

Abstract
Staurozoan classification is highly subjective, based on phylogeny-free inferences, and suborders, families, and genera are commonly defined by homoplasies. Additionally, many characters used in the taxonomy of the group have ontogenetic and intraspecific variation, and demand new and consistent assessments to establish their correct homologies. Consequently, Staurozoa is in need of a thorough systematic revision. The aim of this study is to propose a comprehensive phylogenetic hypothesis for Staurozoa, providing the first phylogenetic classification for the group. According to our working hypothesis based on a combined set of molecular data (mitochondrial markers COI and 16S, and nuclear markers ITS, 18S, and 28S), the traditional suborders Cleistocarpida (animals with claustrum) and Eleutherocarpida (animals without claustrum) are not monophyletic. Instead, our results show that staurozoans are divided into two groups, herein named Amyostaurida and Myostaurida, which can be distinguished by the absence/presence of interradial longitudinal muscles in the peduncle, respectively. We propose a taxonomic revision at the family and genus levels that preserves the monophyly of taxa. We provide a key for staurozoan genera and discuss the evolution of the main characters used in staurozoan taxonomy.
Keywords: Evolution, Taxonomy, Phylogeny, Medusozoa, Stauromedusae
Introduction
Staurozoa is a class of benthic cnidarians, the so-called stalked jellyfishes (Figs. 1 and 2), represented by approximately 50 species (Clark, 1863; Kramp, 1961; Daly et al., 2007). However, from the first stauromedusan species described (Lucernaria quadricornis Müller, 1776) until their proposition as the fifth class of Cnidaria (Marques & Collins, 2004), the group has had a long history of classifications, being labeled as a “puzzling group” (Gwilliam, 1956). While one species was erroneously first placed among sea cucumbers (Manania auricula as Holothuria lagenam referens Müller, 1776), most assessments prior to the 1850’s assumed that they were closely related to sea anemones (Cuvier, 1817; Cuvier, 1830) until Sars (1846) noted that the presence of gastric cirri suggested that they were allied with the jellyfishes. Reflecting this thinking, Goette (1887) included Stauromedusae as a suborder within Scyphozoa, a position that was only recently challenged. Marques & Collins (2004) proposed the class based on a phylogenetic analysis of morphological and life cycle traits, as the clade uniting the fossil group Conulatae and the Stauromedusae. In light of further evidence from the fossil record, a subsequent analysis of a similar dataset contradicted the hypothesis that Conulatae and Stauromedusae form a clade, and proposed the composition of Staurozoa to consist exclusively of the extant Stauromedusae (Van Iten et al., 2006). The same analysis suggested that Staurozoa is the sister group to all other medusozoans (Cubozoa, Hydrozoa, and Scyphozoa), a result corroborated by analyses of nuclear ribosomal data (Collins et al., 2006; see also Van Iten et al., 2014). In contrast, however, analyses of complete mitochondrial genome data (Kayal et al., 2013) suggest that Staurozoa may be the sister group of Cubozoa, and more recent phylogenomic analyses support a clade formed by Staurozoa, Cubozoa, and Scyphozoa (Zapata et al., 2015), demonstrating that more studies are necessary to reach a stable topology for Cnidaria.
Figure 1. Diversity of stalked jellyfishes.

Calvadosia cruxmelitensis: (A) lateral view, (B) oral view (photo credit: David Fenwick); Calvadosia campanulata: (C) lateral view, (D) oral view (photo credit: David Fenwick); Calvadosia nagatensis: (E) oral view (photo credit: Yayoi Hirano); Craterolophus convolvulus: (F, G) lateral view (photo credit: David Fenwick); Depastromorpha africana: (H) lateral view (photo credit: Yayoi Hirano); Haliclystus tenuis: (I) lateral view (photo credit: Yayoi Hirano); Haliclystus borealis: (J) lateral view (photo credit: Yayoi Hirano); Haliclystus octoradiatus: (K) oral view (photo credit: David Fenwick); Haliclystus inabai: (L) lateral view (photo credit: Yayoi Hirano); Kyopoda lamberti: (M) lateral view (photo credit: courtesy of Ronald Shimek); Lipkea sp. Japan: (N) oral view (photo credit: Yayoi Hirano); Stylocoronella riedli: (O) lateral view (proto credit: courtesy of Mat Vestjens and Anne Frijsinger); Lucernaria janetae: (P) lateral and oral views (photo credit: courtesy of Richard Lutz); Manania uchidai: (Q) lateral view (photo credit: Yayoi Hirano); Manania gwilliami: (R) oral view (photo credit: courtesy of Ronald Shimek); Manania handi: (S) lateral view (photo credit: Claudia Mills).
Figure 2. General external anatomy of stalked jellyfishes.

