

Abstract
Objective:
This overview seeked to bring together the microRNA (miRNA) researches on biogenesis and bio-function in these areas of clinical diagnosis and therapy for malignant glioma.
Data Sources:
Using the keyword terms “glioma” and “miRNA,” we performed the literature search in PubMed, Ovid, and web.metstr.com databases from their inception to October 2014.
Study Selection:
In screening out the quality of the articles, factors such as clinical setting of the study, the size of clinical samples were taken into consideration. Animal studied for verification and reviews article were also included in our data collection.
Results:
Despite many advance in miRNA for malignant glioma, further studies were still required to focus on the following aspects: (i) Improving the understanding about biogenesis of miRNA and up-down regulation; (ii) utilizing high-throughput miRNA expression analysis to screen out the core miRNA for glioma; (iii) Focusing related miRNAs on the signal transduction pathways that regulate the proliferation and growth of glioma.
Conclusions:
We discussed the most promising miRNA, correlative signaling pathway and their relation with gliomas in the way of prompting miRNA target into being a clinical therapeutic strategy.
Keywords: Glioma, Malignant, MicroRNA, Pathway, Target, Treatment
INTRODUCTION
Malignant gliomas, as the most common and aggressive brain tumors in children and adults, present a dismal prognosis with an overall 5-year survival of 10%. It remains one of the greatest challenges to get optimal treatment formula. Novel gene expression profiling for addressing molecular characterization of brain tumors has pointed to genetic and epigenetic alterations. MicroRNA (miRNA) expression profiling has been reported to affect the function of genes involved in gliomagenesis, tumor growth, proliferation, apoptosis and posttranscriptional regulation of anti-oncogenes.[1,2] As a result, it is suggestive that miRNA profiling may be an available way to identify glioma origin, prognosis, and response to therapy. As a promising therapeutic tool against malignant glioma, miRNAs attract increasing attention. In this review, we will discuss the most promising miRNA, correlative signaling pathway and their relation with gliomas, as well as existing troubles to prompt miRNA into being a clinical therapeutic strategy for malignant glioma.
BIOGENESIS OF MICRORNA
MiRNAs are short (~22 nucleotides), single-stranded noncoding RNAs, although a minority is generated from intronic areas.[3,4] Since the first miRNAs found in Caenorhabditis elegans, the total number of miRNAs is exponentially rising. The original miRNA Database, generated in 2002 boosted their development of potential targets.[5] The biogenesis of miRNAs is under tight temporal and spatial control, and their dysregulation is associated with somatic stem cell proliferation, even tumorogenesis.[6,7]
Regarding miRNA biogenesis, RNA polymerase II or III transcribes miRNA genes into a long transcript, named primary microRNA (pri-miRNA). Thereafter, the nuclear enzyme crops pri-miRNA into a nucleotide hairpin-like precursor (pre-miRNA). Pre-miRNA is cleaved by an endoribonuclease (Dicer) to form a miRNA duplex after the transportation of pre-miRNA by exportin-5 and RanGTP. The miRNA duplex is unwound, and miRNP (miRNA containing ribonucleoprotein) is assembled from mature miRNA and Argonaut protein. MiRNP as the effector complex manipulates translational inhibition through imperfect complementary base-pairing to untranslated regions (UTRs) of their target mRNA or degradation. The precise mechanism underlying miRNA's regulation of gene expression is partly outlined [Figure 1]. It has been estimated that half of miRNA genes are located in cancer-associated genes or fragile sites.[8] It is believed that these miRNAs regulate 3% of whole genes and up to 30% of coding genes.[9,10] Furthermore, a single miRNA can influence up to 100 mRNAs simultaneously, and a single target can be regulated by one or more miRNAs.[1]
Figure 1.
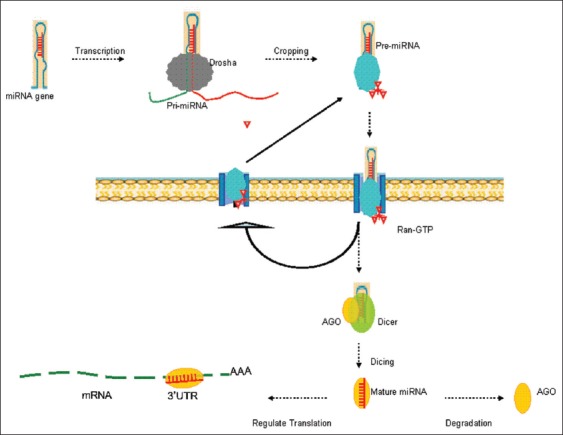
MicroRNA (miRNA) biogenesis is a multistep process: Transcription, Cropping and Dicing (cytoplasm). Primary microRNA (Pri-miRNA): Polyadenilated RNA transcripts; pre-miRNA: The hairpin-loop RNAs; mature miRNA: One strand of the duplex remains on the argonaute proteins (AGO). Pre-miRNA is recognized by the Exportin/Ran-GTP complex transporter and exported to the cytoplasm. Dicer and AGO mediate the process of pre-miRNA. Drosha: The nuclear enzyme of the RNase III family; AGO: Argonaute proteins.
