximal or remote tissues is not efficient in honey bees. Yet, while our results suggest a heavily restricted role for systemic changes in gene expression as a result of the dietary trans-kingdom transfer of naturally occurring small RNA molecules, a number of possible influencing factors remain to be clarified.
First, transfer of consumed plant miRNAs may be limited by insufficient copy number of miRNA in pollen. Bees are thought to consume a maximum of ∼4 mg of pollen per day.57 We observed highly variable levels of MIR156a in pollen from different species and even differences in the levels of MIR156a in pollen from the same species. This is not surprising as other pollen indices, such as protein content and viability, are known to vary extensively between members of the same species, based on genotypic and environmental influences on plant health.58,59 Using the highest level observed for pollen in this study, we can calculate that an adult honey bee might consume approximately 6 × 107 copies of MIR156a per day, resulting in a maximum theoretical delivery of 6 copies per cell if 100% transfer of the diet-derived miRNA were achieved (using 1 × 107 cells per bee as previously25). Thus, even under these optimal theoretical conditions, it is difficult to envision dietary miRNA reaching distal tissues in biologically relevant levels unless aided by specialized mechanisms for concentrating or transferring miRNA in specific cell types. Here, we observed transfer to occur far below the theoretical optimum, which is in line with other studies examining insects.21 Such a limiting factor may be overcome by a number of scenarios. Evidence in multiple systems, including the honey bee, suggests that selective transport and amplification of small RNA exists.60-62 Thus, specific miRNA species not examined here might be taken up more efficiently into tissues. In addition, specialized populations within a given tissue might possess mechanisms for efficient uptake that would result in high levels of miRNA in these cells that remains undetectable within the tissue as a whole in our assays. Finally, specific tissues not examined here might be capable of miRNA absorption. Any of these cases could theoretically result in levels of small RNAs derived from the diet that are capable of altering gene expression in various tissues.
Second, the uptake or bioavailability of dietary miRNA may change in a manner dependent on physiological characteristics of individual bees, such as the developmental stage or age. A recent study has suggested that bee-derived miRNAs may be transferred to larval honey bees via inoculation of royal jelly with miRNAs by adult nurse bees, leading to specific effects on gene expression during larval development.63,64 However, actual uptake and transfer of dietary miRNA has not been demonstrated in any previous study. Oral dsRNA delivery can vary depending on insect life-stage in other insects65 and miRNA uptake may similarly change with age. Thus, it is possible that specific circumstances in the larvae could enhance accumulation of these miRNAs in recipient tissues. Honey bees exhibit an age-based societal structure, known as age polyethism, in which a major life transition is found in the switch from nurse bees, which perform in hive task, to forager bees, which perform tasks outside the hive.55 One hallmark of this transition is a reduction in protein consumption, manifested as a reduction in both the amount of pollen ingested57 and the ability to digest pollen due to reduced digestive enzyme production.66 Thus, it is possible that honey bees larvae or nurse bees would have higher uptake of dietary miRNA due to more efficient digestion of pollen than the bees used in feeding experiments here, which are likely foragers due to the location of their collection on the landing board. However, our results showed no difference in the levels of MIR156a in abdominal tissue of nurse, forager, or newly emerged bees, suggesting that age does not play a role in uptake in adult worker bees. Uptake could also be dissimilar in members of the two reproductive castes of honey bees, the queen and the male drones, and this possibility may be important to examine. In addition, other physiologic conditions may play a role, including potential alteration of the permeability or selective transport mechanisms of the gut epithelia by various stressors and disease states.
Thirdly, the most stringent impediment to uptake is likely the limitation imposed by the highly selective barrier of healthy gut epithelial tissue, which in most organisms studied to date does not possess mechanisms for robust uptake of dietary miRNA.65 Invertebrates as a group do appear to be more competent to absorb dietary miRNA and dsRNA into tissues, although uptake is highly variable in a manner that is dependent on the type of small RNA molecule used and the invertebrate species examined. For example, the worm C. elegans is remarkably proficient in uptake of dietary dsRNA and spread to distal tissues,67 but success with other insects has been variable.61,68 Ingestion of naked dsRNA has been successfully used to modify gene expression in multiple insect taxa, again with species to species variation in successful uptake and regulation of endogenous gene expression. In agreement, oral administration of dsRNA appears to be effective at regulation of gene expression in honey bees.69-76 In addition, high dose injection of dsRNAs into the hemocoel has been used for delivery and inhibition of target genes in multiple tissues in this species77-86 independent of the site of injection, demonstrating that these molecules can be efficiently taken up by disparate tissues once in the open circulatory system. Our results, in conjunction with the previous studies using dsRNA in other insect species, suggest that uptake of longer dsRNAs may be more efficient than shorter miRNAs. However, uptake of short miRNA mimics have been shown to be taken up in some insects,87 again underscoring the diversity of uptake potential among species. Plants possess a number of small RNA types, including TasiRNA, which are generated from longer dsRNA precursors.88 Examining the dietary uptake of these naturally occurring dsRNA may therefore be warranted. In addition to the size of the small RNA species involved, packaging of plant small RNAs in exosomes or other such structure may cause differences in delivery efficiency.