Craterolophus convolvulus: (A) lateral view, (B) oral view. Abbreviations: am, arm; cl, calyx; gd, gonad; mn, manubrium; pd, pedal disk; pe, peduncle; tc, tentacle cluster. Photo credit: David Fenwick.
Although evolutionary studies have supported monophyly of the class (Collins & Daly, 2005; Collins et al., 2006; Kayal et al., 2013), comparatively little effort has been applied toward determining the systematic relationships among species of Staurozoa, with rare exceptions (Collins & Daly, 2005; Lutz et al., 2006). The current classification of Staurozoa is mainly based on the proposals of Clark (1863), Haeckel (1879), Uchida (1929) and Carlgren (1935), and is completely focused on anatomical features. Uchida (1973) proposed a hypothesis of relationship among families of stalked jellyfishes based on the characters that he regarded as important, but this analysis was not derived from specific evolutionary methods. A recent molecular inference, with limited taxon sampling, demonstrated the need for reassessing suprageneric clades, because several were found not to be monophyletic (Collins & Daly, 2005). Additionally, many characters used in the taxonomy of the group have ontogenetic and intraspecific variation, and demand consistent assessments and clarifications to establish their correct homologies (Miranda, Morandini & Marques, 2009). Consequently, staurozoan classification and taxonomy is subjective, based on phylogeny-free inferences, and families and genera may be commonly defined by homoplasies (Collins & Daly, 2005). Therefore, Staurozoa is in need of a thorough systematic revision.
Inferences about the relationships among staurozoan species are especially important because of the phylogenetic status and position of Staurozoa, as a distinct clade separate from the other major cnidarian groups (Anthozoa, Cubozoa, Hydrozoa, and Scyphozoa) (Collins et al., 2006; Van Iten et al., 2006; Kayal et al., 2013; Zapata et al., 2015). The peculiar life cycle of staurozoans (Wietrzykowski, 1912; Kikinger & Salvini-Plawen, 1995; Miranda, Collins & Marques, 2010) is tightly connected to their unique anatomy, in which characters of polypoid and medusoid stages are present in the same stauromedusa (Miranda, Collins & Marques, 2013). Our expectation is that a better understanding and interpretation of the character evolution within the group will provide crucial information for inferences in cnidarian evolution.
Therefore, it is of the utmost importance to carry out an evolutionary analysis encompassing a large number of species of Staurozoa. This study presents the most comprehensive phylogenetic hypothesis for Staurozoa yet proposed and provides the first phylogenetic classification for the group. Further, we provide a key for staurozoan genera and discuss evolution of the main characters used in staurozoan taxonomy.
Material and Methods
Molecular
Twenty-four species from ten genera, plus eight non-identified species (identified to genus level), from different regions of the world, were used in the molecular analyses (Table 1). Tissue samples from the tentacle clusters (or marginal lobes for Lipkea spp.) were removed and preserved in 90–100% ethanol, and stored at −20 °C. DNA extractions were carried out with InstaGene (Bio-Rad) at the Universidade de São Paulo, Instituto de Biociências (IB-USP, Brazil), or using an organic phenol-chloroform method on the automated DNA isolation system, AutoGenPrep 965 (AutoGen Inc., Holliston, MA, USA) at the Smithsonian’s Laboratories of Analytical Biology (LAB, USA), following the manufacturers’ protocols. Genes were amplified using PCR, then purified with AMPure® (Agencourt®) or ExoSAP. Different molecular markers (mitochondrial COI and 16S; nuclear ITS–ITS1+5.8S+ITS2, 18S, and 28S) were targeted for analyses (Tables 2 and 3). These markers were previously adopted and have been shown to be efficient for evolutionary studies in medusozoans (Dawson, 2004; Collins et al., 2006; Collins et al., 2008; Miranda, Collins & Marques, 2010; Nawrocki et al., 2013; Cunha, Genzano & Marques, 2015). DNA sequencing was done using the BigDye® Terminator v3.1 kit (Applied Biosystems) and the same primers used for PCR (Table 2). The procedure was carried out on an ABI PRISM® 3100 Genetic Analyzer (Hitachi). Samples were extracted, amplified and sequenced at LAB (USA) and IB-USP (Brazil). Out-group sequences (Anthozoa, Cubozoa, Hydrozoa, and Scyphozoa) were obtained in GenBank (Table 4).
Table 1. Species used in the phylogenetic analyses (parsimony, maximum likelihood, and Bayesian inference).
Based on mitochondrial molecular markers (COI and 16S) and nuclear molecular markers (ITS, 18S, and 28S).
| Species | Specimens | GenBank accession numbers | Locality | Voucher | ||||
|---|---|---|---|---|---|---|---|---|
| COI | 16S | ITS | 18S | 28S | ||||
| Craterolophus convolvulus | Craterolophus convolvulus GER | KU257472 | AY845343 | KU308618 | AY845344 | AY920781 | Helgoland, North Sea, Germany | USNM 1073330, 1073339 |
| Craterolophus convolvulus UK | KU257473 | KU257497 | KU308619 | – | – | Sennen Cove, Cornwall, England | USNM 1286315 | |
| Craterolophus convolvulus USA | – | KU257498 | KU308620 | KU308557 | KU308586 | Rye Beach, Rye, New Hampshire, USA | MZUSP 002730 | |
| Depastromorpha africana | Depastromorpha africana 1* | – | – | – | KU308558 | – | Kalk Bay, Cape Town, South Africa | MZUSP 002733 |
| Depastromorpha africana 2* | KU257474 | KU257499 | KU308621 | – | KU308587 | Kalk Bay, Cape Town, South Africa | MZUSP 002734 | |
| Haliclystus antarcticus | Haliclystus antarcticus ANT | KU257475 | EU294003 | FJ874779 | EU247811 | KU308588 | Argentine antarctic station Jubany, King George Island, Antarctica | None |
| Haliclystus antarcticus Chile | – | AY845340 | KU308622 | AY845348 | KU308589 | Valdivia, Chile | None | |
| Haliclystus borealis | Haliclystus borealis | – | KU257500 | KU308623 | KU308559 | KU308590 | Muroran, Hokkaido, Japan | USNM 1106650 |
| Haliclystus californiensis | Haliclystus californiensis | GU201831 | GU201829 | KU308624 | KU308560 | KU308591 | Otter Point, Pacific Grove, California, USA | USNM 1106657 |
| Haliclystus octoradiatus | Haliclystus octoradiatus | KU257476 | KU257501 | KU308625 | KU308561 | KU308592 | Cornwall, England | USNM 1286385 |
| Haliclystus “sanjuanensis” | Haliclystus “sanjuanensis” | KU257477 | HM022151 | HM022145 | KU308562 | KU308593 | San Juan Island, Washington, USA | USNM 1106935 |
| Haliclystus stejnegeri | Haliclystus stejnegeri | KU257478 | HM022153 | HM022147 | KU308563 | KU308594 | Daikokujima, Akkeshi, Hokkaido, Japan | KUNHM 002673-B |
| Haliclystus tenuis | Haliclystus tenuis | KU257479 | HM022154 | HM022148 | KU308564 | KU308595 | Muroran, Hokkaido, Japan | USNM 1106651 |
| Kishinouyea corbini | Kishinouyea corbini | – | KU257502 | – | KU308565 | KU308596 | Aracruz, Espirito Santo, Brazil | MZUSP 1563 |
| Kishinouyea nagatensis | Kishinouyea nagatensis | – | – | – | – | KU308597 | Shimoda, Shizuoka, Japan | USNM 1106985 |
| Lipkea ruspoliana | Lipkea ruspoliana 1* | – | KU257503 | KU308626 | KU308566 | KU308598 | Aquarium, Oceanographic Museum of Monaco | USNM 1315313 |
| Lipkea ruspoliana 2* | KU257480 | – | – | – | – | Aquarium, Oceanographic Museum of Monaco | USNM 1315317 | |
| Lucernaria bathyphila | Lucernaria bathyphila Arctic | – | KU257504 | – | KU308567 | – | Arctic, Mid-Ocean Ridge | USNM 1301002 |
| Lucernaria bathyphila Deep Sea | KU257481 | KU257505 | KU308627 | – | KU308599 | Faroe-Shetland Channel between Faroe and Shetland Islands | USNM 1300113 | |
| Lucernaria janetae | Lucernaria janetae 1* | – | AY845342 | FJ874778 | AY845345 | KU308600 | East Pacific Rise | FMNH 10329 |
| Lucernaria janetae 2* | JN700946 | – | – | – | – | East Pacific Rise | None | |
| Lucernaria quadricornis | Lucernaria quadricornis 1* | – | – | – | – | KU308601 | Keret Inlet, off Sredny Island, Black Rock, White Sea, Russia | USNM 1106636 |
| Lucernaria quadricornis 2* | – | KU257506 | KU308628 | KU308568 | – | Near White Sea Biological Station of Moscow State University, White Sea, Russia | USNM 1102441 | |
| Lucernaria sainthilairei | Lucernaria sainthilairei 1* | – | – | – | KU308569 | – | Near White Sea Biological Station of Moscow State University, White Sea, Russia | USNM 1106245 |
| Lucernaria sainthilairei 2* | KU257482 | KU257507 | KU308629 | – | KU308602 | Nicolskaya Inlet, off Bolshoy Medvedok Island, Kandalaksha Bay, White Sea, Russia | USNM 1106643 | |
| Lucernariopsis campanulata | Lucernariopsis campanulata | KU257483 | KU257508 | KU308630 | KU308570 | KU308603 | Battery Rocks, Penzance, Cornwall, England | USNM 1286317 |
| Lucernariopsis cruxmelitensis | Lucernariopsis cruxmelitensis | KU257484 | KU257509 | KU308631 | KU308571 | KU308604 | Penzance, Cornwall, England | USNM 1286321 |
| Lucernariopsis tasmaniensis | Lucernariopsis tasmaniensis | – | KU257510 | KU308632 | KU308572 | – | Kitty Miller Bay, Phillip Island, Victoria, Australia | MV F158628 |
| Lucernariopsis vanhoeffeni | Lucernariopsis vanhoeffeni | KU257485 | KU257511 | KU308633 | KU308573 | KU308605 | Trinity Island, Palmer Archipelago, Antarctica | USNM 1106658 |
| Manania gwilliami | Manania gwilliami 1* | – | KU257512 | KU308634 | KU308574 | KU308606 | Monterey Harbor, California, USA | USNM 1106649 |
| Manania gwilliami 2* | KU257486 | – | – | – | – | Pacific Grove Marine Gardens Park, California, USA | USNM 1106662 | |
| Manania uchidai | Manania uchidai | – | KU257513 | KU308635 | KU308575 | KU308607 | Daikokujima, Akkeshi, Hokkaido, Japan | KUNHM 002673-A |
| Sasakiella cruciformis | Sasakiella cruciformis | KU257487 | KU257514 | – | KU308576 | KU308608 | Oshoro, Hokkaido, Japan | USNM 1106984 |
| Stenoscyphus inabai | Stenoscyphus inabai | KU257488 | KU257515 | KU308636 | KU308577 | KU308609 | Oshoro, Hokkaido, Japan | Photo voucher only |
| Non identified species | ||||||||
| Depastromorpha sp. AUS | Depastromorpha sp. AUS | KU257489 | KU257516 | KU308637 | KU308578 | KU308610 | Outside Port Fairy, Abalone Farm, Victoria, Australia | MV F197278 |
| Kishinouyea sp. Moorea | Kishinouyea sp. Moorea | KU257490 | KU257517 | KU308638 | KU308579 | KU308611 | Moorea, French Polynesia | UF:Cnidaria:7226 |
| Kishinouyea sp. NZ | Kishinouyea sp. NZ | KU257491 | KU257518 | KU308639 | KU308580 | KU308612 | Taputeranga Marine Reserve, Wellington, New Zealand | NIWA 86808 |
| Kishinouyea sp. SAF | Kishinouyea sp. SAF 1* | – | – | – | – | KU308613 | Cape Town, South Africa | MZUSP 002731 |
| Kishinouyea sp. SAF 2* | KU257492 | KU257519 | KU308640 | KU308581 | – | Cape Town, South Africa | MZUSP 002732 | |
| Lipkea sp. JAP | Lipkea sp. JAP 1* | ** | Aquarium, Katsuura, Chiba, Japan | USNM 1315325 | ||||
| Lipkea sp. JAP 2* | KU257493 | KU257520 | – | KU308582 | KU308614 | Aquarium, Katsuura, Chiba, Japan | USNM 1315326 | |
| Lucernaria sp. EPR | Lucernaria sp. EPR | KU257494 | DQ465037 | KU308641 | KU308583 | KU308615 | East Pacific Rise, 20 degrees south | USNM 1086349 |
| Lucernariopsis sp. NZ | Lucernariopsis sp. NZ | KU257495 | KU257521 | KU308642 | KU308584 | KU308616 | Taputeranga Marine Reserve, Wellington, New Zealand | NIWA 86809 |
| Stenoscyphus sp. AUS | Stenoscyphus sp. AUS | KU257496 | KU257522 | KU308643 | KU308585 | KU308617 | Williamstown, The Jawbone, Victoria, Australia | MV F190063 |
Notes:
More than one specimen for each species was used to generate the combined alignment. ANT, Antarctica; AUS, Australia; EPR, East Pacific Rise; GER, Germany; JAP, Japan; NZ, New Zealand; SAF, South Africa; UK, the United Kingdom; USA, the United States of America; FMNH, Field Museum of Natural History, USA; KUNHM, University of Kansas Natural History Museum; MV, Museum Victoria, Australia; MZUSP, Museum of Zoology of the University of São Paulo; NIWA, National Institute of Water and Atmospheric Research; UF, University of Florida Museum of Natural History; USNM, National Museum of Natural History, Smithsonian, USA.
Sequence with less than 200 nucleotides: ACGCCGTGCCAGGCCAAAATGTATTTTGTTACCTGCTGCACGGTGATGCTGAGCGCATTTTCTTTCTTCGTGGGCAAGAAAGAGGTGGTCGATAGATACGAGAGAGAGTGAGAGAGACGCGCGTCGTCCATCTCTCACTGACAATGACCTCAG.
Table 2. Primer sequences for polymerase chain reaction (PCR) and sequencing reaction.
| Genes | Primers | Sequences | References |
|---|---|---|---|
| COI | jgHCO2198 | TITCIACIAAYCAYAARGAYATTGG | Geller et al. (2013) |
| jgLCO1490 | TAIACYTCIGGRTGICCRAARAAYCA | Geller et al. (2013) | |
| 16S | F1mod | TCGACTGTTTACCAAAAACATA | Cunningham & Buss (1993) and Cartwright et al. (2008) |
| R2 | ACGGAATGAACTCAAATCATGTAAG | Cunningham & Buss (1993) and Cartwright et al. (2008) | |
| rnl_f_jl | GACTGTTTACCAAAGACATAGC | Designed by J. Lawley | |
| rnl_r_jl | AAGATAGAAACCTTCCTGTC | Designed by J. Lawley | |
| ITS | jfITS1–5f | GGTTTCCGTAGGTGAACCTGCGGAAGGATC | Dawson & Jacobs (2001) |
| CAS28SB1d | TTCTTTTCCTCCSCTTAYTRATATGCTTAA | Ji, Zhang & He (2003) | |
| C2 | GAAAAGAACTTTGRARAGAGAGT | Chombard et al. (1997) | |
| D2 | TCCGTGTTTCAAGACGGG | Chombard et al. (1997) | |
| 18S (SSU) | AF_cnidarian | GTGGYAATTCTAGAGCTAATACATGCG | Designed by R. Wilson |
| BR_cnidarian | GCGACGGGCGGTGTGTAC | Designed by R. Wilson | |
| IF_cnidarian | GGGGGCATYCGTATTTCGTTG | Designed by R. Wilson | |
| IR_cnidarian | CAACGAAATACGRATGCCCCC | Designed by R. Wilson | |
| C_new cnidarian | CAGCCGCGGTAATTCCAGC | Designed by R. Wilson | |
| L_new cnidarian | CCTRTTCCATTATTCCATGCTC | Designed by R. Wilson | |
| O_new cnidarian | GGTCCAGACATAGTAAGGATTG | Designed by R. Wilson | |
| 1800R18S | GTTCACCTACYGAAACCTTGTT | Redmond et al. (2007) | |
| 28S (LSU) | F63 mod | ACCCGCTGAAYTTAAGCATATHANTMAG | Medina et al. (2001) |
| F63sq | AATAAGCGGAGGAAAAGAAAC | Medina et al. (2001) | |
| F97 | CCYYAGTAACGGCGAGT | Evans et al. (2008) | |
| F635 | CCGTCTTGAAACACGGACC | Medina et al. (2001) | |
| F1379sq | GACAGCAGGACGGTGGYCATGG | Medina et al. (2001) | |
| F1383 | GGACGGTGGCCATGGAAGT | Collins et al. (2008) and Evans et al. (2008) | |
| F1586 | GTGCAGATCTTGGTDGNAGTAGCAAATATTC | Medina et al. (2001) | |
| F1689 | CTAAGMSRYAGGGAAAYTC | Collins et al. (2008) | |
| F2076sq | TAACYTCGGGAWAAGGATTGGCTC | Medina et al. (2001) | |
| F2766sq | AGTTTGGCTGGGGCGGYACA | Medina et al. (2001) | |
| F2800 | GCAGGTGTCCTAAGGYRAGCTC | Voigt et al. (2004) | |
| R635sq | GGTCCGTGTTTCAAGACGG | Medina et al. (2001) | |
| R1411sq | GTTGTTACACACTCCTTAGCGG | Medina et al. (2001) | |
| R1630 | CCYTTCYCCWCTCRGYCTTC | Medina et al. (2001) | |
| R2077sq | GAGCCAATCCTTWTCCCGARGTT | Medina et al. (2001) | |
| R2084 | AGAGCCAATCCTTTTCC | Evans et al. (2008) and Collins et al. (2008) | |
| R2766sq | CAGRTGTRCCGCCCCAGCCAAACT | Medina et al. (2001) | |
| R2800 | GAGCTYRCCTTAGGACACCTGC | Voigt et al. (2004) | |
| R3238 | SWACAGATGGTAGCTTCG | Evans et al. (2008) and Collins et al. (2008) | |
| R3264 | TTCYGACTTAGAGGCGTTCAG | Medina et al. (2001) |
Table 3. Polymerase chain reaction (PCR) conditions for the different molecular markers used in the phylogenetic analyses.
| Molecular marker | PCR condition |
|---|---|
| COI | 94 °C: 5 min |
| 35 × −94 °C: 30 s; 50 °C: 40 s; 72 °C: 1 min | |
| 72 °C: 7 min | |
| 4 °C: forever | |
| 16S | 95 °C: 5 min |
| 35 × −95 °C: 30 s; 45 °C: 50 s; 72 °C: 1 min | |
| 72 °C: 5 min | |
| 4 °C: forever | |
| ITS | 94 °C: 5 min |
| 35 × −94 °C: 30 s; 60 °C: 40 s; 72 °C: 1 min | |
| 72 °C: 10 min | |
| 4 °C: forever | |
| 18S (SSU) | 94 °C: 5 min |
| 35 × −94 °C: 30 s; 57 °C: 30 s; 72 °C: 1 min | |
| 72 °C: 7 min | |
| 4 °C: forever | |
| 28S (LSU) | 94 °C: 3 min |
| 35 × −95 °C: 30 s; 55 °C: 45 s; 72 °C: 1 min | |
| 72 °C: 7 min | |
| 4 °C: forever |
Table 4. Sequences of the cnidarian outgroups used in the phylogenetic analyses of Staurozoa, including their GenBank accession numbers.
| Class | Species | Molecular markers | |
|---|---|---|---|
| 18S (SSU) | 28S (LSU) | ||
| Anthozoa | Stichodactyla gigantea | EU190873 | EU190835 |
| Cubozoa | Carybdea rastonii | AF358108 | AY920787 |
| Chironex fleckeri | GQ849073 | GQ849051 | |
| Tripedalia cystophora | GQ849088 | GQ849065 | |
| Hydrozoa | Aglauropsis aeora | AY920754 | AY920793 |
| Scrippsia pacifica | AF358091 | AY920804 | |
| Scyphozoa | Atolla vanhoeffeni | JX393273 | AY026368 |
| Chrysaora melanaster | JX393281 | AY920780 | |
| Phacellophora camtschatica | JX393290 | AY920778 | |
Sequences were edited in SEQUENCHER™ 4.6 (Gene Codes Corporation) or GENEIOUS (Biomatters: available at http://www.geneious.com/), aligned using MAFFT (maxiterate 2.000, FFT-NS-i; Katoh & Standley, 2013), resulting in six alignments: individual COI, 16S, ITS, 18S, 28S, and a combined dataset (Table 5). Gblocks v.0.91b (Castresana, 2000; Talavera & Castresana, 2007) was run with standard parameters except that half the taxa were allowed to be gaps for any position. Gaps were treated as missing data. Parsimony analyses (PA) were performed with individual and combined dataset, using heuristic search (1,000 random addition replicates, with characters reweighted by maximum value of rescaled consistency indices) in PAUP* 4.1 (Swofford, 2002). The most appropriate model of nucleotide substitution for each dataset was chosen using jModelTest (Darriba et al., 2012), between 88 models, using default settings, and employing the Akaike information criterion (using AICc correction). The following models were used in the Maximum Likelihood and Bayesian analyses: COI–HKY+I+G; 16S–TIM2+I+G; ITS–K80+I+G; 18S–TIM2+I+G; 28S–TIM3+I+G; combined–GTR+I+G (no partitioned analyses were conducted). Maximum Likelihood analyses (ML) were performed with individual and combined dataset, using PhyML 3.0 (Guindon et al., 2010). Branch support was estimated by bootstrapping (Felsenstein, 1985) with 1,000 replicates for the PA (PAUP* 4.1) and ML (PhyML) analyses. The Bayesian inference (BA) was also performed with individual and combined dataset, in MrBayes v3.2 (Ronquist & Huelsenbeck, 2003), with 5,000,000 generations sampled every 1,000 generations, four chains, and four independent runs. One fourth of the topologies were discarded as burnin, and the remaining used to calculate the posterior probability. Following MrBayes v3.2 manual, convergence was assessed by ensuring that the average standard deviation of split frequencies was less than 0.01 after 5,000,000 generations, and that the convergence statistic (PSRF = Potential Scale Reduction Factor) was close to 1.0 for all parameters. FigTree (http://tree.bio.ed.ac.uk/software/figtree/) was used to visualize and edit the resulting trees. The alignments and trees are available in the repository of phylogenetic information TreeBASE at: http://purl.org/phylo/treebase/phylows/study/TB2:S18971.
Table 5. Molecular alignments information.
| Alignments | NT | SA | C | V | Pi | S |
|---|---|---|---|---|---|---|
| COI | 27 | 587 | 344 | 243 | 232 | 11 |
| 16S | 35 | 561 | 301 | 260 | 239 | 21 |
| ITS | 32 | 314 | 206 | 108 | 60 | 48 |
| 18S (SSU) | 42 | 1,562 | 1,275 | 287 | 198 | 89 |
| 28S (LSU) | 42 | 3,004 | 2,285 | 719 | 558 | 161 |
| Combined | 45 | 6,028 | 4,411 | 1,617 | 1,287 | 330 |
Note:
NT, number of taxa; SA, size of alignment; C, conserved sites; V, variable sites; Pi, parsimony informative sites; S, singleton sites.
Selected morphological characters generally used in the taxonomy of Staurozoa were optimized by using ACCTRAN (accelerated transformation) in the combined molecular phylogenetic tree at the generic level, using TNT 1.1 (Goloboff, Farris & Nixon, 2008).
Morphology
Detailed images of morphological structures from specimens (Table 6) fixed in 4% formaldehyde solution with seawater were photographed under the stereomicroscope SteREO Discovery.V8, Zeiss (Germany). Histological procedures were carried out according to the methods developed for Staurozoa (Miranda, Collins & Marques, 2013; modified from Humanson (1962) and Mahoney (1966)). Specimens were cleaned in distilled water; dehydrated in a graded ethanol series (70–100%); cleared in xylene (three steps); infiltrated and embedded in paraffin; serially sectioned transversely (7.0–10.0 μm thick) with a microtome Leica RM2025; cleared in xylene (twice); rehydrated in a graded ethanol series (100-70%); cleaned in distilled water; and stained, using acid fuchsin (15′) (Mallory; Humanson, 1962: 147) and acetic aniline blue (3′) (Mallory; modified from Humanson (1962: 231)), intercalated with distilled water to improve the contrast between structures. Prepared slides were observed and photographed under a microscope Axio Imager M2, Carl Zeiss (Germany).
Table 6. Species of Staurozoa used in the detailed morphological descriptions, with respective localities, voucher catalog numbers, and slides catalog numbers.
| Species | Locality | Voucher catalog number | Slides catalog number |
|---|---|---|---|
| Craterolophus convolvulus | Woods Hole, Massachusetts, USA | USNM 54321 | LEM 17 |
| Depastromorpha africana | Kalk Bay, Cape Town, South Africa | MZUSP 002733 | – |
| Haliclystus tenuis | Muroran, Hokkaido, Japan | USNM 1106652 | LEM 09 |
| Kishinouyea corbini | Aracruz, Espírito Santo, Brazil | MZUSP 1563 | LEM 14 |
| Kishinouyea sp. NZ | Taputeranga Marine Reserve, Wellington, New Zealand | NIWA 86808 | LEM 18 |
| Lipkea sp. Japan | Aquarium, Katsuura, Chiba, Japan | USNM 1315325 | – |
| Lucernariopsis campanulata | Île Verte, Roscoff, France | USNM 1233741 | – |
| Lucernariopsis cruxmelitensis | Wembury, Plymouth, England | USNM 1233742 | – |
| Lucernariopsis tasmaniensis | Gerloff Bay, South Australia, Australia | USNM 1233740 | – |
| Lucernariopsis vanhoeffeni | Janus Island, Palmer Archipelago, Antarctica | USNM 79939 | – |
| Manania uchidai | Muroran, Hokkaido, Japan | USNM 1106645 | LEM 10 |
| Sasakiella cruciformis | Muroran, Hokkaido, Japan | USNM 1106656 | LEM 15 |
Note:
LEM, Laboratory of Marine Evolution of the Institute of Biosciences, University of São Paulo; MZUSP, Museum of Zoology of the University of São Paulo; NZ, New Zealand; NIWA, National Institute of Water and Atmospheric Research; USNM, National Museum of Natural History, Smithsonian, USA.
Results and Discussion
Phylogeny
The PA, ML, and BA topologies based on combined markers are similar (Figs. 3–5). The main difference is the relationships among Lucernariopsis vanhoeffeni, Lucernariopsis campanulata, and Kishinouyea sp. NZ (Figs. 3–5) and the relationships among Kishinouyea corbini, Lucernariopsis tasmaniensis, and Kishinouyea sp. SAF. Single-gene topologies under PA, ML, and BA show varying levels of correspondence to the combined topology (Figs. S1–S15 and 6). At least one molecular marker individually supports each main group observed in the PA, ML, and BA results (Fig. 6). This is the most comprehensive molecular phylogenetic hypothesis that has been presented for Staurozoa, which consequently allows us to carry out a comparative analysis of trait distribution across clades, as well as to provide a major revision for the classification of the class (Figs. 7 and 8; Table 7).
Figure 3. Parsimony phylogenetic hypothesis.