When miRNA expression is amplified in cancer cells, it is considered to function as an oncogene. Conversely, when downregulated, it might act as a tumor suppressor. These alterations of miRNA expression contribute to the cancer phenotype, and further affect the clinic pathological appearance and prognosis. Recent studies have shown that miRNA expression signatures contribute to genetic determination of tumor subtypes and maintenance of the intra-tumoral heterogeneity.[11,12] MiRNA expression varies among different tissue samples and micro-environment states. As a consequence, it has been used as biomarkers of opposite biological states such as cancerous versus normal tissues. More and more data of glioma suggest that miRNA profiling can serve as a trustworthy formula for glioma diagnosis and prognosis.
MICRORNAS AND GLIOMA GENESIS
Recently, microarray studies on miRNAs have shown significant changes of expression in malignant gliomas.[13,14,15,16,17,18,19,20,21,22,23,24,25,26,27,28,29,30,31] The most of over-expressed miRNAs in tumors were involved in propagation processes of glioma. They seemed to act as an oncogene that was implicated in malignant transformation processes of glioma. Herein, we will describe the clinical or experimental utilization of oncogenic miRNAs to identify a series of potential therapeutic target for malignant glioma [Table 1].
Table 1.
Expression of miRNAs and potential relative signaling pathway in malignant glioma
| miRNA | Level of expression | Reference | Signaling pathway |
|---|---|---|---|
| MiR21 | Up-regulation | 13,18 | TGF, STAT3 |
| MiR199b | Up-regulation | 21 | Notch1 |
| MiR203 | Up-regulation | 22 | Cell cycle |
| MiR221/222 | Up-regulation | 18,25 | AKT, apoptosis |
| MiR26b | Up-regulation | 28 | AKT |
| MiR182 | Up-regulation | 42 | / |
| MiR10b | Up-regulation | 27 | AKT |
| MiR181 | Down-regulation | 17,18,29 | Calcium signaling pathway |
| MiR128 | Down-regulation | 19,20 | P53, RTK signaling, mTOR |
| MiR153 | Down-regulation | 47 | / |
| MiR34a | Down-regulation | 23 | P53 pathway, Notch |
| MiR451 | Down-regulation | 50 | AKT, AMPK |
| MiR146b | Down-regulation | 31 | / |
| MiR17 | Down-regulation | 50 | / |
| MiR184 | Down-regulation | 53 | / |
| MiR326 | Down-regulation | 61 | Notch |
| MiR7 | Down-regulation | 24 | AKT, EGFR |
TFG: Transforming growth factor; EGFR: Epidermal growth-factor receptor; miRNA: Micro RNA.
MiR-21 is the most typical example with overexpression and associated with poor prognosis of gliomas.[32,33] Investigation revealed high expression of miR-21 in most of the malignant gliomas versus normal brain. Qu et al. found the higher level of miRNA21 level in serum of patients with higher grade malignant glioma than the control group. It was suggestive that miRNA21 might be a potential diagnostic biomarker for human malignant glioma. The recent meta-analysis of five studies indicated that a single-miR21 as one of the most representative miRNAs could be a powerful clinical biomarker in glioma diagnosis.[34] In accordance, β-catenin has been proposed as an indirect target for miR-21. Knockdown of miR21 inhibited cell invasion by β-catenin regulation via STAT3 pathway.[35] Recent trial demonstrated that miR-21 inhibitor therapy could enhance chemosensitivity of glioma resistant cells to anthracyclines by inducing apoptosis.[36] Expression of miR-199b-5p has been found an impressing divergence in nonmetastatic patients with medulloblastoma (MB) compared to metastatic ones. Over-expression of miR-199b-5p might inhibit the expression of cancer stem cell genes and be further correlated with the corresponding prognosis.[21,37] MiR-203 in ependymoma was recognized as an independent marker for predicting patients’ relapse.[22] MiR-203 could inhibit the proliferation and invasion of glioma cells via suppressing the protein expression of PLD2, which demonstrated the clinical significance of miR-203 in gliomas.[38] MiR-221/222 was up-regulated in high-grade astrocytomas with analogous target specificity and increased cell invasion and poor prognosis in glioma.[39] Bioinformatic analysis reveals that miR221/222 had the potential to regulate about 70 common target genes and exert cooperative effect on the function via AKT pathway.[40] Furthermore, miR-221/222 directionally down-regulated the 3’UTR of the cell cycle regulator and p27kip1.[40] Le Sage et al. demonstrated that over-expression of miR-221/222 regulated cell proliferation through maintaining low levels of p27 kip1.[25] MiR-26a was frequently amplified in human gliomas and was associated with monoallelic PTEN loss.[28] Furthermore, miR-26a-mediated PTEN repression in a mouse model enhanced de novo tumor formation. Qian's investigation revealed that miR26a regulated its direct target prohibitin and promoted glioma progression.[41] Jiang et al. found that miR-182 was up-regulated in primary glioma specimens versus normal brain. MiR-182 expression level in the tumors was statistically correlated with tumor grade and clinical features.[42] It was suggestive that miR-182 is a prognostic marker for glioma progression and patient survival. Sasayama revealed high expression of miR-10b in glioma samples vs. nonneoplastic tissues and the association between its expression level and grade malignancy in glioma.[27] Liu's investigation reported that transforming growth factor-β (TGF-β)-induced miR10a/10b promoted glioma migration through the suppression of PTEN.[43] Given all that these researches provided a new insight of developing miRNA-based diagnosis and targeting treatment for glioma.