In summary, our results provide more definitive evidence that, despite ingesting high levels of pollen-derived miRNA, honey bees display systemic levels of these miRNAs at concentrations far below those which would make canonical functions of miRNAs likely.89 These results suggest that the honey bee can provide an additional relevant model to examine dietary uptake of naturally occurring and artificially provided miRNA, siRNA, and dsRNA in field populations of invertebrates. Such a model can provide a first assessment of the potential effects of GE plants and topical sprays using small RNA on beneficial insects, including critically important and understudied native pollinators.90 However, it is important to note that based on our current knowledge, uptake of diet-derived miRNA, siRNA, and dsRNA in is fact quite varied between different invertebrate species and may ultimately require assessment on a species by species basis. While the balance of data therefore supports the idea that small RNA-based technologies, such as GE plants, are safe to off-target organisms under normal conditions, important concerns still remain as outlined above and as follows. One such concern focuses on the effect that GE-tech mediated overexpression of a novel miRNA will have effects on the amount of miRNA traversing the gut epithelia. A second key point centers on the ability of a variety of stressors and disease states to alter or compromise the permeability or selective transport mechanisms of the gut epithelia, thus allowing an increased level uptake of environmental molecules, including exogenous miRNA, leading to unknown off-target effects in these organisms. Further research will be required to determine whether these factors could impact uptake of natural and GE-tech diet-derived miRNA by honey bees.
Materials and methods
Honey bee and pollen collection
Honey bees were collected from the landing board of outbred colonies in New York, New York consisting of a typical mix of Apis mellifera subspecies found in North America, at different times during the months of April-October. Only visibly healthy bees were collected and all source colonies were visually inspected for symptoms of common bacterial, fungal, and viral diseases of honey bees. When bees of specific life stage were required, nurses and foragers were collected from the same colonies at identical times using a mouth aspirator as before.52 Newly eclosed bees were obtained by placing frames of emerging young bees from healthy colonies into an incubator (34°C, 60% relative humidity) overnight. For feeding experiments, bees returning to the hive without pollen loads were collected. For pollen collection, pollen was collected from the corbiculae of returning foragers from the colonies above. Quantification of pollen-derived miRNA in nurse, forager, and newly eclosed bees was performed two independent times with similar results.
Feeding treatments
For all caged experiments, honey bees were selected as above and kept in 177.4 mL (6 oz.) Square-bottomed Drosophila Stock Bottles (VWR) plugged with modified foam tube plugs (Jaece Industries). Bees were maintained in incubators at 35°C in the presence of PseudoQueen (Contech) as a source of Queen Mandibular Phermone (QMP) and used as per manufacturer's instructions. Bees were fed via a modified 1.5 ml screw-cap tube. Bees were fed a diet of sugar syrup alone for 48 hours, starved for 1 hour to encourage feeding, and then fed 30% sucrose with or without polyfloral pollen provided at a concentration of 50 mg/ml. Feeding experiments were performed four independent times with similar results.
RNA extraction
After cold anesthesia, honey bee gut tissue was removed from abdomens for analysis, and the remaining abdominal tissue was independently used for gene expression analysis. In some experiments, the food bolus was removed from the midgut by removing the peritrophic matrix, which slides out as 1 or 2 discrete sacs, and its contents. All dissected material was placed into RNAlater (Invitrogen) for storage prior to analysis. RNA was then extracted as previously described25 via the miRNeasy kit (Qiagen) using phenol-chloroform extraction, and column-based extraction. Total RNA concentration was calculated by Nanodrop spectrophotometry (ThermoScientific). The average number of cells in a honey bee was calculated as before. Briefly, the number of cells in a honey bee = mass of an average worker bee (90 mg)/mass of an individual cell. Mass of an individual cell was calculated using the equation: mass = cell volume x cell density where an average cell volume is estimated at 10 μm3 with a density of 1. For pollen, ∼10 mg was added to Qiazol and then homogenized using mechanical disruption. RNA was then extracted as above
RNA was eluted in 42 µl H2O. 5 µl (of the 42 µl volume), corresponding to 100-200 ng of RNA for pollen and 1-2 μg for tissue used as a template for miRNA reverse transcription (Taqman MicroRNA Reverse Transcription Kit, Applied Biosystems) in a total reaction volume of 15 μl. We used this relatively large amount of total RNA for reverse transcription to optimize the ability to detect low levels of miRNAs. Following reverse transcription, 1 µl (of the 15 µl volume) of the RT product was then used as template in qPCR with a total reaction volume of 20 µl.
