
Fig. 2.
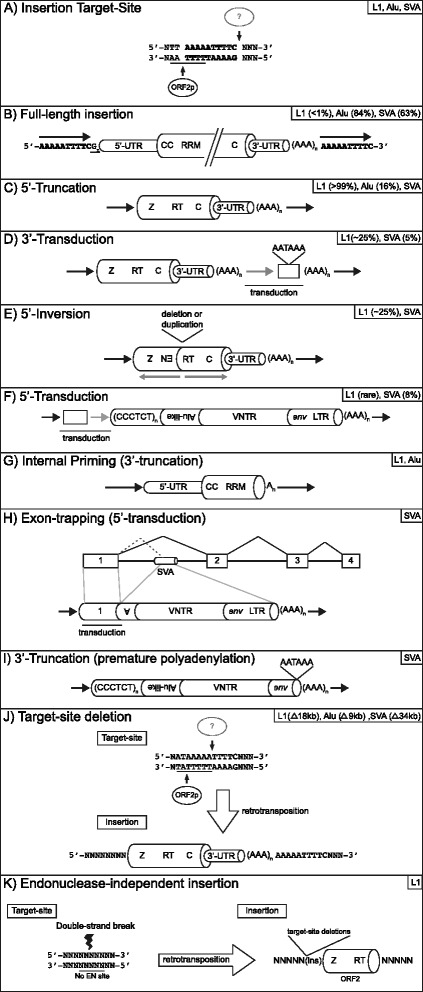
Anatomy of retrotransposon insertions. A variety of structures for retrotransposon insertions (a-k) identified by genomic studies, cell culture retrotransposition assays, and disease-causing insertions that have been reported is shown. Reported frequencies, either from genomic analysis or cell-culture retrotransposition assays, for each structure is located in the upper right hand corner of each panel. If no frequency data has been reported only the element’s name is shown. These structures have provided key insights into the mechanism of target-primed reverse transcription [77], retrotransposon transcript structure [127], and the mechanism by which LINE-1-mediated retrotransposition events contribute to genome evolution [62]. A) LINE-1 target-site. Most insertions occur at asymmetric AT-rich sequences [6, 37, 51, 86]. The first step of TPRT is cleavage of the bottom-strand by ORF2p endonuclease activity at a motif resembling 5′-TTTT/AA-3′ [245]. The nuclease responsible for top-strand cleavage is currently unknown. The nature of the staggered cleavage events generates a target-site duplication (TSD, sequence in bold). a TSD (black horizontal arrows) is used to define the boundaries of an insertion and considered a hallmark of LINE-1-mediated retrotransposition events. b Full-length insertion. It is generally accepted that in order for an element to be retrotransposition-competent it must be full-length. c 5′-truncated insertions. Most LINE-1 s in the human genome are grossly truncated at their 5′-end [6, 36, 37]. In contrast, most Alus [243] and SVA elements are full-length [123, 127]. To date, no consensus sequence has been identified in LINE-1 or SVA insertions regarding the mechanism of 5′-truncation. However, a new report implicates stem-loop structures as a factor driving 5′-truncation in recent Alu insertions [114]. d 3′-transduction. Although the first report of a 3′-transduction was an LINE-1 insertion into the dystrophin gene resulting in Duchenne’s muscular dystrophy in 1994 [176], it would be several years before the significance of this chimeric insertion was uncovered. Several years later, as one of the first insights gained from insertions recovered from cell-culture retrotransposition assays, it was reported that LINE-1 frequently bypassed its own polyA signal (AATAAA) in favor of a downstream one (AATAAA) [42]. Subsequently, elegant experimental analysis revealed that utilizing a downstream polyA signal could result in LINE-1-mediated exon-shuffling [62]. An insertion containing a 3′-transduction will typically contain two homopolymer stretches (AAAn) and contain the 3′-TSD from the source locus (gray horizontal arrow) as part of the transduced sequence. Notably, insertions containing serial 3′-transductions have been reported and can be used to track the evolutionary history of an element [246]. e 5′-end inversions. Another hallmark of LINE-1-mediated retrotransposition events is the inversion of the 5′-end (gray horizontal arrow) of the retrotransposon sequence [35]. Small indels are typically identified at the inversion breakpoint [88]. Inversions have only been reported for LINE-1 s, SVAs, and processed pseudogenes [196]. 