
Figure 1. Schematic illustration of metabolic pathways of neuron-astrocyte signaling relevant to epileptic seizures.
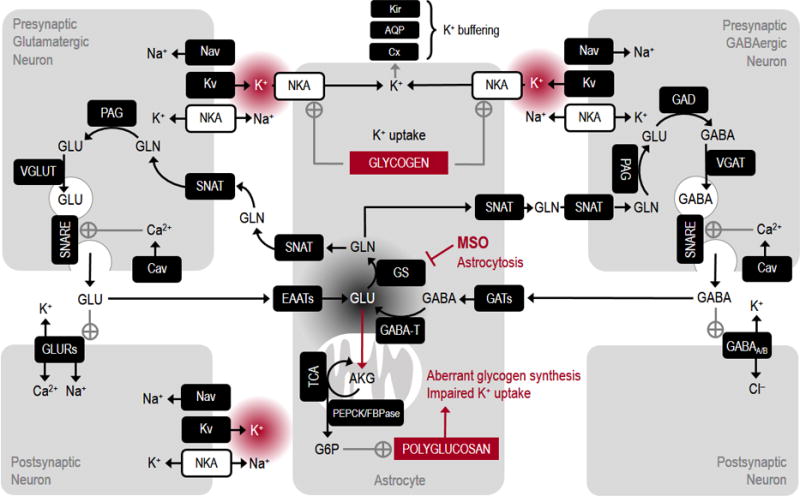
Neuronal electrical activity includes generation and propagation of axonal action potentials through opening of depolarizing Nav and hyperpolarizing Kv. Arrival of traveling depolarization at axon terminal triggers opening of Cav and entry of Ca2+, which in turn activates the secretory machinery of vesicle exocytosis. Relased transmitter (either glutamate or GABA in glutamatergic or GABAergic neurons, respectively) binds to postsynaptic receptors (GLURs or GABAA/B) thereby producing depolarizing excitatory or hyperpolarizing inhibitory ion fluxes on postsynaptic neurons. Depolarization is eventually amplified and propagated in dendrites by Nav and Kv. Transmitter molecules are readily taken up mainly by astrocytes via the relative transporter proteins (EAATs or GATs) and then returned back to neurons in the glutamate-glutamine cycle (note that GABA is converted to glutamate by astrocytic GABA-T). Cycling of neurotransmitter strictly requires the action of astrocyte-specific GS. Inhibition of GS by MSO and/or loss-of-function during epilepsy-induced astrocytosis determines the depletion of presynaptic transmitter molecules and accumulation of glutamate in astrocytes. High glutamate levels in these cells result in excessive channeling of glutamate in TCA cycle and dowstreaming stimulation of gluconeogenic pathway (e.g. PEPCK and FBPase). The activation of FBPase is correlated with new synthesis of aberrant glycogen molecules called polyglucosans, which are partly insoluble and thus inaccessible to degrading enzymes. Ion gradients are primarily re-established by action of NKA in both neurons and astrocytes. However, astrocytic NKA is poised for reuptake of neuronally released K+, a process requiring glycogenolysis. Thus, defective glycogenolysis due to accumulation of polyglucosans impairs astrocytic K+ uptake. Excess extracellular K+ shifts neuronal membrane potential towards chronically depolarized values and possibly initiates and perpetuates the epileptic seizure. Note that in the figure, the stimulation of polyglucosans synthesis by glucose-6-phosphate (G6P) is present only for illustration purposes, as the exact mechanisms leading to polyglucosans synthesis are presently unknown. AQP, aquaporin; Cav, voltage-gated Ca2+ channel; Cx, connexin; EAATs, excitatory amino acid transporters; FBPase, fructose-1,6-bisphosphatase; GAD, glutamate decarboxylase; GABAA/B, GABA receptors; GABA-T, GABA transaminase; GATs, GABA transporters; GLURs, glutamate receptors; GS, glutamine synthetase; Kir, inward rectifier K+ channel; Kv, voltage-gated K+ channel; MSO, L-methionine-SR-sulfoximine; Nav, voltage-gated Na+ channel; NKA, Na+-K+-activated adenosintrisphosphatase; PAG, phosphate-activated glutaminase; PEPCK, phosphoenolpyruvate carboxykinase; SNARE, soluble N-ethylmaleimide-sensitive factor attachment protein receptor; SNAT, sodium-coupled neutral amino acid transporter; TCA, tricarboxylic acid cycle; VGAT, vesicular GABA transporter; VGLUT, vesicular glutamate transporter.