
Fig. 3.
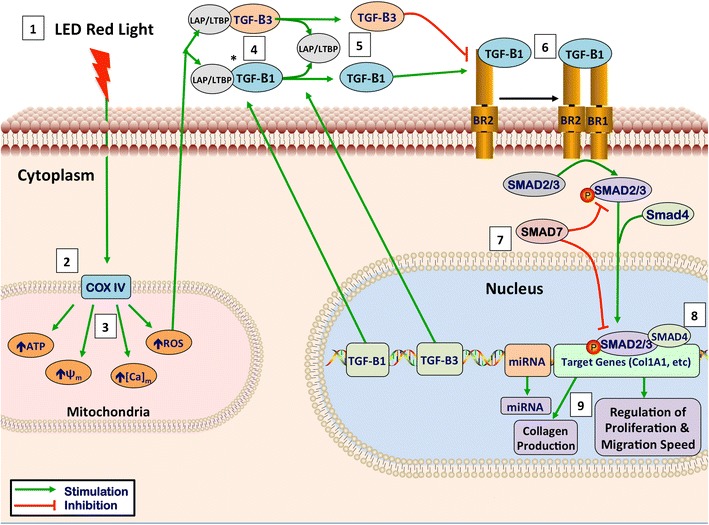
Theoretical mechanism of LED red light photobiomodulation. 1 Light has optimal tissue penetration when its wavelength is within the “optical window,” (600–1070 nm). Red light (620–750 nm) takes advantage of this penetration window [15]. 2 LED-RL stimulates the photo-acceptor copper complex in cytochrome C oxidase, stimulating the photodissociation of nitric oxide (NO), leading to upregulation of the electron transport chain [15, 16]. 3 This stimulation of the electron transport leads to the following intramitochondrial changes: increased generation of ATP and reactive oxygen species (ROS), increased intramitochondrial calcium concentration, and an increase in the mitochondrial membrane potential [16]. 4 TGF-Betas are secreted associated with latency-associated peptide (LAP). These associated latency peptides determine the activity TGF-Beta subtypes [17]. Reactive oxygen species have been shown to trigger a conformational change in LAP, thus freeing TGF-Beta 1 from its latency peptide [17–20]. It has been suggested that activation of TGF-Beta 3 may function by a similar ROS-induced release of LAP [20]. 5 TGF-B1, upon being released from its latency complex, binds to the TGF-Beta receptor II (TGF-BRII) stimulating activation [21]. TGF-B3 inhibits activation of the TGF-BRII and antagonizes TGF-B1 signaling [21]. 6 TGF-BRII then forms a heteromeric complex with TGF-BRI, causing the phosphorylation of specific serine residues [21, 22]. 7 When activated, the intracytoplasmic domain of the TGF-Beta receptor complex phosphorylates SMAD proteins. Once phosphorylated, pSMAD2/3 has the ability to migrate to the nucleus and associates with DNA-binding partners to cause changes in target gene expression [22]. SMAD7 activation functions as a negative feedback loop, inhibiting TGF-B1 signaling [23]. 8 SMAD signaling alters COL1A1 gene expression leading to changes in extracellular collagen deposition and changes in fibroblast proliferation [24]. In addition, SMAD signaling contributes to the collagen deposition and pathogenesis of skin fibrosis [25]. These pathways, and related pathways such as Akt, are believed to contribute to skin fibrosis through modulation of fibroblast proliferation and migration speed [26]. 9 TGF-B1 has been shown to increase cell proliferation at low levels; however, high levels of TGF-B1 inhibits dermal fibroblast proliferation, supporting the idea that modulation of levels of TGF-B1 may contribute to LED-RL’s modulation of proliferation [27]. In addition, the release of TGF-B1 or TGF-B3 from their LAP has been suggested as a possible mechanism behind the photobiomodulation of LED-RL [20]. Figure legend: ψm, mitochondrial membrane potential; [Ca]m, intramitochondrial calcium concentration; ATP, adenosine triphosphate; ROS, reactive oxygen species; LAP/LTBP, latency-associate peptide/latent transforming growth factor beta binding protein; TGF-B1, transforming growth factor beta-1; TGF-B3, transforming growth factor beta-3; BR2, transforming growth factor receptor II; BR1, transforming growth factor receptor I; miRNA, microRNA