Analysis based on combined data of mitochondrial markers COI and 16S, and nuclear markers ITS, 18S (SSU), and 28S (LSU). Single most parsimonious tree, length: 1682.18 steps. Bootstrap indices under parsimony at each node. ANT, Antarctica; AUS, Australia; EPR, East Pacific Rise; GER, Germany; JAP, Japan; NZ, New Zealand; SAF, South Africa; UK, the United Kingdom; USA, the United States of America.
Figure 5. Bayesian phylogenetic hypothesis.

Analysis based on combined data of mitochondrial markers COI and 16S, and nuclear markers ITS, 18S (SSU), and 28S (LSU). Posterior probability at each node. ANT, Antarctica; AUS, Australia; EPR, East Pacific Rise; GER, Germany; JAP, Japan; NZ, New Zealand; SAF, South Africa; UK, the United Kingdom; USA, the United States of America.
Figure 6. Support of each individual molecular marker for the main groups observed in the combined analyses.

White squares represent non monophyletic groups, and gray squares represent monophyletic groups. First row: individual molecular markers under parsimony analyses; second row: individual molecular markers under maximum likelihood analyses; third row: individual molecular markers under Bayesian analyses. PA, parsimony; ML, maximum likelihood; BA, Bayesian. “?” indicates groups whose monophyly could not be corroborate for a particular molecular marker (only one species).
Figure 7. New proposal of classification based on molecular phylogenetic analyses.

“?” indicates groups not included in the analysis, classified according to morphological evidence. EPR, East Pacific Rise; UK, the United Kingdom; USA, the United States of America.
Figure 8. Historical proposals of classifications for Staurozoa.