On the other side of the same coin, several miRNAs with suppressive expression have been identified in brain gliomas. For example, Conti A performed the analysis of miRNA expression pattern in gliomas at different stages of malignancy and suggested that miR-181 seemed to become into a criteria between high-grade and low-grade gliomas. In biological, the family of miRNA181 with low expression was associated with the processes of glioma progression in different manners.[17,18] According to Chen’ study, B-cell lymphoma-2 (Bcl-2) was a potential target of miR-181a for enhancing the effect of radiation treatment on malignant glioma cells. Present study demonstrated that aplysin was believed to enhance the effect of temozolomide on glioma cells by increasing miR181 expression.[44] At the molecular level, expression of cyclin B1 (a positive cell cycle regulator) could be negatively regulated by miR181, which inhibited glioma cell proliferation.[45] Furthermore, miR-181c could enhance the effect of miR-21 as an anticancer target against glioma. Down-regulation of miR-128 was prevailing in high-grade gliomas and the plasma levels of miR-128 with significant alteration in glioblastoma multiforme (GBM) patients could discriminate glioma from a healthy population with high specificity and sensitivity.[46] Down-regulation of miR-128 was correlated with increased proliferation and poor differentiation of glioma cells, and vice versa.[19,20] MiR-153 inhibited Bcl-2 and myeloid cell leukemia sequence-1) expressions by directly targeting the 3’UTR regions of their respective mRNAs. It was therefore considered that miR-153 exerted pro-apoptotic effects in gliomas.[47] Transient transfection of miR-153 into glioblastoma stem cells could impair their self-renewal ability and induce their differentiation. Meanwhile, miR-153 could also repress tumor stem cell growth and induce apoptosis.[48] These studies indicated that reactivation of miR-153 expression suggested novel therapeutic strategies for malignant gliomas. Glioma patients further demonstrated that miR-34a expression levels positively correlated with tumor malignancy grades and lower expression levels correlated with worse progression-free survival and overall survival.[49] It was suggested that miR-34a expression could be potential useful signature for predicting prognosis of glioma. MiR-34a was shown a positive correlation with p53 in MB and GBM samples. Loss of miR-34a presents a mechanism of down-regulating p53 pathway through modulation of melanoma associated antigens (MAGE-A) genes. It was considered that expression levels of miR-34a and p53 in MB were associated with drug resistance and chemosensitivity.[23] MiR-451 was considered as a tumor suppressor function in human gliomas and played an important role in tumor growth, invasion and apoptosis. Down-regulation of miR-451 might conditionally activated AMPK signaling pathway and therefore achieved the increased cell survival.[50] MiR-451 expression was down-regulated in glioma samples and was inversely correlated with malignancy grades of glioma. Furthermore, miR-451 had tumor suppressive traits and could repress glioma progression through targeting CAB39 directly and inhibiting AKT pathway indirectly.[51] Using microarrays, miR-146b was significantly down-regulated in human glioblastoma tissue. Alteration of miR-146b expression might reduce the migration and invasion of glioma cells.[31] Recent study revealed that decreased miR-146b-5p expression was strongly correlated with chromosome 10q loss in malignant gliomas. The over-expression miR-146b-5p in glioma cells led to the inhibition of tumor cell migration and invasion.[52] Reduced expression of miR-17 and miR-184 was identified between Grade IV secondary gliomas and corresponding Grade II primary gliomas. Both of the miRNAs act a potential role of inhibiting cell viability, proliferation, and invasion.[53]
MICRORNA AS A TARGET IN SIGNALING PATHWAYS
The most important pathways regulating glioma cell proliferation include the epidermal growth factor receptor (EGFR) and PI3K/AKT signaling pathways, p53 pathway, TGF-dependent apoptotic signaling cascade, as well as the Notch pathway and nuclear Factor-κB (NF-κB) pathway.[54] MiRNAs were linked with the regulation of all above-mentioned pathways suggesting their intimate implication in different phenotypes of malignant gliomas.[55]
To date, bio-informational analysis revealed that miR-21 was associated with TGF-β signaling pathway in GBM.[56] In addition, inhibition of miR-21 with antisense oligonucleotides resulted in suppression of EGFR signaling pathway and regulation of glioma proliferation.[57] Furthermore, leucine rich repeat interacting protein 1 acted as a direct target for miR-21 and could directly inhibit NF-κB signaling pathway, thus disrupting invasiveness and migration of gliomas.[58] MiR-221/222 in previous literature was found to regulate several genes involved in AKT signaling pathways and the pro-apoptotic gene P53 upregulated modulator of apoptosis.[15,39] In addition, a recent investigation revealed that chronic miR221/222 mediated MGMT down-regulation could render glioma cells unable to repair genetic damage.[59] The result indicated that the target of miR221/222 pointed to MGMT of DNA repairment in the process of gliomagenesis. It was suggestive that the direct/indirect targets of miR-128 covered the angiopoietin-related protein (5ARP5), p53, Bmi-1, and the cell cycle gene activator (E2F-3a). The functions of these genes were involved in the regulation of cell renewal, proliferation, differentiation, and tumorigenesis.[19,20] In addition, miR-128 might act on caspase-mediated apoptosis and suppress tumor growth. MiR-451 was attributed as a tumor suppressor function targeting of the AKT and AMPK signaling pathway. Down-regulation of miR-451 was suggested to have an impact in cell growth, migration, invasion, and apoptosis. Based on a multidimensional genomic data set of glioblastoma from Tumor Cancer Genome Atlas (TCGA) datasets, miR-26a was validated to be a cooperating component of a frequently occurring amplicon, where contained oncogenes CENTG1 that regulate AKT pathways. By integrating DNA copy number, mRNA, miRNA, and DNA methylation data, the clinic further investigated several potential targets of miR-26a in glioma, including PTEN, RB1, and MAP3K2/MEKK2.[60] MiR-326 was down-regulated in gliomas and was identified as a direct regulator of Notch expression in glioblastoma stem cells.[61] Similar in kind, miR-7 (with down-regulation in tumor samples) reduced the viability and invasiveness of glioblastoma cells by directly targeting EGFR and AKT pathway.[24]
DIAGNOSTIC AND PROGNOSTIC VALUE OF MULTI-MICRORNA SIGNATURE
The heterogeneous and unpredictable nature of malignant glioma has attracted scientists’ attention on molecular markers that might complement the examination of brain tumor pathology. At the moment, miRNA isolation and detection was no longer an obstacle. In addition, miRNAs could also be detected in formalin-fixed paraffin-embedded tissues, serum, cerebrospinal fluid, plasma, and urine. MiRNA-based q-real-time polymerase chain reaction diagnostic test afforded an accurate classification of brain tumors with a remarkable sensitivity and specificity of 84% and 95%, respectively.[62] Skog's study shed a bright light on detection of exosomes which was a small complex containing miRNAs and proteins. As a result, miRNA profiling of tumors and normal tissues improved to become an important instrument to classify, diagnose, and predict the outcome.[63] Ongoing clinical trials aimed to identify certain miRNAs with potential prognostic capacity in blood samples and/or cerebrospinal fluid of glioma patients during the course of treatment.