Quantification of microRNA and determination of copy number
To quantify levels of select miRNAs, standard TaqMan reverse transcription-quantitative (“real time”) polymerase chain reaction (RT-QPCR) was utilized, as we previously described.25 TaqMan primer/probes for quantification were used as follows: ath-MIR156a (AB 4427975 (000333)) and ame-miR277 (AB 4427975 (000298))(Applied Biosystems). Mature miRNA copy numbers were then calculated from raw Ct values through generation of individual standard curves for each miRNA Taqman primer/probe set after making serial dilutions of a known quantity of synthetic oligonucleotide mimic for each mature miRNA ath-MIR156a (AB 4464066 (000333)) and ame-miR277 (AB 4464066 (000298)) (Applied Biosystems). While previously we have reported that we can accurately detect 1-3 copies at a Ct of 37, here we structured the study such that we did not have to rely upon such extreme sensitivity for our calculations, thus improving our confidence in copy number quantification. Specifically, we made a standard curve (as shown in Suppl Fig. 1A) where our last point of measurement was 70 copies at a Ct of 33.2. Quantification of mRNA via qPCR was performed as before.52
Species identification of honey bee-collected pollen
For source plant species identification of pollen collected by honey bees, the rRNA internal transcribed spacer 2 (ITS2) sequence was determined. To obtain ITS2 sequence from pollen, cDNA was synthesized from RNA extracted above, using the iScript cDNA Synthesis Kit (Biorad), and 5 μl of cDNA was then used as a template for PCR using the GoTaq polymerase and buffer (Promega). The universal plant ITS2 primer sequences used (ITS2F: 5′- ATGCGATACTTGGTGTGAAT-3′ and ITS3R: 5′- GACGCTTCTCCAGACTACAAT-3′) and PCR conditions have been described.91 After visualizing a fraction of the PCR products on a gel, the remainder was purified using a PCR Purification Kit (Qiagen) and sequenced (Genewiz). Sequences were annotated and the genus (and highest ranked species) of the plant of origin was identified using the ITS2 Database.92
Statistical analysis
Data is expressed as Mean ± standard error of the mean (SEM). N = number of independent animals. For all data, mean values were calculated for each population and normality was assessed using Shapiro–Wilk tests on log10 transformed values. When two groups were compared, significance was assessed using unpaired t-tests with Welch's correction when values fit normal distributions or Mann-Whitney U nonparametric tests when they did not fit normal distributions. When more than two groups were compared, ANOVA was used to assess significance with Tukey's multiple comparison test.
Supplementary Material
Disclosure of potential conflicts of interest
No potential conflicts of interest were disclosed.
Acknowledgments
The authors acknowledge the technical assistance of Nina Deoras and Jessica Sam in the completion of select experiments. Also, the authors thank Jennifer Mansfield for helpful comments and critical review of the manuscript.
References
- 1.Bartel DP. MicroRNAs: genomics, biogenesis, mechanism, and function. Cell 2004; 116:281-97; PMID:14744438; http://dx.doi.org/ 10.1016/S0092-8674(04)00045-5 [DOI] [PubMed] [Google Scholar]
- 2.Ha M, Kim VN. Regulation of microRNA biogenesis. Nat Rev Mol Cell Biol 2014; 15:509-24; PMID:25027649; http://dx.doi.org/ 10.1038/nrm3838 [DOI] [PubMed] [Google Scholar]
- 3.Vickers KC, Palmisano BT, Shoucri BM, Shamburek RD, Remaley AT. MicroRNAs are transported in plasma and delivered to recipient cells by high-density lipoproteins. Nat Cell Biol 2011; 13:423-33; PMID:21423178; http://dx.doi.org/ 10.1038/ncb2210 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 4.Gupta SK, Bang C, Thum T. Circulating microRNAs as biomarkers and potential paracrine mediators of cardiovascular disease. Circ Cardiovasc Genet 2010; 3:484-8; PMID:20959591; http://dx.doi.org/ 10.1161/CIRCGENETICS.110.958363 [DOI] [PubMed] [Google Scholar]
- 5.Hale A, Lee C, Annis S, Min P-K, Pande R, Creager MA, Julian CG, Moore LG, Mitsialis SA, Hwang SJ, et al.. Biochim Biophys Acta. BBA - Molecular Cell Research 2014; 1843:2528-42; PMID:24983771; http://dx.doi.org/ 10.1016/j.bbamcr.2014.06.012 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 6.Valadi H, Ekström K, Bossios A, Sjöstrand M, Lee JJ, Lötvall JO. Exosome-mediated transfer of mRNAs and microRNAs is a novel mechanism of genetic exchange between cells. Nat Cell Biol 2007; 9:654-9; PMID:17486113; http://dx.doi.org/ 10.1038/ncb1596 [DOI] [PubMed] [Google Scholar]
- 7.Zhang Y, Liu D, Chen X, Li J, Li L, Bian Z, Sun F, Lu J, Yin Y, Cai X, et al.. Secreted monocytic miR-150 enhances targeted endothelial cell migration. Mol Cell 2010; 39:133-44; PMID:20603081; http://dx.doi.org/ 10.1016/j.molcel.2010.06.010 [DOI] [PubMed] [Google Scholar]