Classification proposed in this study (F), based on molecular phylogenetic analysis and on additional morphological evidence. In red, new names proposed by the author of respective classification.
Table 7. New proposal for classification of Staurozoa based on the phylogenetic hypotheses (Figs. 3–5 and 7), also considering non-sampled genera (see text for further explanation).
| Upper Rank | Family | Genus | Species | ||
|---|---|---|---|---|---|
| CLASS STAUROZOA Marques & Collins, 2004 | Order Stauromedusae Haeckel, 1879 | Suborder Amyostaurida nov. | Craterolophidae Uchida, 1929 | Craterolophus Clark, 1863 | C. convolvulus (Johnston, 1835)* |
| C. macrocystis von Lendenfeld, 1884 | |||||
| Kishinouyeidae Uchida, 1929 | Calvadosia Clark, 1863 | C. campanulata (Lamouroux, 1815)* | |||
| C. capensis (Carlgren, 1938) | |||||
| C. corbini (Larson, 1980) | |||||
| C. cruciformis (Okubo, 1917) | |||||
| C. cruxmelitensis (Corbin, 1978) | |||||
| C. hawaiiensis (Edmondson, 1930) | |||||
| C. nagatensis (Oka, 1897) | |||||
| C. tsingtaoensis (Ling, 1937) | |||||
| C. tasmaniensis (Zagal et al., 2011) | |||||
| C. vanhoeffeni (Browne, 1910) | |||||
| Suborder Myostaurida nov. | Haliclystidae Haeckel, 1879 | Depastromorpha Carlgren, 1935 | D. africana Carlgren, 1935* | ||
| Depastrum Gosse, 1858 | D. cyathiforme (Sars, 1846)* | ||||
| Haliclystus Clark, 1863 | H. antarcticus Pfeffer, 1889 | ||||
| H. auricula Clark, 1863* | |||||
| H. borealis Uchida, 1933 | |||||
| H. californiensis Kahn et al., 2010 | |||||
| H. inabai (Kishinouye, 1893) | |||||
| H. kerguelensis Vanhöffen, 1908 | |||||
| H. monstrosus (Naumov, 1961) | |||||
| H. octoradiatus Clark, 1863 | |||||
| H. salpinx Clark, 1863 | |||||
| H. “sanjuanensis” nomen nudum | |||||
| H. sinensis Ling, 1937 | |||||
| H. stejnegeri Kishinouye, 1899 | |||||
| H. tenuis Kishinouye, 1910 | |||||
| Halimocyathus Clark, 1863 | H. platypus Clark, 1863* | ||||
| Manania Clark, 1863 | M. atlantica (Berrill, 1962) | ||||
| M. auricula (Fabricius, 1780)* | |||||
| M. distincta (Kishinouye, 1910) | |||||
| M. gwilliami Larson & Fautin, 1989 | |||||
| M. handi Larson & Fautin, 1989 | |||||
| M. hexaradiata (Broch, 1907) | |||||
| M. uchidai (Naumov, 1961) | |||||
| Kyopodiidae Larson, 1988 | Kyopoda Larson, 1988 | K. lamberti Larson, 1988* | |||
| Lipkeidae Vogt, 1886 | Lipkea Vogt, 1886 | L. ruspoliana Vogt, 1886* | |||
| L. stephensoni Carlgren, 1933 | |||||
| L. sturdzii (Antipa, 1893) | |||||
| Lucernariidae Johnston, 1847 | Lucernaria Müller, 1776 | L. australis Vanhöffen, 1908 | |||
| L. bathyphila Haeckel, 1879 | |||||
| L. haeckeli (Antipa, 1892) | |||||
| L. infundibulum Haeckel, 1879 | |||||
| L. janetae Collins & Daly, 2005 | |||||
| L. quadricornis Müller, 1776* | |||||
| L. sainthilairei (Redikorzev, 1925) | |||||
| L. walteri (Antipa, 1892) | |||||
| Stylocoronella Salvini-Plawen, 1966 | S. riedli Salvini-Plawen, 1966* | ||||
| S. variabilis Salvini-Plawen, 1987 | |||||
Note:
Type species for each genus.
Figure 4. Maximum likelihood phylogenetic hypothesis.

Analysis based on combined data of mitochondrial markers COI and 16S, and nuclear markers ITS, 18S (SSU), and 28S (LSU). Bootstrap indices under maximum likelihood at each node. ANT, Antarctica; AUS, Australia; EPR, East Pacific Rise; GER, Germany; JAP, Japan; NZ, New Zealand; SAF, South Africa; UK, the United Kingdom; USA, the United States of America.
Higher level systematics
Suborders Amyostaurida nov. and Myostaurida nov.
The class Staurozoa has traditionally been divided into the subgroups Cleistocarpida and Eleutherocarpida (Fig. 8), based on the presence and absence, respectively, of an internal structure called the claustrum (Fig. 9; Table 8). However, a preliminary phylogenetic analysis for the class (Collins & Daly, 2005) suggested that these groups, proposed by Clark (1863) (Fig. 8), were not monophyletic. Our study, with better taxon sampling, corroborates this preliminary result, and refutes the suborders Eleutherocarpida and Cleistocarpida (Fig. 8). Instead, our working hypothesis based on our combined set of molecular data (Fig. 7) shows that staurozoans are divided into two well-supported groups, which can be distinguished one from the other by the absence/presence of interradial longitudinal muscles in the peduncle (or stalk) (Figs. 10 and 11; Table 8). We propose two new suborders for the Staurozoa: Myostaurida (from the Greek myos: muscle; stauro: cross) and Amyostaurida composed of species with and without interradial muscles in the peduncle, respectively (Figs. 7, 8F, 10 and 11; Table 8). Presence of longitudinal muscles in the peduncle (Figs. 10A and 11) is a character easily recognizable with a cross-section of the middle region of the peduncle (Uchida, 1929; Ling, 1937; Ling, 1939; Berrill, 1963; Miranda, Collins & Marques, 2013), and consequently a useful feature for distinguishing the two major subgroups of stalked jellyfishes (see discussion about character evolution below).
Figure 9. Claustrum connecting adjacent septa.

Craterolophus convolvulus: (A) beginning of claustrum delimitation (indicated by black arrow) between adjacent septa (sp) in peduncle; (B) claustrum (cs) completely delimited at base of calyx, enclosing accessory radial pockets (ar); Manania uchidai: (C) claustrum (cs) completely delimited at base of calyx, enclosing accessory radial pockets (ar); (D) claustrum (cs) between accessory radial pockets (ar) and principal radial pockets (pr) (associated with gonads) in calyx, and a central manubrium (mn); Calvadosia sp. 2 NZ: (E) absence of claustrum connecting adjacent septa (sp) in peduncle; (F) absence of claustrum at the base of calyx between adjacent septa (sp); (G) gastric radial pocket (gp) associated with gonads (gd). Cross-sections. Photo credit: Lucília Miranda.
Table 8. Main morphological characters used in the taxonomy of Staurozoa and their occurrence in each genus.
| Staurozoan genera | ||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Characters | States | Craterolophus | Calvadosia | Depastromorpha | Depastrum | Haliclystus | Halimocyathus | Manania | Kyopoda | Lipkea | Lucernaria | Stylocoronella |
| Claustrum | Present | X | X | X | X | X | ||||||
| Absent | X | X | X | X | X | X | ||||||
| Muscles in peduncle | Present | X | X | X | X | X | X | X | X | X | ||
| Absent | X | X | ||||||||||
| Number of chambers in peduncle | 4 | X | X | X | X | X | X | ? | ||||
| 1 | X | X | X | X | X | |||||||
| 4 basal, 1 medial | X | |||||||||||
| 1 basal, 4 medial | X | |||||||||||
| Anchors and primary tentacles | Absent | X | X | ? | X | |||||||
| Primary tentacles | X | X | Migration (see text) | |||||||||
| Anchors | X | X | X | X | X | |||||||
| Pad-like adhesive structures | Absent | X | X | X | X | X | ||||||
| Tentacles | X | X | X | X | X | X | X | |||||
| Arms | X | |||||||||||
| Coronal muscle | Divided | X | X | X | X | |||||||
| Entire | X | X | X | ? | X | X | X | |||||
| Vestigial | X | |||||||||||
Note:
Based on Gosse (1860), Clark (1863), Mayer (1910), Uchida (1929), Uchida & Hanaoka (1933), Uchida & Hanaoka (1934), Carlgren (1935), Ling (1937), Kramp (1961), Larson (1980), Larson (1988), Hirano (1986), Hirano (1997), Larson & Fautin (1989), Kikinger & Salvini-Plawen (1995) and Pisani et al. (2007).
Figure 10. Interradial longitudinal muscles in peduncle.

Manania uchidai: (A) presence of interradial longitudinal muscles (mu); Calvadosia cruciformis: (B) absence of interradial longitudinal muscles (indicated by arrow). Cross-sections. Photo credit: Lucília Miranda.
Figure 11. Hypothesis of character evolution for staurozoan genera.

ACCTRAN optimization of selected morphological and life-history features according to our molecular phylogenetic analyses. Synapomorphies and symplesiomorphies are based on Collins & Daly (2005). The presence of claustrum as a potential symplesiomorphy of Staurozoa (Collins & Daly, 2005) is equivocal, and the state in outgroups needs careful reconsideration based on detailed histological studies. If considered a symplesiomorphy of Staurozoa, claustrum was lost in Calvadosia, Haliclystus, and in the clade Lucernaria + Lipkea (most parsimonious reconstruction). Anchors are adhesive structures resulting from metamorphosis of eight primary tentacles (perradial and interradial). Coronal muscle divided into eight sections by the adradial arms or entire. The species with 4/1-chambered peduncle have four chambers basally and one chamber in the middle of the peduncle.
Family Craterolophidae Uchida, 1929
Type genus: Craterolophus Johnston, 1835
Craterolophinae was proposed by Uchida (1929) (Fig. 8C) as a subfamily of Cleistocarpidae, defined as stauromedusae with claustrum and without longitudinal interradial muscles in the peduncle (Figs. 9 and 10; Table 8). This classification was followed by Carlgren (1935) (Fig. 8D). The subfamily is monogeneric and contains only two valid species: Craterolophus convolvulus (Johnston, 1835) and Craterolophus macrocystis von Lendenfeld, 1884.
We followed Daly et al. (2007) and elevated Craterolophinae to the family level, as Craterolophidae (Figs. 7, 8E and 8F), including only the genus Craterolophus (Figs. 7 and 8; Table 7). We included specimens of C. convolvulus from Europe (Germany and the United Kingdom) and from the U.S.A. (Table 1) in our analysis. However, there was no specimen available of C. macrocystis; the species is very rare, having been recorded only twice (Hutton, 1880; von Lendenfeld, 1884). Therefore, the monophyly of the genus and, consequently, the family, remains to be tested.
Family Kishinouyeidae Uchida, 1929
Type genus: Calvadosia Clark, 1863
The family Kishinouyeidae was proposed by Uchida (1929) to include the genera Kishinouyea and Sasakiella (Fig. 8C). Carlgren (1935) proposed an amendment to also include the genus Lucernariopsis (Fig. 8D).
The monophyly of the family was tested and corroborated in our analysis (Figs. 3–5). However, the two traditional genera Lucernariopsis and Kishinouyea did not resolve as monophyletic (Figs. 3–5). According to current taxonomy, the distinction between the three genera of this family is subtle. Kishinouyea and Sasakiella differ by the absence and presence, respectively, of primary tentacles (Ling, 1937). Both Kishinouyea and Lucernariopsis do not have primary tentacles in adults, but they are thought to differ in the internal anatomy of the peduncle. Whereas species of Kishinouyea (and Sasakiella) have four chambers basally and one chamber in the middle of the peduncle, species of Lucernariopsis have just one chamber throughout the peduncle (Uchida, 1929; Kramp, 1961). However, these characters change during development (Uchida, 1929; Hirano, 1986). Additionally, a cross-section at the very base of the peduncle is rarely reported in the description of species; most only include information concerning the middle region of the peduncle (e.g., Kishinouyea hawaiiensis in Edmondson, 1930; Lucernariopsis capensis in Carlgren, 1938; Miranda et al., 2012), or do not mention where the peduncle was sectioned (e.g., Corbin, 1978), causing some doubt about whether this distinction is reliable in defining these genera. Recently, Lucernariopsis tasmaniensis was described with “a single cruciform chamber that becomes four-chambered basally within pedal disc” (Zagal et al., 2011), a character that corresponds to the genera Kishinouyea and Sasakiella (Kramp, 1961). Not surprisingly, our phylogenetic hypothesis (Figs. 3–5) indicates that the traditional distinctions between these genera are not robust.
We suggest that the three genera of Kishinouyeidae be synonymized due to the lack of characters to differentiate them. Kishinouyea Mayer, 1910 would have priority over Lucernariopsis Uchida, 1929 and Sasakiella Okubo, 1917. However, there is a further nomenclatural problem in Uchida’s (1929) proposal of the genus Lucernariopsis based on Lucernaria campanulata (Lamouroux, 1815; Gwilliam, 1956: 10). Previously, Clark (1863) had recognized Lamouroux’ species as not assignable to Lucernaria, since the species does not have interradial muscles in the peduncle, and proposed the new genus name Calvadosia (non Calvadosia Cossmann 1921; junior synonym of Calvadosiella Wenz 1939; Mollusca, Gastropoda) to accommodate it. Thus, following the rule of priority, the proper generic name of Lucernariopsis Uchida, 1929 would be Calvadosia Clark, 1863. Consequently, Calvadosia has priority over Kishinouyea Mayer, 1910, and we therefore synonymize Kishinouyea, Sasakiella, and Lucernariopsis within Calvadosia. The name of the family remains the same, according to ICZN, article 40.1.
Family Haliclystidae Haeckel, 1879
Type genus: Haliclystus Clark, 1863
Haliclystidae was proposed by Haeckel (1879) as a subfamily of Lucernaridae, including the genera Haliclystus and Lucernaria (Fig. 8B). Uchida (1929) raised Haliclystidae to the family level, adding the genera Stenoscyphus, Capria (currently regarded as congeneric with Lipkea), and Lucernariopsis (Fig. 8C). The family was dismissed by Carlgren (1935), who divided the genera of “Haliclystidae” into three subfamilies of Eleutherocarpidae: Lucernariinae (Haliclystus, Stenoscyphus, and Lucernaria), Lipkeinae (Lipkea), and Kishinouyiinae (Lucernariopsis) (Fig. 8D).
Our phylogenetic analyses show a close relationship between Haliclystus, Stenoscyphus, Depastromorpha, and Manania (Figs. 3–5). Based on this evidence and on morphological similarities (see below, Table 8), we propose that these genera should be assigned to the family Haliclystidae (Figs. 7 and 8F; Table 7). We also include in this family the not yet sampled genera Depastrum and Halimocyathus, but this needs to be tested in future studies.
According to the phylogeny, Stenoscyphus inabai is closely related to Haliclystus borealis and Haliclystus tenuis (Figs. 3–5), and deeply nested within Haliclystus spp. In order to keep Haliclystus monophyletic, and since the name Haliclystus Clark, 1863 has priority over the name Stenoscyphus (a monospecific genus) Kishinouye, 1902, we synonymize Stenoscyphus with Haliclystus (Figs. 7 and 8F; Table 7). Some limited developmental data has already suggested a close relationship between these two genera (Hirano, 1986). The main difference between the former genus Stenoscyphus and Haliclystus is an entire and divided coronal muscle, respectively (Kramp, 1961; Hirano, 1986). Therefore, Haliclystus inabai is the only described Haliclystus with an entire coronal muscle (Table 8).
Genetic data suggest that Depastromorpha is more closely related to Haliclystus than to Manania (Figs. 3–5). Both Depastromorpha and Manania possess the claustrum (Figs. 9 and 11), a structure also present in Depastrum and Halimocyathus in the family Haliclystidae (Table 8) (Clark, 1863; Carlgren, 1935; Kramp, 1961), suggesting that this structure may have been lost in the lineage leading to Haliclystus (Figs. 7 and 11; ACCTRAN optimization). In general, Depastrum, Depastromorpha, Manania, and Halimocyathus have similar morphologies, in addition to the presence of claustrum. For example, Depastrum, Depastromorpha, and Manania (and probably Halimocyathus, see Clark, 1863) have an entire coronal muscle (Table 8; Carlgren, 1935; Kramp, 1961); Manania, Depastromorpha, and Halimocyathus have pad-like adhesive structures in the outermost secondary tentacles (Table 7; Clark, 1863; Carlgren, 1935; Kramp, 1961; Larson & Fautin, 1989; Zagal et al., 2011); and Depastrum and Manania have coronal muscle on the exumbrellar (external) side of the primary tentacles/anchors (Fig. 12), unlike all the other genera of stalked jellyfishes (although the condition in Halimocyathus is uncertain) (Carlgren, 1935).
Figure 12. Coronal muscle.