For example, Srinivasan's study revealed that a multi-miRNA expression signature successfully distinguished malignancy degrades of gliomas with a remarkable sensitivity and specificity.[64] In addition, another study from Lages's team presented seven dysregulated miRNAs that allow discrimination of oligodendrogliomas from glioblastomas.[65] Hayes et al. identified a nine-miRNA signature that stratified high- and low-risk patients in GBM. In addition, the majority of nine miRNAs have been previously linked to malignant glioma biology or treatment response.[66] Further miRNA-mRNA interaction within the nine-miRNA signature revealed a relevant regulatory network. Barbano R explored and identified 22 miRNAs distinguishing Grade II gliomas from malignant gliomas by array analysis. A six-miRNA signature of those miRNAs demonstrated a significant association with prognosis based on the TCGA datasets.[33] MiR-196a with up-regulation in higher grade gliomas was positively associated with poorer prognosis. Multivariate analysis revealed that miR-196 was an independent predictor of overall survival in glioblastoma patients.[67] Recent research from malignant gloma samples developed a five-miRNA signature (including risky miR-196a) that could identify patients with a high risk of an unfavorable outcome in anaplastic gliomas regardless of histology type.[68] These results echoed the hypothesis that specific multi-miRNA signature served as a prognostic marker for patient risk stratification in malignant gliomas.
CURRENT CHALLENGES
MicroRNA-based target therapies have proven as an attractive approach. Despite the large progress in the field, many challenges are still ahead. One of the biggest obstacles is the identification of definitive single- and multi-miRNA signature for malignant glioma by large and comprehensive profiling studies. These results will develop a set of diagnostic and prognostic biomarkers for clinical evaluation and therapy. Another emerging theme in miRNA cancer research relates to the ability of a single-miRNA to target multiple effectors. Therefore, miRNA target specificity is considered as the rate-limiting step for miRNA-targeting therapy against malignant gliomas. How to improve the specificity of miRNA targeting effectors? In the process of biogenesis, miRNAs primary function as posttranscriptional negative regulators of gene expression through binding to their mRNA targets [Figure 1]. The integrative analysis of miRNA and mRNA expression profiling may facilitate the identification of a miR's targets by considerable bioinformatic algorithms. The miRNA-mRNA interactions from the large scale of datasets may be reduced to several relevant signaling pathways, through which miRNAs regulate the progression of malignant gliomas [Figure 2]. According to multi-miRNA signature and corresponding mRNA alteration for some subtype of gliomas, combined miRNA-based target therapy may allow a specific regulation on core signaling pathway.
Figure 2.
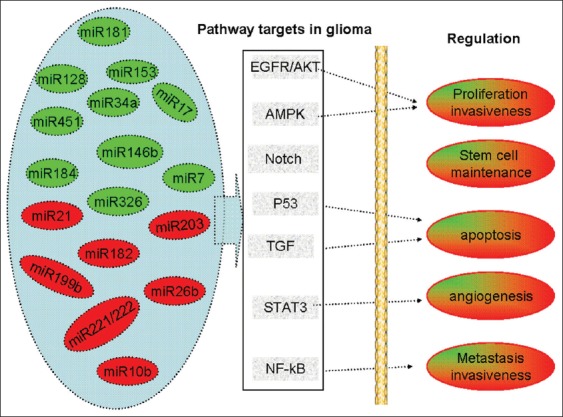
MicroRNAs with regulatory potential over major signaling pathways affected in gliomagenesis and impacted in cellular functions.
In conclusion, microRNAs beneficially or not regulate tumor cell proliferation, apoptosis, stem cell maintenance, invasion, and angiogenesis by the reciprocal interaction with target mRNAs. However, the information of details in this process remains incomplete, and further research is necessary for a better understanding of the role of miRNAs in glioma cells. Minimally invasive approaches for miRNAs may be a promising tool of monitoring glioma progression. Before miRNA-based target therapy being translated into clinical practice, a large scale of clinical samples should be independently validated for core and promising candidate in miRNAs data. Consequently, core miRNAs should undertake further investigation in multicenter clinical trials. In the future, tailored treatment regimens should be framed by miRNA profiles and other genetic alterations.