- 8.Kosaka N, Iguchi H, Yoshioka Y, Takeshita F, Matsuki Y, Ochiya T. Secretory mechanisms and intercellular transfer of microRNAs in living cells. Journal of Biological Chemistry 2010; 285:17442; PMID:20353945; http://dx.doi.org/ 10.1074/jbc.M110.107821 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 9.Pegtel DM, Cosmopoulos K, Thorley-Lawson DA, van Eijndhoven MAJ, Hopmans ES, Lindenberg JL, de Gruijl TD, Wurdinger T, Middeldorp JM. Functional delivery of viral miRNAs via exosomes. Proc Natl Acad Sci USA 2010; 107:6328-33; PMID:20304794; http://dx.doi.org/ 10.1073/pnas.0914843107 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 10.Mittelbrunn MIA, zquez CGER-VA, Villarroya-Beltri C, lez SGA, nchez-Cabo FATSA, lez MANGA, Bernad A, nchez-Madrid FSA. unidirectional transfer of microRnA-loadedexosomes from T cells to antigen-presenting cells. Nat Commun 2011; 2:282-10; PMID:21505438; http://dx.doi.org/ 10.1038/ncomms1285 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 11.Hergenreider E, Heydt S, Tréguer K, Boettger T, Horrevoets AJG, Zeiher AM, Scheffer MP, Frangakis AS, Yin X, Mayr M, et al.. Atheroprotective communication between endothelial cells and smooth muscle cells through miRNAs. Nat Cell Biol 2012; 14:249-56; PMID:22327366; http://dx.doi.org/ 10.1038/ncb2441 [DOI] [PubMed] [Google Scholar]
- 12.Zhuang G, Wu X, Jiang Z, Kasman I, Yao J, Guan Y, Oeh J, Modrusan Z, Bais C, Sampath D, et al.. Tumour-secreted miR-9 promotes endothelial cell migration and angiogenesis by activating the JAK-STAT pathway. EMBO J 2012; 31:3513-23; PMID:22773185; http://dx.doi.org/ 10.1038/emboj.2012.183 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 13.Jung M, Schaefer A, Steiner I, Kempkensteffen C, Stephan C, Erbersdobler A, Jung K. Robust MicroRNA stability in degraded RNA preparations from human tissue and cell samples. Clin Chem 2010; 56:998-1006; PMID:20378769; http://dx.doi.org/ 10.1373/clinchem.2009.141580 [DOI] [PubMed] [Google Scholar]
- 14.Howard KM, Jati Kusuma R, Baier SR, Friemel T, Markham L, Vanamala J, Zempleni J. Loss of miRNAs during processing and storage of cow's ( Bos taurus) milk. J Agric Food Chem 2015; 63:588-92; PMID:25565082; http://dx.doi.org/ 10.1021/jf505526w [DOI] [PMC free article] [PubMed] [Google Scholar]
- 15.Sarkies P, Miska EA. Molecular biology. Is there social RNA? Science 2013; 341:467-8; PMID:23908213; http://dx.doi.org/ 10.1126/science.1243175 [DOI] [PubMed] [Google Scholar]
- 16.Knip M, Constantin ME, Thordal-Christensen H. Trans-kingdom cross-talk: Small RNAs on the move. PLoS Genet 2014; 10:e1004602; PMID:25188222; http://dx.doi.org/ 10.1371/journal.pgen.1004602 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 17.Weiberg A, Bellinger M, Jin H. ScienceDirectConversations between kingdoms: small RNAs. Curr Opin Biotechnol 2015; 32:207-15; PMID:25622136; http://dx.doi.org/ 10.1016/j.copbio.2014.12.025 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 18.Zhang L, Hou D, Chen X, Li D, Zhu L, Zhang Y, Li J, Bian Z, Liang X, Cai X, et al.. Exogenous plant MIR168a specifically targets mammalian LDLRAP1: evidence of cross-kingdom regulation by microRNA. Cell Res 2011; 22:107-26; PMID:21931358; http://dx.doi.org/ 10.1038/cr.2011.158 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 19.Liu H, Wang X, Wang H-D, Wu J, Ren J, Meng L, Wu Q, Dong H, Wu J, Kao T-Y, et al.. Escherichia coli noncoding RNAs can affect gene expression and physiology of Caenorhabditis elegans. Nat Commun 2012; 3:1073-11; PMID:23011127; http://dx.doi.org/ 10.1038/ncomms2071 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 20.Ivashuta S, Zhang Y, Wiggins BE, Ramaseshadri P, Segers GC, Johnson S, Meyer SE, Kerstetter RA, McNulty BC, Bolognesi R, et al.. Environmental RNAi in herbivorous insects. Rna 2015; 21:840-50; PMID:25802407; http://dx.doi.org/ 10.1261/rna.048116.114 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 21.Zhang Y, Wiggins BE, Lawrence C, Petrick J, Ivashuta S, Heck G. Analysis of plant-derived miRNAs in animal small RNA datasets. BMC Genomics 2012; 13:1-1; PMID:22214261; http://dx.doi.org/ 10.1186/1471-2164-13-1 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 22.Petrick JS, Brower-Toland B, Jackson AL, Kier LD. Safety assessment of food and feed from biotechnology-derived crops employing RNA-mediated gene regulation to achieve desired traits: a scientific review. Regul Toxicol Pharmacol 2013; 66:167-76; PMID:23557984; http://dx.doi.org/ 10.1016/j.yrtph.2013.03.008 [DOI] [PubMed] [Google Scholar]
- 23.Hirschi KD. New foods for thought. Trends Plant Sci 2012; 17:123-5; PMID:22265093; http://dx.doi.org/ 10.1016/j.tplants.2012.01.004 [DOI] [PubMed] [Google Scholar]