Craterolophus convolvulus: (A) divided coronal muscle (cm); Lipkea sp. Japan: (B) entire coronal muscle (cm); Manania uchidai: (C) external (exumbrellar) coronal muscle (cm) in relation to anchor (an); Depastromorpha africana: (D) internal (subumbrellar) coronal muscle in relation to anchor (an). Photo credit: Lucília Miranda.
Most species of the family Haliclystidae have primary tentacles that metamorphose into anchors (Figs. 11 and 13). In the genera Manania and Depastromorpha, there is a knobbed remnant of each primary tentacle, with a glandular pad-like adhesive structure at the base (Figs. 13C and 13D) (Carlgren, 1935; Larson & Fautin, 1989; Zagal et al., 2011). However, the anchors in Manania are small and sometimes referred to as primary tentacles (Naumov, 1961; Larson & Fautin, 1989). In Halimocyathus, the anchors were described as “pistilliform,” “very small,” with “uniform thickness from the knob to the base” (Clark, 1863: 536, 538), but broader than the secondary tentacles (Mayer, 1910), so they are probably similar to the anchors in Manania, but possibly even more diminutive. In Haliclystus, the transformation of the primary tentacles into anchors is more obvious (Fig. 13E) (Clark, 1863; Hirano, 1986; Miranda, Morandini & Marques, 2009), although a knobbed remnant of the primary tentacles can be observed in some species (Clark, 1878; Miranda, Morandini & Marques, 2009; Kahn et al., 2010). However, the genus Depastrum seems to be an exception, with unmetamorphosed perradial and interradial primary tentacles (Clark, 1863; Mayer, 1910) (Table 8).
Figure 13. Primary tentacles and anchors.