Footnotes
Edited by: Li-Shao Guo
Source of Support: This work was supported by grants from the National Key Basic Research Program (No. NKBRP 2010CB945203), the National Natural Science Foundation of China (No. 81271382); the Program of Shanghai Subject Chief Scientist (No. 09XD1403300); Foundation of Shanghai Jiaotong University, School of Medicine (No. 14XJ10034); the Program of Shanghai Health and Family Planning Committee (No. 201440495); Shanghai Municipal Natural Science Foundation (No. 14ZR1427100).
Conflict of Interest: None declared.
REFERENCES
- 1.Sumazin P, Yang X, Chiu HS, Chung WJ, Iyer A, Llobet-Navas D, et al. An extensive microRNA-mediated network of RNA-RNA interactions regulates established oncogenic pathways in glioblastoma. Cell. 2011;147:370–81. doi: 10.1016/j.cell.2011.09.041. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 2.Riddick G, Fine HA. Integration and analysis of genome-scale data from gliomas. Nat Rev Neurol. 2011;7:439–50. doi: 10.1038/nrneurol.2011.100. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 3.Ha M, Kim VN. Regulation of microRNA biogenesis. Nat Rev Mol Cell Biol. 2014;15:509–24. doi: 10.1038/nrm3838. [DOI] [PubMed] [Google Scholar]
- 4.Wuchty S, Arjona D, Li A, Kotliarov Y, Walling J, Ahn S, et al. Prediction of Associations between microRNAs and gene expression in glioma biology. PLoS One. 2011;6:e14681. doi: 10.1371/journal.pone.0014681. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 5.Kozomara A, Griffiths-Jones S. miRBase: Integrating microRNA annotation and deep-sequencing data. Nucleic Acids Res. 2011;39:D152–7. doi: 10.1093/nar/gkq1027. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 6.Shenoy A, Blelloch RH. Regulation of microRNA function in somatic stem cell proliferation and differentiation. Nat Rev Mol Cell Biol. 2014;15:565–76. doi: 10.1038/nrm3854. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 7.Melo SA, Sugimoto H, O’Connell JT, Kato N, Villanueva A, Vidal A, et al. Cancer exosomes perform cell-independent microRNA biogenesis and promote tumorigenesis. Cancer Cell. 2014;26:707–21. doi: 10.1016/j.ccell.2014.09.005. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 8.Hummel R, Maurer J, Haier J. MicroRNAs in brain tumors: A new diagnostic and therapeutic perspective? Mol Neurobiol. 2011;44:223–34. doi: 10.1007/s12035-011-8197-x. [DOI] [PubMed] [Google Scholar]
- 9.González-Gómez P, Sánchez P, Mira H. MicroRNAs as regulators of neural stem cell-related pathways in glioblastoma multiforme. Mol Neurobiol. 2011;44:235–49. doi: 10.1007/s12035-011-8196-y. [DOI] [PubMed] [Google Scholar]
- 10.Berindan-Neagoe I, Monroig Pdel C, Pasculli B, Calin GA. MicroRNAome genome: A treasure for cancer diagnosis and therapy. CA Cancer J Clin. 2014;64:311–36. doi: 10.3322/caac.21244. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 11.Kumar RM, Cahan P, Shalek AK, Satija R, DaleyKeyser AJ, Li H, et al. Deconstructing transcriptional heterogeneity in pluripotent stem cells. Nature. 2014;516:56–61. doi: 10.1038/nature13920. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 12.Hu Z, Chen X, Zhao Y, Tian T, Jin G, Shu Y, et al. Serum microRNA signatures identified in a genome-wide serum microRNA expression profiling predict survival of non-small-cell lung cancer. J Clin Oncol. 2010;28:1721–6. doi: 10.1200/JCO.2009.24.9342. [DOI] [PubMed] [Google Scholar]
- 13.Chen Y, Liu W, Chao T, Zhang Y, Yan X, Gong Y, et al. MicroRNA-21 down-regulates the expression of tumor suppressor PDCD4 in human glioblastoma cell T98G. Cancer Lett. 2008;272:197–205. doi: 10.1016/j.canlet.2008.06.034. [DOI] [PubMed] [Google Scholar]
- 14.Zhou X, Ren Y, Moore L, Mei M, You Y, Xu P, et al. Downregulation of miR-21 inhibits EGFR pathway and suppresses the growth of human glioblastoma cells independent of PTEN status. Lab Invest. 2010;90:144–55. doi: 10.1038/labinvest.2009.126. [DOI] [PubMed] [Google Scholar]
- 15.Zhang J, Han L, Ge Y, Zhou X, Zhang A, Zhang C, et al. miR-221/222 promote malignant progression of glioma through activation of the Akt pathway. Int J Oncol. 2010;36:913–20. doi: 10.3892/ijo_00000570. [DOI] [PubMed] [Google Scholar]
- 16.Zhang CZ, Zhang JX, Zhang AL, Shi ZD, Han L, Jia ZF, et al. MiR-221 and miR-222 target PUMA to induce cell survival in glioblastoma. Mol Cancer. 2010;9:229. doi: 10.1186/1476-4598-9-229. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 17.Shi L, Cheng Z, Zhang J, Li R, Zhao P, Fu Z, et al. Hsa-mir-181a and hsa-mir-181b function as tumor suppressors in human glioma cells. Brain Res. 2008;1236:185–93. doi: 10.1016/j.brainres.2008.07.085. [DOI] [PubMed] [Google Scholar]