- 24.Witwer KW. XenomiRs and miRNA homeostasis in health and disease: evidence that diet and dietary miRNAs directly and indirectly influence circulating miRNA profiles. RNA Biol 2012; 9:1147-54; PMID:22951590; http://dx.doi.org/ 10.4161/rna.21619 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 25.Snow JW, Hale AE, Isaacs SK, Baggish AL, Chan SY. Ineffective delivery of diet-derived microRNAs to recipient animal organisms. RNA Biol 2013; 10:1107-16; PMID:23669076; http://dx.doi.org/ 10.4161/rna.24909 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 26.Witwer KW, McAlexander MA, Queen SE, Adams RJ. Real-time quantitative PCR and droplet digital PCR for plant miRNAs in mammalian blood provide little evidence for general uptake of dietary miRNAs: Limited evidence for general uptake of dietary plant xenomiRs. RNA Biol 2013; 10:1080-6; PMID:23770773; http://dx.doi.org/ 10.4161/rna.25246 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 27.Dickinson B, Zhang Y, Petrick JS, Heck G, Ivashuta S, Marshall WS. correspondence. Nat Biotechnol 2013; 31:965-7; PMID:24213763; http://dx.doi.org/ 10.1038/nbt.2737 [DOI] [PubMed] [Google Scholar]
- 28.Yang J, Farmer LM, Agyekum AAA, Hirschi KD. Detection of dietary plant-based small RNAs in animals. Cell Res 2015; 25:517-20; PMID:25721324; http://dx.doi.org/ 10.1038/cr.2015.26 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 29.Mlotshwa S, Pruss GJ, MacArthur JL, Endres MW, Davis C, Hofseth LJ, Peña MM, Vance V. A novel chemopreventive strategy based on therapeutic microRNAs produced in plants. Cell Res 2015; 25:521-4; PMID:25721325; http://dx.doi.org/ 10.1038/cr.2015.25 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 30.Baier SR, Nguyen C, Xie F, Wood JR, Zempleni J. MicroRNAs are absorbed in biologically meaningful amounts from nutritionally relevant doses of cow milk and affect gene expression in peripheral blood mononuclear cells, HEK-293 kidney cell cultures, and mouse livers. J Nutr 2014; 144:1495-500; PMID:25122645; http://dx.doi.org/ 10.3945/jn.114.196436 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 31.Witwer KW. Diet-responsive mammalian miRNAs are likely endogenous. J Nutr 2014; 144:1880-1; PMID:25332488; http://dx.doi.org/ 10.3945/jn.114.202523 [DOI] [PubMed] [Google Scholar]
- 32.Baier SR, Xie F, Zempleni J. Reply to witwer. J Nutr 2014; 144:1882; PMID:25332489 [DOI] [PubMed] [Google Scholar]
- 33.Chen X, Zen K, Zhang C-Y. Reply to Lack of detectable oral bioavailability of plant microRNAs after feeding in mice. Nat Biotechnol 2013; 31:967-9; PMID:24213764; http://dx.doi.org/ 10.1038/nbt.2741 [DOI] [PubMed] [Google Scholar]
- 34.Cottrill KA, Chan SY. Diet-derived microRNAs: Separating the dream from reality. microRNA Diagn Therap 2014; 1:46-57; http://dx.doi.org/ 10.2478/micrnat-2014-0001 [DOI] [Google Scholar]
- 35.Zhou Z, Li X, Liu J, Dong L, Chen Q, Liu J, Kong H, Zhang Q, Qi X, Hou D, et al.. Honeysuckle-encoded atypical microRNA2911 directly targets influenza A viruses. Cell Res 2015; 25:39-49; PMID:25287280; http://dx.doi.org/ 10.1038/cr.2014.130 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 36.Liang G, Zhu Y, Sun B, Shao Y, Jing A, Wang J, Xiao Z. Assessing the survival of exogenous plant microRNA in mice. Food Sci Nutr 2014; 2:380-8; PMID:25473495; http://dx.doi.org/ 10.1002/fsn3.113 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 37.Beatty M, Guduric-Fuchs J, Brown E, Bridgett S, Chakravarthy U, Hogg RE, Simpson DA. Small RNAs from plants, bacteria and fungi within the order Hypocreales are ubiquitous in human plasma. BMC Genomics 2014; 15:933; PMID:25344700; http://dx.doi.org/ 10.1186/1471-2164-15-933 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 38.Witwer KW, Hirschi KD. Transfer and functional consequences of dietary microRNAs in vertebrates: concepts in search of corroboration: negative results challenge the hypothesis that dietary xenomiRs cross the gut and regulate genes in ingesting vertebrates, but important questions persist. Bioessays 2014; 36:394-406; PMID:24436255; http://dx.doi.org/ 10.1002/bies.201300150 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 39.Petrick JS, Moore WM, Heydens WF, Koch MS, Sherman JH, Lemke SL. Regulatory toxicology and pharmacology. Regul Toxicol Pharmacol 2015; 71:8-23; PMID:25445299; http://dx.doi.org/ 10.1016/j.yrtph.2014.10.016 [DOI] [PubMed] [Google Scholar]
- 40.Jiang M, Sang X, Hong Z. Beyond nutrients: food-derived microRNAs provide cross-kingdom regulation. Bioessays 2012; 34:280-4; PMID:22354805; http://dx.doi.org/ 10.1002/bies.201100181 [DOI] [PubMed] [Google Scholar]