Craterolophus convolvulus: (A) absence of primary tentacles and anchors (indicated by black arrow) between arms; Calvadosia cruciformis: (B) presence of primary tentacles (pt); Manania uchidai: (C) anchors (an) with a knobbed remnant of primary tentacles; Depastromorpha africana: (D) anchors (an) with a knobbed remnant of primary tentacles; Haliclystus tenuis: (E) anchors (an). Photo credit: Lucília Miranda.
Based on morphological evidence, we include Depastrum Gosse, 1858 and Haliclystus Clark, 1863 in the same family (Figs. 7 and 8F; Table 7). However, there is a nomenclatural issue related to these genera. Haeckel (1879) proposed both the subfamilies Depastridae and Haliclystidae in the same book (Fig. 8B). Both names were used by Uchida (1929), but Carlgren (1935), Kramp (1961) and Daly et al. (2007) used only Depastrinae/Depastridae, and replaced Haliclystidae by Lucernariidae (Fig. 8). Consequently, the prevailing name would be Depastridae. However, there are two caveats: (1) Depastrum cyathiforme, the single species of the genus (Table 7), is not sampled in this study and consequently its position in the phylogeny (i.e., its relationship with other genera) is more tentative (Fig. 7; based only on morphological similarities); and (2) the last report of D. cyathiforme in the literature was about 40 years ago (den Hartog, 1976). Therefore, we believe it is better for nomenclatural stability to use the name Haliclystidae over Depastridae, and as first revisers refer to the International Code on Zoological Nomenclature (ICZN), article 24.2.2.
Family Kyopodiidae Larson, 1988
Type genus: Kyopoda Larson, 1988
The Kyopodiidae is a monospecific family proposed by Larson (1988) as part of Eleutherocarpida. Kyopoda lamberti Larson, 1988 has an unusual morphology: its calyx is reduced and the gonads and gastric cavity reside at the base of the peduncle (Larson, 1988).
There was no specimen available of K. lamberti to be included in our phylogenetic analyses. In addition, its particular morphology hampers attempts to identify a relationship with other genera of Staurozoa, which makes future study focusing on the homologies of K. lamberti with other Staurozoa especially interesting. Therefore, we presently retain the monogeneric family Kyopodiidae and assign it to the suborder Myostaurida (Figs. 7 and 8F; Table 7) because K. lamberti has interradial longitudinal muscles associated with the infundibula (Larson, 1988).
Family Lipkeidae Vogt, 1886
Type genus: Lipkea Vogt, 1886
The monogeneric family Lipkeidae was proposed by Vogt (1886) and presently encompasses three species: Lipkea ruspoliana Vogt, 1886, Lipkea sturdzii (Antipa, 1893), and Lipkea stephensoni Carlgren, 1933 (cf. Carlgren, 1935; Kramp, 1961; Daly et al., 2007) (Table 7). Lipkea is closely related to Lucernaria (Figs. 3–5), but there are enough characters to easily distinguish these two genera (Table 8) and we retain Lipkea as the exclusive genus of Lipkeidae (Fig. 8F; Table 7).
Family Lucernariidae Johnston, 1847
Type genus: Lucernaria Müller, 1776
The family Lucernariidae was proposed by Johnston (1847), including only the genus Lucernaria. Whereas Clark (1863) used the name Lucernariae for all of Stauromedusae, Haeckel (1879) was actually the originator of the name Stauromedusae, in which he placed the family Lucernariidae, divided into two subfamilies: 1) Haliclystidae, including the genera Haliclystus and Lucernaria; and 2) Halicyathidae, including Halicyathus (=Halimocyathus) and Craterolophus (Fig. 8B). Carlgren (1935) proposed Lucernariinae as a subfamily of Clark’s (1863) family Eleutherocarpidae, including Lucernaria, Haliclystus, and Stenoscyphus (Fig. 8D), and a similar classification was used by Kramp (1961). Kikinger & Salvini-Plawen (1995), and then Daly et al. (2007), used Lucernariidae as a family of suborder Eleutherocarpina and suborder Eleutherocarpida, respectively, including the genera Haliclystus, Stenoscyphus, Lucernaria, and Stylocoronella (Fig. 8E). However, the topologies presented by Collins & Daly (2005) contradicted monophyly of this grouping (cf. Haeckel, 1879, i.e., when including at least Lucernaria and Haliclystus), a pattern corroborated in our results (Figs. 3–5). Accordingly, we propose that Lucernariidae be limited to the genera Lucernaria and Stylocoronella (Figs. 7 and 8F; Table 7). This hypothesis has to be tested further because Stylocoronella has not yet been available for inclusion in our molecular-based phylogenetic analysis (Fig. 7), but it is consistent with the morphological similarities of Lucernaria and Stylocoronella (Table 8). Kikinger & Salvini-Plawen (1995) superficially remarked that Stylocoronella spp. appear to be congeneric with Lucernaria, although they presented a fundamental difference concerning the fate of the primary tentacles. In Lucernaria, the primary tentacles reduce to absent through development (Berrill, 1962), whereas in Stylocoronella the primary tentacles are retained (Table 8) and become integrated among the adradial clusters of the secondary tentacles (Kikinger & Salvini-Plawen, 1995). However, this developmental difference cannot be distinguished in adults, making its application difficult. Additionally, the coronal muscle seems to be vestigial in Stylocoronella (Table 8) (Kikinger & Salvini-Plawen, 1995), but this information needs further observations.
Character state evolution
Stalked jellyfishes have relatively few external characters useful for taxonomy (Hirano, 1997). Consequently, some internal features are also employed to differentiate these animals (Uchida, 1929; Ling, 1937; Ling, 1939; Miranda, Collins & Marques, 2013). However, most of these characters vary intraspecifically and ontogenetically and they have to be assessed and cautiously employed to differentiate species (Miranda, Morandini & Marques, 2009). We review the main characters used in the traditional taxonomy of Staurozoa (Table 8) and interpret their significance based on the new phylogenetic hypothesis for the class (Figs. 3–5 and 7; Table 7).
Claustrum
The claustrum (Fig. 9) is a membrane that divides the gastrovascular cavity (Clark, 1863; Gross, 1900) of some stauromedusae (Table 8) and represents an additional level of complexity of their gastrovascular system (Berrill, 1963; Collins & Daly, 2005). Stauromedusae with claustrum have eight gastric radial pockets in the calyx (Fig. 9; Gross, 1900; Berrill, 1963). The four external pockets, known as accessory radial pockets (or exogon pockets; Thiel, 1966), extend into the marginal tentacles and anchors, continuing into the peduncle as the gastric chambers (Berrill, 1963). The four internal pockets, known as principal radial pockets (or mesogon pockets; Thiel, 1966), are the true radial pockets of these stauromedusae because they contain the gonads, as do the four gastric radial pockets of species without claustrum (Clark, 1863; Gross, 1900; Berrill, 1963). Clark (1863) proposed that the stalked jellyfishes should be divided into two main groups based on the presence and absence of the claustrum, respectively: Cleistocarpidae and Eleutherocarpidae (Fig. 8A). Since then, the claustrum has played an important role in the systematics of stauromedusae (Collins & Daly, 2005) and the main classifications have been based on this character, although with different levels of importance (Clark, 1863; Haeckel, 1879; Gross, 1900; Uchida, 1929; Carlgren, 1935; Gwilliam, 1956; Kramp, 1961; Uchida, 1973) (Fig. 8).
A preliminary phylogeny based on nuclear and mitochondrial molecular markers suggested that neither Cleistocarpida nor Eleutherocarpida are monophyletic and that the claustrum “is a more labile feature than suspected and that it may have been lost on more than one occasion,” and should not be used to diagnose subgroups within the class Staurozoa (Collins & Daly, 2005: 229). These conclusions are corroborated by our analysis (Figs. 3–5 and 11; Table 8). Most of the genera in the family Haliclystidae (suborder Myostaurida) have claustrum (Depastromorpha, Depastrum, Halimocyathus, and Manania), except the type genus Haliclystus (Fig. 8F; Tables 7 and 8). In addition, species of Craterolophus, family Craterolophidae (suborder Amyostaurida), also have claustrum (Tables 7 and 8), indicating a homoplastic character (Fig. 11).
Claustrum has also been described in the medusa stage of Cubozoa (Thiel, 1966). However, the internal organization of this structure is different between Staurozoa and Cubozoa (gonads associated with the exogon in Cubozoa; Thiel, 1966), and the existence of a typical staurozoan claustrum in Cubozoa is doubtful (Thiel, 1966). Therefore, if the claustrum in Staurozoa is not homologous to the structure in Cubozoa, claustrum appeared at least twice in the evolution of stalked jellyfishes, and it was lost in Haliclystus (Fig. 11, ACCTRAN). Alternatively, if considered a symplesiomorphy of Staurozoa (Collins & Daly, 2005), claustrum was lost in Calvadosia, Haliclystus, and in the clade Lucernaria + Lipkea (most parsimonious reconstruction).
Interradial longitudinal muscles in the peduncle
The stalked jellyfishes can have four interradial longitudinal muscle bundles, formed by epitheliomuscular cells, in the peduncle (Fig. 10A) (Miranda, Collins & Marques, 2013). These muscles have been generally used to distinguish genera and families of Stauromedusae (Table 8). Clark (1863), for example, distinguished the genus Calvadosia from Lucernaria based on the absence and presence of these muscles, respectively. Uchida (1929) separated stauromedusae without claustrum into three families, one of them (Kishinouyeidae) without muscles in the peduncle. At the same time, Uchida (1929) divided stauromedusae with claustrum into two subfamilies, Depastrinae with muscles in the peduncle, and Craterolophinae without these muscles (Fig. 8C).
Additionally, Uchida (1929) proposed using the shape of the muscle in the peduncle as seen in cross-section as a specific character of Haliclystus stejnegeri in relation to its congeners. Gwilliam (1956: 7) accepted the use of the muscular system to differentiate higher hierarchical levels (e.g., genera and families), but considered it virtually impossible to apply at the specific level due to considerable intraspecific variation, and because the shape depends on both the size (age) and degree of contraction of a given specimen.
Accordingly, the muscles in the peduncle have been treated inconsistently in classification schemes for Staurozoa. For instance, Uchida (1929) assigned Kishinouyea and Sasakiella to the family Kishinouyeidae, but incongruously assigned Lucernariopsis to the Haliclystidae, where it stands out by being the only other genus in the family without muscles in the peduncle (Fig. 8C). Finally, Uchida (1973) clearly considered the presence of claustrum as more important than the muscles in the peduncle in classification.
Our phylogenetic hypothesis reveals that Staurozoa can be divided into two main clades (Figs. 3–5): one only with species possessing the four interradial longitudinal muscles in the peduncle, and the other exclusively formed by species without interradial longitudinal muscles in the peduncle (Table 8). Accordingly, we propose two new suborders for class Staurozoa, order Stauromedusae based on the presence and absence of interradial longitudinal muscles in the peduncle, suborder Myostaurida and Amyostaurida, respectively (Figs. 7 and 8F; Table 7).
Collins et al. (2006) inferred that four interradial, intramesogleal longitudinal muscles associated with peristomial pits (infundibula) were symplesiomorphic in Staurozoa, and shared by the ancestral staurozoan with some (but not all) other medusozoans, a hypothesis we have used in our reconstruction (Fig. 11). Four intramesogleal muscles are characteristic of polyps of scyphozoans (Thiel, 1966; Marques & Collins, 2004; Collins & Daly, 2005). Cubopolyps also possess intramesogleal muscles, though the number is not fixed (Chapman, 1978; Marques & Collins, 2004). In hydropolyps, the musculature consists of a layer of longitudinal epidermal muscular fibers and circular gastrodermal fibers (Marques & Collins, 2004). According to this hypothesis, the longitudinal interradial muscles in the peduncle were lost in the clade Amyostaurida (Fig. 11). Additional clues to understand the likely evolutionary polarity of this character could come from detailed examination of its ontogenetic origins across Staurozoa. However, few stauropolyps have ever been studied (Wietrzykowski, 1912; Kikinger & Salvini-Plawen, 1995), and there is no information concerning the presence/absence of interradial longitudinal muscles in developing stauropolyps of Amyostaurida.
Chambers in the peduncle
The peduncle of stauromedusae can have four perradial chambers delimited by gastrodermis (Fig. 14A) (Miranda, Collins & Marques, 2013), which are connected apically to the gastrovascular system of the calyx (Berrill, 1963). The number of chambers in the peduncle has been one of the characters most used in the literature to distinguish staurozoan genera (Clark, 1863; Mayer, 1910; Uchida, 1929; Kramp, 1961). The animals can either have one chamber in the peduncle (e.g., Lucernaria; Kramp, 1961); four chambers (e.g., Haliclystus; Kramp, 1961); four chambers in lower section of the peduncle, which fuse to form one chamber medially (e.g., Kishinouyea; Mayer, 1910); or one chamber in lower position with four chambers medially (e.g., some Manania, Larson & Fautin, 1989) (Table 8). When animals have four chambers in the medial position of the peduncle, these chambers fuse apically at the transition between peduncle and calyx (Uchida & Hanaoka, 1933; Miranda, Collins & Marques, 2013). Also, the number of chambers in the peduncle appears to vary during development of different species (Mayer, 1910; Uchida, 1929; Hirano, 1986), which makes its interpretation more complex. For instance, Wietrzykowski (1911) and Wietrzykowski (1912) observed Haliclystus octoradiatus with one chamber until the stage of 32 tentacles, when, progressively, four independent chambers are formed upward. This pattern was later observed in different species of Haliclystus, whose juveniles have a single-chambered peduncle, later divided into four chambers from the base to the top of the peduncle (Hirano, 1986).
Figure 14. Chambers in the peduncle.

Haliclystus tenuis: (A) four perradial chambers (pc) in peduncle; Calvadosia corbini: (B) one central gastric chamber (indicated by arrow) in the middle region of peduncle. Cross-sections. Photo credit: Lucília Miranda.
Manania is probably the taxon with the widest variation concerning the number of peduncular chambers (Table 8): four chambers were reported throughout the peduncle in Manania distincta, Manania gwilliami, and Manania handi (Kishinouye, 1910; Larson & Fautin, 1989); four chambers medially and one chamber basally (the lower portion of the peduncle) in Manania atlantica and Manania uchidai (Naumov, 1961; Berrill, 1962); and one chamber throughout the peduncle in Manania auricula (Clark, 1863) and Manania hexaradiata (Broch, 1907; Kramp, 1961; Naumov, 1961). However, as the number of chambers in the peduncle in some Manania species is known to vary with ontogeny (Uchida, 1929; Hirano, 1986), the number of chambers is not a robust character to differentiate species and even staurozoan genera. For example, Clark (1863) considered Halimocyathus sufficiently different from Manania, both taxa described by him. One important difference in his descriptions is the four-chambered peduncle in the former, and single-chambered in the latter. However, different species of Manania were also later described with a four-chambered peduncle (Larson & Fautin, 1989). Therefore, as a general rule, even though the number of chambers in the peduncle seems to be an important character, it should be cautiously employed in the taxonomy of staurozoans (Uchida, 1929; Hirano, 1986).
There have also been some misinterpretations of the number of chambers in the peduncle, making it more difficult to employ this character in taxonomy. Calvadosia nagatensis (Mayer, 1910) and Calvadosia hawaiiensis (Edmondson, 1930) were reported with a four-chambered peduncle, but in fact they have one cruciform chamber throughout the peduncle and only at the level of the pedal disk can the four chambers be observed, sometimes separated by an axial canal (Uchida, 1929; Ling, 1939; Larson, 1980). In another example, Haliclystus was suggested to be closely related to Lucernaria because Haliclystus antarcticus and species of Lucernaria were reported to have a single chamber in the peduncle (Mayer, 1910: 536). In actuality, H. antarcticus has four chambers in the peduncle (Pfeffer, 1889; Carlgren, 1930; Miranda, Collins & Marques, 2013).
Ontogenetic data led Uchida (1929: 153) to hypothesize that “the single-chambered condition of the peduncle is more primitive than the four-chambered one.” However, there is a broad occurrence of four chambers in peduncles of Staurozoa, present at least in Craterolophus, Depastromorpha, Depastrum, Haliclystus, Halimocyathus, and some Manania, and this state would be a potential synapomorphy of Staurozoa (Fig. 11, ACCTRAN), as the four perradial chambers in the peduncle of stalked jellyfishes are not found in any other cnidarian life history stage (Collins & Daly, 2005).
Anchors (rhopalioids) and primary tentacles
During the early development of a stauropolyp, eight primary tentacles develop, four perradial and four interradial (Wietrzykowski, 1912; Hirano, 1986; Kikinger & Salvini-Plawen, 1995), which are probably homologous to the primary tentacles present in other medusozoans (Fig. 11; Uchida, 1929; Thiel, 1966). During the metamorphosis of a stauropolyp into an adult stauromedusa, these eight primary tentacles can have four different developmental fates: 1) they disappear by resorption (Berrill, 1963); 2) they metamorphose into adhesive structures called anchors (Hirano, 1986); 3) they remain as primary tentacles but with a modified shape (Ling, 1937); 4) they change their shape (filiform to capitate), migrate and cluster together with the secondary tentacles (Kikinger & Salvini-Plawen, 1995) (Fig. 13; Table 8).
In many species, primary tentacles are present in juvenile stauromedusae, but disappear during development (Uchida, 1929; Berrill, 1962; Larson, 1980). This suggested that an “erratic occurrence of these primary tentacles (…) indicates that they are negligible as diagnostic characters and of small significance” (Elmhirst, 1922: 221, also highlighted by Uchida, 1929: 150). There is fragmented information about this character, at least partly for a widespread lack of observation of young specimens of most species: Lamouroux (1815) reported that primary tentacles are sometimes observed in C. campanulata, probably in juveniles and in abnormal individuals; Uchida (1929), Ling (1939) and Larson (1980) reported the presence of rudiments of primary tentacles in very young specimens of Calvadosia nagatensis and Calvadosia corbini, as was also observed in Craterolophus convolvulus (Gross, 1900; Carlgren, 1935) and in species of Lucernaria (Berrill, 1963; Collins & Daly, 2005).
In some cases, the eight primary tentacles can also be retained throughout the life of the specimen (Fig. 13) and this condition was distinctive for the former genus Sasakiella (Ling, 1937), which comprised two species, presently Calvadosia tsingtaoensis and Calvadosia cruciformis (Table 7). These two species are differentiated by the number of primary tentacles retained, four in perradial positions in C. tsingtaoensis, and eight, in both the perradii and interradii, in C. cruciformis (Ling, 1937: 15). There may be, however, intraspecific variation for the character, probably related to development: in “a few extreme cases examined the four perradial primary tentacles [of C. cruciformis] are clearly seen but the four interradial ones are reduced to short rudiments. In young specimens all eight of them are well developed” (Ling, 1937: 19).
The development of Stylocoronella riedli and Stylocoronella variabilis shows that the primary filiform tentacles persist in these species, but are transformed into capitate tentacles and clustered together with the secondary tentacles at the tips of the adradial arms (Kikinger & Salvini-Plawen, 1995), a condition never reported in other genera of stalked jellyfishes (Table 8).
Particular marginal structures are also found in Lipkea, a genus morphologically quite distinct from all other stauromedusae (Uchida, 1929: 151) (Fig. 1N). Species of Lipkea have a variable number of lobes (or lappets) at the margin of the calyx (Pisani et al., 2007). Lipkea ruspoliana was described with perradial and interradial lobes, which were suggested to be homologous to the eight primary tentacles, not to the arms of other stauromedusae that are normally adradial (Uchida, 1929). According to this hypothesis, lobes would be highly metamorphosed primary tentacles (Uchida, 1929). However, L. sturdzii and L. stephensoni were described with adradial lobes (Antipa, 1893; Carlgren, 1933). The homology between lobes and primary tentacles was then questioned by Carlgren (1933), who referred to the lobes as modified arms, which was subsequently followed by the description of Lipkea with adradial marginal lobes and without perradial and interradial anchors (Kramp, 1961). Recently, the lobes of L. ruspoliana have been interpreted to be modified tentacles, with an adradial position (Pisani et al., 2007). We consider that the homology of these structures is still under debate, demanding further investigation, particularly of their development.
Primary tentacles can also metamorphose into anchors, adhesive structures that allow momentary adhesion to the substrate through their abundant glandular and supporting cells (Uchida, 1929; Hyman, 1940; Franc, 1994; Miranda, Collins & Marques, 2013). Species of Haliclystidae tend to have the primary tentacles metamorphosed totally or partially (i.e., with a knobbed remnant of the primary tentacles, Figs. 13C–13E) into anchors (Figs. 7 and 11; Tables 7 and 8).
The shape of anchors has frequently been used in the taxonomy of Haliclystus (Gwilliam, 1956; Miranda, Morandini & Marques, 2009; Kahn et al., 2010). However, their morphology has intraspecific and ontogenetic variation, and consequently it must be carefully assessed when employed to differentiate species of the genus (Miranda, Morandini & Marques, 2009; Kahn et al., 2010).
Pad-like adhesive structures
Pad-like structures can be present individually in the outermost secondary tentacles of the tentacular cluster (Larson & Fautin, 1989), or as a broad structure on the tip of each arm (Larson, 1980; Miranda et al., 2012) (Fig. 15; Table 8). Apparently, the pads help the animal to adhere to its substrate. Calvadosia corbini was observed in situ attached to algae by the pedal disk or by the pad-like adhesive structures on the arms’ tips (Larson, 1980). In aquaria, C. corbini mainly use the pads to attach to the substratum, and the relatively large size of the pad compared to the pedal disk makes the importance of this structure for attachment clear (Larson, 1980). The glandular pads located on the anchor and on the abaxial tentacles of Kyopoda lamberti were hypothesized to temporarily serve to reattach the stauromedusae if it becomes detached (Larson, 1988).
Figure 15. Pad-like adhesive structures.