- 18.Novakova J, Slaby O, Vyzula R, Michalek J. MicroRNA involvement in glioblastoma pathogenesis. Biochem Biophys Res Commun. 2009;386:1–5. doi: 10.1016/j.bbrc.2009.06.034. [DOI] [PubMed] [Google Scholar]
- 19.Cui JG, Zhao Y, Sethi P, Li YY, Mahta A, Culicchia F, et al. Micro-RNA-128 (miRNA-128) down-regulation in glioblastoma targets ARP5 (ANGPTL6), Bmi-1 and E2F-3a, key regulators of brain cell proliferation. J Neurooncol. 2010;98:297–304. doi: 10.1007/s11060-009-0077-0. [DOI] [PubMed] [Google Scholar]
- 20.Zhang Y, Chao T, Li R, Liu W, Chen Y, Yan X, et al. MicroRNA-128 inhibits glioma cells proliferation by targeting transcription factor E2F3a. J Mol Med (Berl) 2009;87:43–51. doi: 10.1007/s00109-008-0403-6. [DOI] [PubMed] [Google Scholar]
- 21.Garzia L, Andolfo I, Cusanelli E, Marino N, Petrosino G, De Martino D, et al. MicroRNA-199b-5p impairs cancer stem cells through negative regulation of HES1 in medulloblastoma. PLoS One. 2009;4:e4998. doi: 10.1371/journal.pone.0004998. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 22.Costa FF, Bischof JM, Vanin EF, Lulla RR, Wang M, Sredni ST, et al. Identification of microRNAs as potential prognostic markers in ependymoma. PLoS One. 2011;6:e25114. doi: 10.1371/journal.pone.0025114. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 23.Weeraratne SD, Amani V, Neiss A, Teider N, Scott DK, Pomeroy SL, et al. miR-34a confers chemosensitivity through modulation of MAGE-A and p53 in medulloblastoma. Neuro Oncol. 2011;13:165–75. doi: 10.1093/neuonc/noq179. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 24.Kefas B, Godlewski J, Comeau L, Li Y, Abounader R, Hawkinson M, et al. microRNA-7 inhibits the epidermal growth factor receptor and the Akt pathway and is down-regulated in glioblastoma. Cancer Res. 2008;68:3566–72. doi: 10.1158/0008-5472.CAN-07-6639. [DOI] [PubMed] [Google Scholar]
- 25.le Sage C, Nagel R, Egan DA, Schrier M, Mesman E, Mangiola A, et al. Regulation of the p27(Kip1) tumor suppressor by miR-221 and miR-222 promotes cancer cell proliferation. EMBO J. 2007;26:3699–708. doi: 10.1038/sj.emboj.7601790. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 26.Kefas B, Comeau L, Floyd DH, Seleverstov O, Godlewski J, Schmittgen T, et al. The neuronal microRNA miR-326 acts in a feedback loop with notch and has therapeutic potential against brain tumors. J Neurosci. 2009;29:15161–8. doi: 10.1523/JNEUROSCI.4966-09.2009. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 27.Sasayama T, Nishihara M, Kondoh T, Hosoda K, Kohmura E. MicroRNA-10b is overexpressed in malignant glioma and associated with tumor invasive factors, uPAR and RhoC. Int J Cancer. 2009;125:1407–13. doi: 10.1002/ijc.24522. [DOI] [PubMed] [Google Scholar]
- 28.Huse JT, Brennan C, Hambardzumyan D, Wee B, Pena J, Rouhanifard SH, et al. The PTEN-regulating microRNA miR-26a is amplified in high-grade glioma and facilitates gliomagenesis in vivo. Genes Dev. 2009;23:1327–37. doi: 10.1101/gad.1777409. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 29.Lakomy R, Sana J, Hankeova S, Fadrus P, Kren L, Lzicarova E, et al. MiR-195, miR-196b, miR-181c, miR-21 expression levels and O-6-methylguanine-DNA methyltransferase methylation status are associated with clinical outcome in glioblastoma patients. Cancer Sci. 2011;102:2186–90. doi: 10.1111/j.1349-7006.2011.02092.x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 30.Gillies JK, Lorimer IA. Regulation of p27Kip1 by miRNA 221/222 in glioblastoma. Cell Cycle. 2007;6:2005–9. doi: 10.4161/cc.6.16.4526. [DOI] [PubMed] [Google Scholar]
- 31.Xia H, Qi Y, Ng SS, Chen X, Li D, Chen S, et al. MicroRNA-146b inhibits glioma cell migration and invasion by targeting MMPs. Brain Res. 2009;1269:158–65. doi: 10.1016/j.brainres.2009.02.037. [DOI] [PubMed] [Google Scholar]
- 32.Qian X, Long L, Shi Z, Liu C, Qiu M, Sheng J, et al. Star-branched amphiphilic PLA-b-PDMAEMA copolymers for co-delivery of miR-21 inhibitor and doxorubicin to treat glioma. Biomaterials. 2014;35:2322–35. doi: 10.1016/j.biomaterials.2013.11.039. [DOI] [PubMed] [Google Scholar]