5′-end inversion is presumed not to occur for Alus due to their short length. It has been hypothesized that a phenomenon referred to as twin-priming may account for the frequent inversions associated with LINE-1-mediated retrotransposition events [88]. f 5′-transduction. In some instances, LINE-1 [82] or SVA transcription [126, 127] may initiate upstream of the internal promoter generating a chimeric transcript. Retrotransposition of this sequence results in duplication of the sequence 5′- of the source locus at a new genomic location. It has been speculated that 5′-transductions are relatively common for SVA elements due to their weaker internal promoter compared to LINE-1, which has a very strong internal promoter, where only a handful of 5′-transductions have been reported [82]. g Internal priming. Occasionally following bottom-strand cleavage, internal A-rich sequences upstream in the retrotransposon RNA may basepair with the T-rich overhang at the target-site instead of the 3′-polyA tail, followed by first-strand cDNA synthesis by ORF2p [247–249]. These insertions can be deemed a type of 3′-truncation. h Exon-trapping. Retrotransposons are dispersed throughout the genome including intronic sequence. LINE-1, Alu, SVA all have been reported to contain numerous splice sites and be incorporated into the transcriptome [105, 127, 128, 134, 250]. Interestingly, LINE-1 internal splicing can generate a transcript lacking ORF1 but maintaining a functional ORF2 [251]. In some instances, at least for SVA, retrotransposition of chimeric transcripts containing upstream exons may occur [127, 128, 132]. Notably, SVA itself is thought to have originated from alternative splicing from genomic repeats [126] and SVA-related elements (e.g. LAVA, PVA) appear to have acquired distinct 3′-domains via splicing in gibbons [125, 135, 136, 138]. I) 3′-truncation. Premature polyadenylation using either canonical or non-canonical polyadenylation sites results in LINE-1 or SVA RNAs lacking 3′-sequence [127, 252]. If this RNA is retrotransposed, it will result in a 3′-truncated insertion. Consistent with the dispensability of SVA domains [130], 3′-truncations may be more frequent for SVA compared to LINE-1. In principle, 3′-truncated LINE-1 RNAs containing ORF1 coding sequence might be actively retrotransposed as in the case of ORF1 mNEOi in cell culture [144] and the presence of half-LINE-1 (HAL1) insertions in mammalian genomes [253]. j Target-site deletion. Another surprise from cell culture retrotransposition assays was the discovery of large deletions associated with new retrotransposition events [82, 83]. Genomic deletions up to 1 MB have been associated with LINE-1 mediated retrotransposition events in vivo [153]. These insertions occur at a LINE-1 EN cleavage site, are generated by ORF2 reverse-transcriptase activity, and end in a 3-polyA tail. Currently, the mechanism driving 5-targe-site deletions is unclear; yet, it is tempting to speculate that chromatin looping along with cleavage by LINE-1 or another nuclease may play important roles [82, 83]. k Endonuclease-independent (ENi) insertion. Eni insertions were discovered by the Moran lab when carrying out retrotransposition assays in different Chinese Hamster Ovary (CHO) cell lines lacking key DNA repair factors [213]. Frequent retrotransposition was observed for an engineered LINE-1 element construct, with a catalytically inactive EN, in these cells but not HeLa cells. Characterization of recovered insertions revealed LINE-1 integration at genomic sites not resembling the LINE-1 EN consensus cleavage site. In addition, the insertions were typically truncated at both the 5′-and 3′-ends [213]. These data suggest that LINE-1 can serve as a “molecular band-aid” [254] at double-stranded DNA breaks [213–215] and that LINE-1 s lacking a functional EN domain may be able to retrotranspose in certain contexts. Building on these studies it was later reported that LINE-1 s can also integrate at dysfunctional telomeres in an endonuclease-independent manner [216]