- 41.Nolte-'t Hoen ENM, van Rooij E, Bushell M, Zhang CY, Dashwood RH, James WPT, Harris C, Baltimore D. The role of microRNA in nutritional control. J Int Med 2015; 278:99-109; PMID:25832550; http://dx.doi.org/ 10.1111/joim.12372 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 42.Tosar JP, Rovira C, Naya H, Cayota A. Mining of public sequencing databases supports a non-dietary origin for putative foreign miRNAs: underestimated effects of contamination in NGS. RNA 2014; 20:754-7; PMID:24729469; http://dx.doi.org/ 10.1261/rna.044263.114 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 43.Hirschi KD, Pruss GJ, Vance V. Dietary delivery: a new avenue for microRNA therapeutics? Trends Biotechnol 2015; 33:431-2; PMID:26113189; http://dx.doi.org/ 10.1016/j.tibtech.2015.06.003 [DOI] [PubMed] [Google Scholar]
- 44.Title AC, Denzler R, Stoffel M. Uptake and function studies of maternal milk-derived MicroRNAs. J Biol Chem 2015; 290:23680-91; PMID:26240150; http://dx.doi.org/ 10.1074/jbc.M115.676734 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 45.Laubier J, Castille J, Le Guillou S, Le Provost F. No effect of an elevated miR-30b level in mouse milk on its level in pup tissues. RNA Biol 2015; 12:26-9; PMID:25763824; http://dx.doi.org/ 10.1080/15476286.2015.1017212 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 46.Waser NM, Ollerton J, editors Plant-pollinator interactions: From specialization to generalization. University of Chicago Press 2006; 98(4): 899-900 [Google Scholar]
- 47.Zhou M, Luo H. MicroRNA-mediated gene regulation: potential applications for plant genetic engineering. Plant Mol Biol 2013; 83:59-75; PMID:23771582; http://dx.doi.org/ 10.1007/s11103-013-0089-1 [DOI] [PubMed] [Google Scholar]
- 48.Palli SR. RNA interference in Colorado potato beetle: steps toward development of dsRNA as a commercial insecticide. Curr Opin Insect Sci 2014; 6:1-8; PMID:26705514; http://dx.doi.org/19591667 10.1016/j.cois.2014.09.011 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 49.Chambers C, Shuai B. Profiling microRNA expression in Arabidopsis pollen using microRNA array and real-time PCR. BMC Plant Biol 2009; 9:87; PMID:19591667; http://dx.doi.org/ 10.1186/1471-2229-9-87 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 50.Grant-Downton R, Le Trionnaire G, Schmid R, Rodriguez-Enriquez J, Hafidh S, Mehdi S, Twell D, Dickinson H. MicroRNA and tasiRNA diversity in mature pollen of Arabidopsis thaliana. BMC Genomics 2009; 10:643; PMID:20042113; http://dx.doi.org/ 10.1186/1471-2164-10-643 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 51.Li XM, Sang YL, Zhao XY, Zhang XS. High-throughput sequencing of small RNAs from pollen and silk and characterization of miRNAs as candidate factors involved in pollen-silk interactions in maize. PLoS One 2013; 8:e72852; PMID:23991159; http://dx.doi.org/ 10.1371/journal.pone.0072852 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 52.Jefferson JM, Dolstad HA, Sivalingam MD, Snow JW. Barrier immune effectors are maintained during transition from nurse to forager in the honey bee. PLoS One 2013; 8:e54097; PMID:23320121; http://dx.doi.org/ 10.1371/journal.pone.0054097 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 53.Nunes FMF, Ihle KE, Mutti NS, Simoes ZLP, Amdam GV. The gene vitellogenin affects microRNA regulation in honey bee (Apis mellifera) fat body and brain. J Exp Biol 2013; 216:3724-32; PMID:23788711; http://dx.doi.org/ 10.1242/jeb.089243 [DOI] [PubMed] [Google Scholar]
- 54.Esslinger SM, Schwalb B, Helfer S, Michalik KM, Witte H, Maier KC, Martin D, Michalke B, Tresch A, Cramer P, et al.. DrosophilamiR-277 controls branched-chain amino acid catabolism and affects lifespan. RNA Biol 2014; 10:1042-56; PMID:23669073; http://dx.doi.org/20119486 10.4161/rna.24810 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 55.Johnson BR. Division of labor in honeybees: form, function, and proximate mechanisms. Behav Ecol Sociobiol 2010; 64:305-16; PMID:20119486; http://dx.doi.org/ 10.1007/s00265-009-0874-7 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 56.Peng YS, Nasr ME, Marston JM. The digestion of dandelion pollen by adult worker honeybees. Physiol Entomol 1985; 10:75-82; http://dx.doi.org/ 10.1111/j.1365-3032.1985.tb00021.x [DOI] [Google Scholar]
- 57.Crailsheim K, Schneider L, Hrassnigg N, Buhlmann G, Brosch U, Gmeinbauer R, Schoffmann B. Pollen consumption and utilization in worker honeybees (Apis mellifera carnica): dependence on individual age and function. J Insect Physiol 1992; 38:409-19; http://dx.doi.org/ 10.1016/0022-1910(92)90117-V [DOI] [Google Scholar]
- 58.Marshall DL, Avritt JJ, Maliakal-Witt S, Medeiros JS, Shaner MGM. The impact of plant and flower age on mating patterns. Ann Bot 2010; 105:7-22; PMID:19875519; http://dx.doi.org/ 10.1093/aob/mcp260 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 59.Yeamans RL, Roulston TH, Carr DE. Pollen quality for pollinators tracks pollen quality for plants in Mimulus guttatus. Ecosphere 2014; 5 :Art91,1-8; http://dx.doi.org/ 10.1890/ES14-00099.1 [DOI] [Google Scholar]