Calvadosia tasmaniensis: (A–B) pad (pa) on the tip of an arm separate from the secondary tentacles (tc); Calvadosia cruxmelitensis: (C) pad (pa) on the tip of an arm, with secondary tentacles (tc) arising directly from it; Craterolophus convolvulus: (D) pads (pa) in the outermost secondary tentacles (tc); Calvadosia vanhoeffeni: (E) pads (pa) in the outermost secondary tentacles (tc); Calvadosia campanulata: (F) pads (pa) in the outermost secondary tentacles (tc). Photo credit: Lucília Miranda.
There is only scattered information on the ontogeny of the pad-like adhesive structures. They apparently appear in the outermost tentacles late in development of C. cruciformis (Hirano, 1986: 197). Also, the broad adhesive pad-like structure on the tip of each arm hypothetically results from the fusion of several secondary outermost tentacles in C. corbini (Larson, 1980). Pad-like adhesive structures in the outermost tentacles and on the tips of the arms were considered to be homologous by Corbin (1978), but this requires more rigorous study.
This character has already been used to diagnose subfamilies (Carlgren, 1935). However, Carlgren (1935) overlooked the occurrence of pad-like adhesive structures in the outermost tentacles of some species of Haliclystus, which emphasizes the variation of this character within genera (Gwilliam, 1956). The pads in Haliclystus (especially in Haliclystus californiensis; Gwilliam, 1956; Kahn et al., 2010) are never as large as those found in Manania and Calvadosia, but their presence in Haliclystus should be taken into account in considering the relevance of this character for taxonomy.
The presence of these adhesive structures has been used in species descriptions. For instance, Larson (1980) included the pad-like adhesive structures on the tips of the arms as a distinguishing feature of C. corbini. However, he probably overlooked the presence of the structure in C. hawaiiensis because the character is neither well illustrated nor described in the original description by Edmondson (1930), but nevertheless present (Grohmann, Magalhães & Hirano, 1999).
The presence of individual adhesive glandular pads in the outermost secondary tentacles is widespread in Staurozoa, occurring in Craterolophus (Carlgren, 1935), Calvadosia (Uchida, 1929; Carlgren, 1935), Haliclystus (Gwilliam, 1956; Kahn et al., 2010), Depastromorpha (Carlgren, 1935), Halimocyathus (Clark, 1863), Manania (Carlgren, 1935; Larson & Fautin, 1989), and Kyopoda (Larson, 1988). It is apparently absent in Lucernaria (Carlgren, 1935), Stylocoronella (Kikinger & Salvini-Plawen, 1995), and Depastrum (Clark, 1863; Carlgren, 1935), and perhaps not even applicable in Lipkea (Pisani et al., 2007), where they are not seen in any form. Calvadosia is the only genus including species with a broad pad-like adhesive structure on the tip of each arm. This structure is apparently a synapomorphy of the clade “(((Calvadosia tasmaniensis, Calvadosia sp. 4 South Africa), Calvadosia corbini), Calvadosia sp. 3 Moorea)” (Figs. 7 and 16). The feature is also present in C. hawaiiensis and Calvadosia capensis, suggesting that they too may belong to this clade. Calvadosia cruxmelitensis has a particular adhesive pad-like structure on the tip of each arm, in which the secondary tentacles arise directly from this structure, differing from other species with pad-like adhesive structures on the tips of the arms, in which the pad is externally separated from the stem of the secondary tentacles (Corbin, 1978) (Figs. 14A–14C and 16).
Figure 16. Evolution of pad-like adhesive structures in Kishinouyeidae.