- 33.Barbano R, Palumbo O, Pasculli B, Galasso M, Volinia S, D’Angelo V, et al. A miRNA signature for defining aggressive phenotype and prognosis in gliomas. PLoS One. 2014;9:e108950. doi: 10.1371/journal.pone.0108950. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 34.Qu S, Guan J, Liu Y. Identification of microRNAs as novel biomarkers for glioma detection: A meta-analysis based on 11 articles. J Neurol Sci. 2015;348:181–7. doi: 10.1016/j.jns.2014.11.036. [DOI] [PubMed] [Google Scholar]
- 35.Han L, Yue X, Zhou X, Lan FM, You G, Zhang W, et al. MicroRNA-21 expression is regulated by ß-catenin/STAT3 pathway and promotes glioma cell invasion by direct targeting RECK. CNS Neurosci Ther. 2012;18:573–83. doi: 10.1111/j.1755-5949.2012.00344.x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 36.Giunti L, da Ros M, Vinci S, Gelmini S, Iorio AL, Buccoliero AM, et al. Anti-miR21 oligonucleotide enhances chemosensitivity of T98G cell line to doxorubicin by inducing apoptosis. Am J Cancer Res. 2015;5:231–42. [PMC free article] [PubMed] [Google Scholar]
- 37.Andolfo I, Liguori L, De Antonellis P, Cusanelli E, Marinaro F, Pistollato F, et al. The micro-RNA 199b-5p regulatory circuit involves Hes1, CD15, and epigenetic modifications in medulloblastoma. Neuro Oncol. 2012;14:596–612. doi: 10.1093/neuonc/nos002. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 38.Chen Z, Li D, Cheng Q, Ma Z, Jiang B, Peng R, et al. MicroRNA-203 inhibits the proliferation and invasion of U251 glioblastoma cells by directly targeting PLD2. Mol Med Rep. 2014;9:503–8. doi: 10.3892/mmr.2013.1814. [DOI] [PubMed] [Google Scholar]
- 39.Zhang C, Zhang J, Hao J, Shi Z, Wang Y, Han L, et al. High level of miR-221/222 confers increased cell invasion and poor prognosis in glioma. J Transl Med. 2012;10:119. doi: 10.1186/1479-5876-10-119. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 40.Zhang J, Han L, Ge Y, Zhou X, Zhang A, Zhang C, et al. MiR-221/222 promote malignant progression of glioma through activation of the Akt pathway. Int J Oncol. 2010;36:913–20. doi: 10.3892/ijo_00000570. [DOI] [PubMed] [Google Scholar]
- 41.Qian X, Zhao P, Li W, Shi ZM, Wang L, Xu Q, et al. MicroRNA-26a promotes tumor growth and angiogenesis in glioma by directly targeting prohibitin. CNS Neurosci Ther. 2013;19:804–12. doi: 10.1111/cns.12149. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 42.Jiang L, Mao P, Song L, Wu J, Huang J, Lin C, et al. MiR-182 as a prognostic marker for glioma progression and patient survival. Am J Pathol. 2010;177:29–38. doi: 10.2353/ajpath.2010.090812. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 43.Liu S, Sun J, Lan Q. TGF-ß-induced miR10a/b expression promotes human glioma cell migration by targeting PTEN. Mol Med Rep. 2013;8:1741–6. doi: 10.3892/mmr.2013.1709. [DOI] [PubMed] [Google Scholar]
- 44.Gong A, Ge N, Yao W, Lu L, Liang H. Aplysin enhances temozolomide sensitivity in glioma cells by increasing miR-181 level. Cancer Chemother Pharmacol. 2014;74:531–8. doi: 10.1007/s00280-014-2534-5. [DOI] [PubMed] [Google Scholar]
- 45.Wang F, Sun JY, Zhu YH, Liu NT, Wu YF, Yu F. MicroRNA-181 inhibits glioma cell proliferation by targeting cyclin B1. Mol Med Rep. 2014;10:2160–4. doi: 10.3892/mmr.2014.2423. [DOI] [PubMed] [Google Scholar]
- 46.Wang Q, Li P, Li A, Jiang W, Wang H, Wang J, et al. Plasma specific miRNAs as predictive biomarkers for diagnosis and prognosis of glioma. J Exp Clin Cancer Res. 2012;31:97. doi: 10.1186/1756-9966-31-97. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 47.Xu J, Liao X, Wong C. Downregulations of B-cell lymphoma 2 and myeloid cell leukemia sequence 1 by microRNA 153 induce apoptosis in a glioblastoma cell line DBTRG-05MG. Int J Cancer. 2010;126:1029–35. doi: 10.1002/ijc.24823. [DOI] [PubMed] [Google Scholar]
- 48.Zhao S, Deng Y, Liu Y, Chen X, Yang G, Mu Y, et al. MicroRNA-153 is tumor suppressive in glioblastoma stem cells. Mol Biol Rep. 2013;40:2789–98. doi: 10.1007/s11033-012-2278-4. [DOI] [PubMed] [Google Scholar]
- 49.Gao H, Zhao H, Xiang W. Expression level of human miR-34a correlates with glioma grade and prognosis. J Neurooncol. 2013;113:221–8. doi: 10.1007/s11060-013-1119-1. [DOI] [PubMed] [Google Scholar]
- 50.Gal H, Pandi G, Kanner AA, Ram Z, Lithwick-Yanai G, Amariglio N, et al. MIR-451 and Imatinib mesylate inhibit tumor growth of Glioblastoma stem cells. Biochem Biophys Res Commun. 2008;376:86–90. doi: 10.1016/j.bbrc.2008.08.107. [DOI] [PubMed] [Google Scholar]