- 60.Bellés X. Beyond Drosophila: RNAi in vivo and functional genomics in insects. Annu Rev Entomol 2010; 55:111-28; PMID:19961326; http://dx.doi.org/ 10.1146/annurev-ento-112408-085301 [DOI] [PubMed] [Google Scholar]
- 61.Wynant N, Santos D, Vanden Broeck J. Biological mechanisms determining the success of RNA interference in insects. Int Rev Cell Mol Biol 2014; 312:139-67; PMID:25262241; http://dx.doi.org/ 10.1016/B978-0-12-800178-3.00005-1 [DOI] [PubMed] [Google Scholar]
- 62.Aronstein K, PANKIWI T, Saldivar E. SID-I is implicated in systemic gene silencing in the honey bee. J Apicult Res 2006; 45:20-4 [Google Scholar]
- 63.Guo X, Su S, Skogerboe G, Dai S, Li W, Li Z, Liu F, Ni R, Guo Y, Chen S, et al.. Recipe for a busy bee: microRNAs in Honey Bee caste determination. PLoS One 2013; 8:e81661; PMID:24349106; http://dx.doi.org/ 10.1371/journal.pone.0081661 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 64.Shi YY, Wu XB, Huang ZY, Wang ZL, Yan WY, Zeng ZJ. Epigenetic modification of gene expression in honey bees by heterospecific gland secretions. PLoS One 2012; 7:e43727; PMID:22928024; http://dx.doi.org/ 10.1371/journal.pone.0043727 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 65.Huvenne H, Smagghe G. Mechanisms of dsRNA uptake in insects and potential of RNAi for pest control: a review. J Insect Physiol 2010; 56:227-35; PMID:19837076; http://dx.doi.org/ 10.1016/j.jinsphys.2009.10.004 [DOI] [PubMed] [Google Scholar]
- 66.Moritz B, Crailsheim K. Physiology of protein digestion in the midgut of the honeybee (Apis-Mellifera L). J Insect Physiol 1987; 33:923-31; http://dx.doi.org/ 10.1016/0022-1910(87)90004-7 [DOI] [Google Scholar]
- 67.Timmons L, Fire A. Specific interference by ingested dsRNA. Nature 1998; 395:854; PMID:9804418; http://dx.doi.org/ 10.1038/27579 [DOI] [PubMed] [Google Scholar]
- 68.Zhang J, Khan SA, Hasse C, Ruf S, Heckel DG, Bock R. Pest control. Full crop protection from an insect pest by expression of long double-stranded RNAs in plastids. Science 2015; 347:991-4; PMID:25722411; http://dx.doi.org/ 10.1126/science.1261680 [DOI] [PubMed] [Google Scholar]
- 69.Patel A, Fondrk MK, Kaftanoglu O, Emore C, Hunt G, Frederick K, Amdam GV. The making of a queen: TOR pathway is a key player in diphenic caste development. PLoS One 2007; 2:e509, 1-7; PMID:17551589 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 70.Nunes FMF, Simões ZLP. A non-invasive method for silencing gene transcription in honeybees maintained under natural conditions. Insect Biochem Molec 2009; 39:157-60; PMID:19049870; http://dx.doi.org/ 10.1016/j.ibmb.2008.10.011 [DOI] [PubMed] [Google Scholar]
- 71.Maori E, Paldi N, Shafir S, Kalev H, Tsur E, Glick E, Sela I. IAPV, a bee-affecting virus associated with Colony Collapse Disorder can be silenced by dsRNA ingestion. Insect Mol Biol 2009; 18:55-60; PMID:19196347; http://dx.doi.org/ 10.1111/j.1365-2583.2009.00847.x [DOI] [PubMed] [Google Scholar]
- 72.Hunter W, Ellis J, Hayes J, Westervelt D, Glick E, Williams M, Sela I, Maori E, Pettis J, Cox-Foster D. Large-scale field application of RNAi technology reducing Israeli acute paralysis virus disease in honey bees (Apis mellifera, hymenoptera: Apidae). PLoS Pathog 2010; 6:e1001160; PMID:21203478; http://dx.doi.org/ 10.1371/journal.ppat.1001160 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 73.Mutti NS, Wang Y, Kaftanoglu O, Amdam GV. Honey bee PTEN–description, developmental knockdown, and tissue-specific expression of splice-variants correlated with alternative social phenotypes. PLoS One 2011; 6:e22195; PMID:21779392; http://dx.doi.org/ 10.1371/journal.pone.0022195 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 74.Jarosch A, Stolle E, Crewe RM, Moritz RFA. Alternative splicing of a single transcription factor drives selfish reproductive behavior in honeybee workers (Apis mellifera). Proc Natl Acad Sci USA 2011; 108:15282-7; PMID:21896748; http://dx.doi.org/ 10.1073/pnas.1109343108 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 75.Nazzi F, Brown SP, Annoscia D, Del Piccolo F, Di Prisco G, Varricchio P, Vedova Della G, Cattonaro F, Caprio E, Pennacchio F. Synergistic parasite-pathogen interactions mediated by host immunity can drive the collapse of honeybee colonies. PLoS Pathog 2012; 8:e1002735; PMID:22719246; http://dx.doi.org/ 10.1371/journal.ppat.1002735 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 76.Desai SD, Eu YJ, Whyard S, Currie RW. Reduction in deformed wing virus infection in larval and adult honey bees (Apis mellifera L.) by double-stranded RNA ingestion. Insect Mol Biol 2012; 21:446-55; PMID:22690671; http://dx.doi.org/ 10.1111/j.1365-2583.2012.01150.x [DOI] [PubMed] [Google Scholar]
- 77.Beye M, Härtel S, Hagen A, Hasselmann M, Omholt S. Specific developmental gene silencing in the honey bee using a homeobox motif. Insect Mol Biol 2002; 11:527-32; PMID:12421410; http://dx.doi.org/ 10.1046/j.1365-2583.2002.00361.x [DOI] [PubMed] [Google Scholar]
- 78.Amdam GV, Simões ZLP, Guidugli KR, Norberg K, Omholt SW. Disruption of vitellogenin gene function in adult honeybees by intra-abdominal injection of double-stranded RNA. BMC Biotechnol 2003; 3:1; PMID:12546706; http://dx.doi.org/ 10.1186/1472-6750-3-1 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 79.Aronstein K, Saldivar E. Characterization of a honey bee Toll related receptor gene Am18w and its potential involvement in antimicrobial immune defense. Apidologie 2005; 36:3-14; http://dx.doi.org/ 10.1051/apido:2004062 [DOI] [Google Scholar]
- 80.Seehuus S-C, Norberg K, Gimsa U, Krekling T, Amdam GV. Reproductive protein protects functionally sterile honey bee workers from oxidative stress. Proc Natl Acad Sci USA 2006; 103:962-7; PMID:16418279; http://dx.doi.org/ 10.1073/pnas.0502681103 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 81.Maleszka J, Foret S, Saint R, Maleszka R. RNAi-induced phenotypes suggest a novel role for a chemosensory protein CSP5 in the development of embryonic integument in the honeybee (Apis mellifera). Dev Genes Evol 2007; 217:189-96; PMID:17216269; http://dx.doi.org/ 10.1007/s00427-006-0127-y [DOI] [PubMed] [Google Scholar]
- 82.Wang Y, Mutti NS, Ihle KE, Siegel A, Dolezal AG, Kaftanoglu O, Amdam GV. Down-regulation of honey bee IRS gene biases behavior toward food rich in protein. PLoS Genet 2010; 6:e1000896; PMID:20369023; http://dx.doi.org/ 10.1371/journal.pgen.1000896 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 83.Mustard JA, Pham PM, Smith BH. Modulation of motor behavior by dopamine and the D1-like dopamine receptor AmDOP2 in the honey bee. J Insect Physiol 2010; 56:422-30; PMID:19945462; http://dx.doi.org/ 10.1016/j.jinsphys.2009.11.018 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 84.Jarosch A, Moritz RFA. Systemic RNA-interference in the honeybee Apis mellifera: tissue dependent uptake of fluorescent siRNA after intra-abdominal application observed by laser-scanning microscopy. J Insect Physiol 2011; 57:851-7; PMID:21439290; http://dx.doi.org/ 10.1016/j.jinsphys.2011.03.013 [DOI] [PubMed] [Google Scholar]
- 85.Jarosch A, Moritz RF. RNA interference in honeybees: off-target effects caused by dsRNA. Apidologie 2012; 43:128-38; http://dx.doi.org/ 10.1007/s13592-011-0092-y [DOI] [Google Scholar]
- 86.Paldi N, Glick E, Oliva M, Zilberberg Y, Aubin L, Pettis J, Chen Y, Evans J. Effective gene silencing in a microsporidian parasite associated with honeybee (Apis mellifera) colony declines. Appl Environ Microbiol 2010; 76:5960; PMID:20622131; http://dx.doi.org/ 10.1128/AEM.01067-10 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 87.Jayachandran B, Hussain M, Asgari S. An insect trypsin-like serine protease as a target of microRNA: utilization of microRNA mimics and inhibitors by oral feeding. Insect Biochem Molec 2013; 43:398-406; PMID:23108205; http://dx.doi.org/ 10.1016/j.ibmb.2012.10.004 [DOI] [PubMed] [Google Scholar]
- 88.Axtell MJ. Classification and Comparison of Small RNAs from Plants. Annu Rev Plant Biol 2013; 64:137-59; PMID:23330790; http://dx.doi.org/ 10.1146/annurev-arplant-050312-120043 [DOI] [PubMed] [Google Scholar]
- 89.Brown BD, Naldini L. Exploiting and antagonizing microRNA regulation for therapeutic and experimental applications. Nat Rev Genet 2009; 10:578-85; PMID:19609263; http://dx.doi.org/ 10.1038/nrg2628 [DOI] [PubMed] [Google Scholar]
- 90.Goulson D, Nicholls E, Botías C, Rotheray EL. Bee declines driven by combined stress from parasites, pesticides, and lack of flowers. Science 2015; 347:1255957; PMID:25721506; http://dx.doi.org/ 10.1126/science.1255957 [DOI] [PubMed] [Google Scholar]
- 91.Chen S, Yao H, Han J, Liu C, Song J, Shi L, Zhu Y, Ma X, Gao T, Pang X, et al.. Validation of the ITS2 region as a novel DNA barcode for identifying medicinal plant species. PLoS One 2010; 5:e8613; PMID:20062805; http://dx.doi.org/ 10.1371/journal.pone.0008613 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 92.Koetschan C, Forster F, Keller A, Schleicher T, Ruderisch B, Schwarz R, Muller T, Wolf M, Schultz J. The ITS2 Database III–sequences and structures for phylogeny. Nucl Acids Res 2009; 38:D275-9; PMID:19920122; http://dx.doi.org/ 10.1093/nar/gkp966 [DOI] [PMC free article] [PubMed] [Google Scholar]
Associated Data
This section collects any data citations, data availability statements, or supplementary materials included in this article.