- 51.Tian Y, Nan Y, Han L, Zhang A, Wang G, Jia Z, et al. MicroRNA miR-451 downregulates the PI3K/AKT pathway through CAB39 in human glioma. Int J Oncol. 2012;40:1105–12. doi: 10.3892/ijo.2011.1306. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 52.Li Y, Wang Y, Yu L, Sun C, Cheng D, Yu S, et al. miR-146b-5p inhibits glioma migration and invasion by targeting MMP16. Cancer Lett. 2013;339:260–9. doi: 10.1016/j.canlet.2013.06.018. [DOI] [PubMed] [Google Scholar]
- 53.Malzkorn B, Wolter M, Liesenberg F, Grzendowski M, Stühler K, Meyer HE, et al. Identification and functional characterization of microRNAs involved in the malignant progression of gliomas. Brain Pathol. 2010;20:539–50. doi: 10.1111/j.1750-3639.2009.00328.x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 54.Huang Z, Cheng L, Guryanova OA, Wu Q, Bao S. Cancer stem cells in glioblastoma – molecular signaling and therapeutic targeting. Protein Cell. 2010;1:638–55. doi: 10.1007/s13238-010-0078-y. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 55.Sana J, Hajduch M, Michalek J, Vyzula R, Slaby O. MicroRNAs and glioblastoma: Roles in core signalling pathways and potential clinical implications. J Cell Mol Med. 2011;15:1636–44. doi: 10.1111/j.1582-4934.2011.01317.x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 56.Yu J, Cai X, He J, Zhao W, Wang Q, Liu B. Microarray-based analysis of gene regulation by transcription factors and microRNAs in glioma. Neurol Sci. 2013;34:1283–9. doi: 10.1007/s10072-012-1228-1. [DOI] [PubMed] [Google Scholar]
- 57.Zhang KL, Han L, Chen LY, Shi ZD, Yang M, Ren Y, et al. Blockage of a miR-21/EGFR regulatory feedback loop augments anti-EGFR therapy in glioblastomas. Cancer Lett. 2014;342:139–49. doi: 10.1016/j.canlet.2013.08.043. [DOI] [PubMed] [Google Scholar]
- 58.Li H, Jia Z, Li A, Jenkins G, Yang X, Hu J, et al. Resveratrol repressed viability of U251 cells by miR-21 inhibiting of NF-k pathway. Mol Cell Biochem. 2013;382:137–43. doi: 10.1007/s11010-013-1728-1. [DOI] [PubMed] [Google Scholar]
- 59.Quintavalle C, Mangani D, Roscigno G, Romano G, Diaz-Lagares A, Iaboni M, et al. MiR-221/222 target the DNA methyltransferase MGMT in glioma cells. PLoS One. 2013;8:e74466. doi: 10.1371/journal.pone.0074466. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 60.Kim H, Huang W, Jiang X, Pennicooke B, Park PJ, Johnson MD. Integrative genome analysis reveals an oncomir/oncogene cluster regulating glioblastoma survivorship. Proc Natl Acad Sci U S A. 2010;107:2183–8. doi: 10.1073/pnas.0909896107. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 61.Guessous F, Zhang Y, Kofman A, Catania A, Li Y, Schiff D, et al. MicroRNA-34a is tumor suppressive in brain tumors and glioma stem cells. Cell Cycle. 2010;9:1031–6. doi: 10.4161/cc.9.6.10987. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 62.Mueller WC, Spector Y, Edmonston TB, St Cyr B, Jaeger D, Lass U, et al. Accurate classification of metastatic brain tumors using a novel microRNA-based test. Oncologist. 2011;16:165–74. doi: 10.1634/theoncologist.2010-0305. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 63.Kim TM, Huang W, Park R, Park PJ, Johnson MD. A developmental taxonomy of glioblastoma defined and maintained by MicroRNAs. Cancer Res. 2011;71:3387–99. doi: 10.1158/0008-5472.CAN-10-4117. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 64.Srinivasan S, Patric IR, Somasundaram K. A ten-microRNA expression signature predicts survival in glioblastoma. PLoS One. 2011;6:e17438. doi: 10.1371/journal.pone.0017438. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 65.Lages E, Guttin A, El Atifi M, Ramus C, Ipas H, Dupré I, et al. MicroRNA and target protein patterns reveal physiopathological features of glioma subtypes. PLoS One. 2011;6:e20600. doi: 10.1371/journal.pone.0020600. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 66.Hayes J, Thygesen H, Tumilson C, Droop A, Boissinot M, Hughes TA, et al. Prediction of clinical outcome in glioblastoma using a biologically relevant nine-microRNA signature. Mol Oncol. 2015;9:704–14. doi: 10.1016/j.molonc.2014.11.004. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 67.Guan Y, Mizoguchi M, Yoshimoto K, Hata N, Shono T, Suzuki SO, et al. MiRNA-196 is upregulated in glioblastoma but not in anaplastic astrocytoma and has prognostic significance. Clin Cancer Res. 2010;16:4289–97. doi: 10.1158/1078-0432.CCR-10-0207. [DOI] [PubMed] [Google Scholar]
- 68.Yan W, Li R, Liu Y, Yang P, Wang Z, Zhang C, et al. MicroRNA expression patterns in the malignant progression of gliomas and a 5-microRNA signature for prognosis. Oncotarget. 2014;5:12908–15. doi: 10.18632/oncotarget.2679. [DOI] [PMC free article] [PubMed] [Google Scholar]