Most parsimonious reconstruction of pad-like adhesive structures in Kishinouyeidae according to our molecular phylogenetic hypothesis.
Coronal muscle
The coronal or marginal muscle is a band of epitheliomuscular cells at the calyx margin of stauromedusae (Gwilliam, 1956; Miranda, Collins & Marques, 2013). It is considered a synapomorphy of Medusozoa, probably lost in Hydrozoa (Collins et al., 2006), often associated with the swimming movement of jellyfishes (Arai, 1997). In the benthic medusae of Staurozoa, the contraction of the coronal musculature, along with contraction of the longitudinal muscles, considerably reduces the total volume of the animal, probably making its adherence to substrate more efficient in highly hydrodynamic habitats (Hyman, 1940; Miranda, Collins & Marques, 2013).
Coronal muscle can be either entire (undivided) or discontinuous (divided into perradial and interradial portions by the arms) (Figs. 11 and 12) (Clark, 1863; Carlgren, 1935; Gwilliam, 1956; Kramp, 1961). These two states have been used to differentiate genera hitherto (Table 8; Clark, 1863; Mayer, 1910; Uchida, 1929; Carlgren, 1935; Gwilliam, 1956; Kramp, 1961). In addition, the coronal muscle “appears to be vestigial or becomes ontogenetically depressed in Stylocoronella” (Kikinger & Salvini-Plawen, 1995: 908).
The position of coronal muscle in relation to the anchor/primary tentacles has also been used in the taxonomy of staurozoans (Carlgren, 1935; Gwilliam, 1956). In Manania, for example, the coronal muscle lies on the exumbrellar (external) side of the anchors (Gwilliam, 1956) (Fig. 12C), whereas in Depastromorpha the coronal muscle lies on the subumbrellar side (internal) of the anchors (Fig. 12D) (Carlgren, 1935). According to Carlgren (1935), only Manania and Depastrum have an external coronal muscle in relation to anchor/primary tentacles, but the phylogenetic signal of this character still has to be tested, specifically when specimens of D. cyathiforme become available for molecular study.
Taxonomic synopsis of staurozoa
Class Staurozoa Marques & Collins, 2004.
Order Stauromedusae Haeckel, 1879.
Suborder Amyostaurida nov.
Diagnosis: Stauromedusae without interradial longitudinal muscle in peduncle.
Family Craterolophidae Uchida, 1929
Diagnosis: No interradial longitudinal muscles in peduncle. Peduncle with four perradial chambers. Claustrum present. Without perradial and interradial anchors (rhopalioids) between arms. Individual pad-like adhesive structures can be present in outermost secondary tentacles. Coronal muscle divided.
Genus Craterolophus Clark, 1863
Type species: Craterolophus convolvulus (Johnston, 1835)
Diagnosis: Same as family.
Diversity: There are two valid species: Craterolophus convolvulus (Johnston, 1835) and Craterolophus macrocystis von Lendenfeld, 1884.
Family Kishinouyeidae Uchida, 1929
Diagnosis: No interradial longitudinal muscles in peduncle. Peduncle with one central gastric chamber and some species with four chambers at base of peduncle (pedal disk). Claustrum absent. No perradial and interradial anchors (rhopalioids) between arms (C. cruciformis with 4 interradial and 4 perradial primary tentacles, and C. tsingtaoensis with 4 perradial primary tentacles only). Species can have individual pad-like adhesive structures in outermost secondary tentacles or broad pads along tips of arms. Coronal muscle divided.
Genus Calvadosia Clark, 1863
Type species: Calvadosia campanulata (Lamouroux, 1815)
Diagnosis: Same as family.
Diversity: According to our phylogenetic and nomenclatural proposal (Figs. 3–5 and 7; Table 7), Calvadosia encompasses the species of the formerly-recognized genera Kishinouyea, Sasakiella, and Lucernariopsis. Therefore, Calvadosia has 10 species: Calvadosia campanulata (Lamouroux, 1815), Calvadosia nagatensis (Oka, 1897), Calvadosia vanhoeffeni (Browne, 1910), Calvadosia cruciformis (Okubo, 1917), Calvadosia hawaiiensis (Edmondson, 1930), Calvadosia tsingtaoensis (Ling, 1937), Calvadosia capensis (Carlgren, 1938), Calvadosia cruxmelitensis (Corbin, 1978), Calvadosia corbini (Larson, 1980), and Calvadosia tasmaniensis (Zagal et al., 2011).
Our molecular results suggest the probable existence of new species of the genus (Fig. 7; Calvadosia sp. 1 NZ, Calvadosia sp. 2 NZ, Calvadosia sp. 3 Moorea, Calvadosia sp. 4 SAF), which are being properly collected and/or morphologically analyzed in order to be tested and adequately described.
Suborder Myostaurida nov.
Diagnosis: Stauromedusae with four interradial longitudinal muscular bands in peduncle.
Family Haliclystidae Haeckel, 1879
Diagnosis: Four interradial longitudinal muscles in peduncle. Perradial and interradial anchors/primary tentacles between arms. Gonads in calyx.
Genus Depastromorpha Carlgren, 1935
Type species: Depastromorpha africana Carlgren, 1935
Diagnosis: Four interradial longitudinal muscles in peduncle. Peduncle with four perradial chambers. Claustrum present. Perradial and interradial anchors (rhopalioids) between arms. Adhesive (glandular) cushions surrounding base of eight anchors, which have knobbed remnants of primary tentacles. Individual pad-like adhesive structures in outermost secondary tentacles. Rudimentary adradial arms. Entire coronal muscle internal to anchors.
Diversity: Monospecific, Depastromorpha africana Carlgren, 1935.
The species was recently recorded for Australia and New Zealand (Grohmann, Magalhães & Hirano, 1999; Cairns et al., 2009; Zagal et al., 2011); however, the molecular results show that the specimen from Australia, Depastromorpha sp. AUS (Fig. 7), could be a new species, but more detailed analysis is needed.
Genus Depastrum Gosse, 1858
Type species: Depastrum cyathiforme (Sars, 1846)
Diagnosis: Four interradial longitudinal muscles in peduncle. Peduncle with four perradial chambers. Claustrum present. No perradial and interradial anchors (rhopalioids) between arms, but one or more primary tentacles on perradius and interradius. No pad-like adhesive structures at secondary tentacles. No discernible arms, but eight (vestigial) sinuosities. Tentacles on each of the eight adradial groups arranged in one or several rows around calyx margin. Coronal muscle entire.
Diversity: Monospecific, Depastrum cyathiforme (Sars, 1846).
Genus Haliclystus Clark, 1863
Type species: Haliclystus auricula Clark, 1863
Diagnosis: Four interradial longitudinal muscles in peduncle. Peduncle with four perradial chambers. Claustrum absent. With perradial and interradial anchors (rhopalioids) between arms. Individual pad-like adhesive structures can be present in outermost secondary tentacles. Coronal muscle divided or entire.
Diversity: According to our phylogenetic hypothesis (Figs. 3–5 and 7), the genus Stenoscyphus should be synonymized to Haliclystus. Haliclystus is the most diverse genus of Staurozoa, represented by 13 species: Haliclystus auricula Clark, 1863; Haliclystus octoradiatus Clark, 1863; Haliclystus salpinx Clark, 1863; Haliclystus inabai (Kishinouye, 1893); Haliclystus antarcticus Pfeffer, 1889; Haliclystus stejnegeri Kishinouye, 1899; Haliclystus kerguelensis Vanhöffen, 1908; Haliclystus tenuis Kishinouye, 1910; Haliclystus borealis Uchida, 1933; Haliclystus sinensis Ling, 1937; Haliclystus monstrosus (Naumov, 1961); Haliclystus californiensis Kahn et al., 2010; and Haliclystus “sanjuanensis” nomen nudum.
The molecular results show a possible new species from Australia, Haliclystus sp. AUS (Fig. 7), previously identified as Stenoscyphus inabai (McInnes, 1989; Falconer, 2013), which is being collected and morphologically analyzed in order to be properly described.
Genus Halimocyathus Clark, 1863
Type species: Halimocyathus platypus Clark, 1863
Diagnosis: Four interradial longitudinal muscles in peduncle. Peduncle with four perradial chambers. Claustrum present. Small perradial and interradial anchors between arms. Eight distinct arms, twice as long as broad. Individual pad-like adhesive structures in outermost secondary tentacles. Coronal muscle entire (?).
Diversity: Halimocyathus platypus Clark, 1863 is the only species currently valid for the genus. A second species, Halimocyathus lagena (cf. Mayer, 1910; Kramp, 1914; Kramp, 1943; Kramp, 1961), is a synonym of Manania auricula (Clark, 1863; Larson & Fautin, 1989).
Halimocyathus platypus was described based on only one specimen (Clark, 1863), and its validity and relationship with Manania spp. still has to be tested in light of molecular and morphological data whenever new material becomes available.
Genus Manania Clark, 1863
Type species: Manania auricula (Fabricius, 1780)
Diagnosis: Four interradial longitudinal muscles in peduncle. Peduncle with four perradial chambers, or one central gastric chamber, or one chamber in lower position with four chambers medially. Claustrum present. Perradial and interradial anchors between arms. Adhesive (glandular) cushions surrounding bases of eight anchors, which have knobbed remnants of primary tentacles. Eight short arms. Individual pad-like adhesive structures in outermost secondary tentacles. Entire coronal muscle, external to anchors.
Diversity: The genus Manania comprises seven valid species: Manania auricula (Fabricius, 1780); Manania hexaradiata (Broch, 1907); Manania distincta (Kishinouye, 1910); Manania atlantica (Berrill, 1962); Manania uchidai (Naumov, 1961); Manania gwilliami Larson & Fautin, 1989; and Manania handi Larson & Fautin, 1989.
Family Kyopodiidae Larson, 1988
Diagnosis: Four interradial longitudinal muscles in peduncle. Peduncle with four chambers. Claustrum absent. Body vermiform. Basal portion of peduncle enlarged, with stomach and gonads, which are absent from calyx. No evident arms. Eight adradial groups of secondary tentacles in several ranks just proximal to calyx margin. Eight primary tentacles (also called anchors, four perradial and four interradial) between groups of secondary tentacles. Individual pad-like adhesive structures present in outermost secondary tentacles. Coronal muscle entire.
Genus Kyopoda Larson, 1988
Type species: Kyopoda lamberti Larson, 1988
Diagnosis: Same as family.
Diversity: Kyopoda lamberti Larson, 1988 is the single species described for the genus.
Family Lipkeidae Vogt, 1886
Diagnosis: Four interradial longitudinal muscles in peduncle. Peduncle with one central gastric chamber. Claustrum absent. Perradial and interradial anchors absent. Eight (or more) marginal lobes (lappets). Pad-like adhesive structures absent. Coronal muscle entire.
Genus Lipkea Vogt, 1886
Type species: Lipkea ruspoliana Vogt, 1886
Diagnosis: Same as family.
Diversity: Three valid species: Lipkea ruspoliana Vogt, 1886; Lipkea sturdzii (Antipa, 1893), and Lipkea stephensoni Carlgren, 1933.
The molecular results suggest a possible new species from Japan, Lipkea sp. Japan (Fig. 7), which is being morphologically analyzed in order to be properly described. Unidentified specimens of Lipkea have also been observed in Australia and in New Zealand (Zagal et al., 2011) and the species affinities of these stauromedusae requires further studies.
Family Lucernariidae Johnston, 1847
Diagnosis: Four interradial longitudinal muscles in peduncle. Peduncle with one central gastric chamber. Claustrum absent. Perradial and interradial anchors/primary tentacles absent between arms. Pad-like adhesive structures absent.
Genus Lucernaria Müller, 1776
Type species: Lucernaria quadricornis Müller, 1776
Diagnosis: Four interradial longitudinal muscles in peduncle. Peduncle with one central gastric chamber. Claustrum absent. Perradial and interradial anchors absent between arms. Pad-like adhesive structures absent. Coronal muscle divided.
Diversity: Eight valid species: Lucernaria quadricornis Müller, 1776; Lucernaria bathyphila Haeckel, 1879; Lucernaria infundibulum Haeckel, 1879; Lucernaria haeckeli (Antipa, 1892); Lucernaria walteri (Antipa, 1892); Lucernaria australis Vanhöffen, 1908; Lucernaria sainthilairei (Redikorzev, 1925); and Lucernaria janetae Collins & Daly, 2005.
The molecular results raised possible taxonomic issues because L. bathyphila is not monophyletic (Figs. 3–5), sharing a close relationship with other deep-sea Lucernaria, L. janetae. Therefore, a detailed study of evolutionary relationships among species of Lucernaria is needed.
Genus Stylocoronella Salvini-Plawen, 1966
Type species: Stylocoronella riedli Salvini-Plawen, 1966
Diagnosis: Four interradial longitudinal muscles in peduncle. Peduncle with one central gastric chamber. Claustrum absent. Perradial and interradial anchors absent between arms. Pad-like adhesive structures absent. Coronal muscle vestigial.
Diversity: Two valid species: Stylocoronella riedli Salvini-Plawen, 1966 and Stylocoronella variabilis Salvini-Plawen, 1987.
Identification Key for the Genera of Staurozoa
-
Interradial longitudinal muscles in peduncle presentMyostaurida (2)
Interradial longitudinal muscles in peduncle absentAmyostaurida (10)
-
Body vermiform, with sac-like swelling at base of peduncle, containing stomach and gonads, which are absent from calyxKyopoda
Body not vermiform, without sac-like swelling at base of peduncle, gonads on calyx(3)
-
Primary tentacles typically absent/reduced/adradial in stauromedusa, consequently never metamorphosed into anchors(4)
Primary tentacles present (perradial and interradial) in stauromedusa, which can be metamorphosed into anchors (with or without a knobbed remnant of primary tentacle)(6)
-
Coronal muscle vestigial or absent (poorly developed)Stylocoronella
Coronal muscle present, either entire or divided(5)
-
Entire coronal muscleLipkea
Divided coronal muscleLucernaria
-
Claustrum absentHaliclystus
Claustrum present(7)
-
Long arms clearly recognizable (twice as long as broad)Halimocyathus
Short or rudimentary arms(8)
-
Pad-like adhesive structure in secondary and primary tentacles absentDepastrum
Pad-like adhesive structure at the base of anchors (modified primary tentacles) and in outermost secondary tentacles(9)
-
Coronal muscle on exumbrellar (external) side of anchorsManania
Coronal muscle on subumbrellar (internal) side of anchorsDepastromorpha
-
Claustrum absentCalvadosia
Claustrum presentCraterolophus.
Concluding Remarks
The traditional classification of Staurozoa was previously established based on subjective interpretations of anatomical similarities (Clark, 1863; Haeckel, 1879; Uchida, 1929; Uchida, 1973; Carlgren, 1935). We provide here the first classification based on a robust and comparatively complete phylogenetic analysis, including about half of the known species of Staurozoa (Figs. 3–5). Therefore, we propose a major taxonomic revision (Fig. 7; Table 7) at the suborder, family, and genus levels, in order to preserve the monophyly of taxa. Our phylogenetic analysis has also allowed for a reassessment of the evolution of the main characters used in traditional staurozoan classification. We were not able to present new data for the genera Kyopoda, Stylocoronella, Depastrum, and Halimocyathus, but provide hypotheses for their phylogenetic placements based on reported morphology (Fig. 7; Table 7). These hypotheses require new collection and detailed analysis of morphology and genetics in order to assess their validity.
Stalked jellyfishes are fascinating animals, with a peculiar anatomy related to their life cycle. Further evolutionary studies of their representatives are especially needed to gain a more complete understanding of potential homologies shared by this group and other cnidarians. In addition, such studies would support a broad spectrum of research endeavors not yet addressed for Staurozoa, such as conservation, macroecology, and biogeography.
Supplemental Information
Single most parsimonious tree, length: 163.59 steps. Bootstrap indices under parsimony at each node. ANT, Antarctica; AUS, Australia; EPR, East Pacific Rise; GER, Germany; JAP, Japan; NZ, New Zealand; SAF, South Africa; UK, the United Kingdom.
{kind=link}
Single most parsimonious tree, length: 322.30 steps. Bootstrap indices under parsimony at each node. ANT, Antarctica; AUS, Australia; EPR, East Pacific Rise; GER, Germany; JAP, Japan; NZ, New Zealand; SAF, South Africa; UK, the United Kingdom; USA, the United States of America.
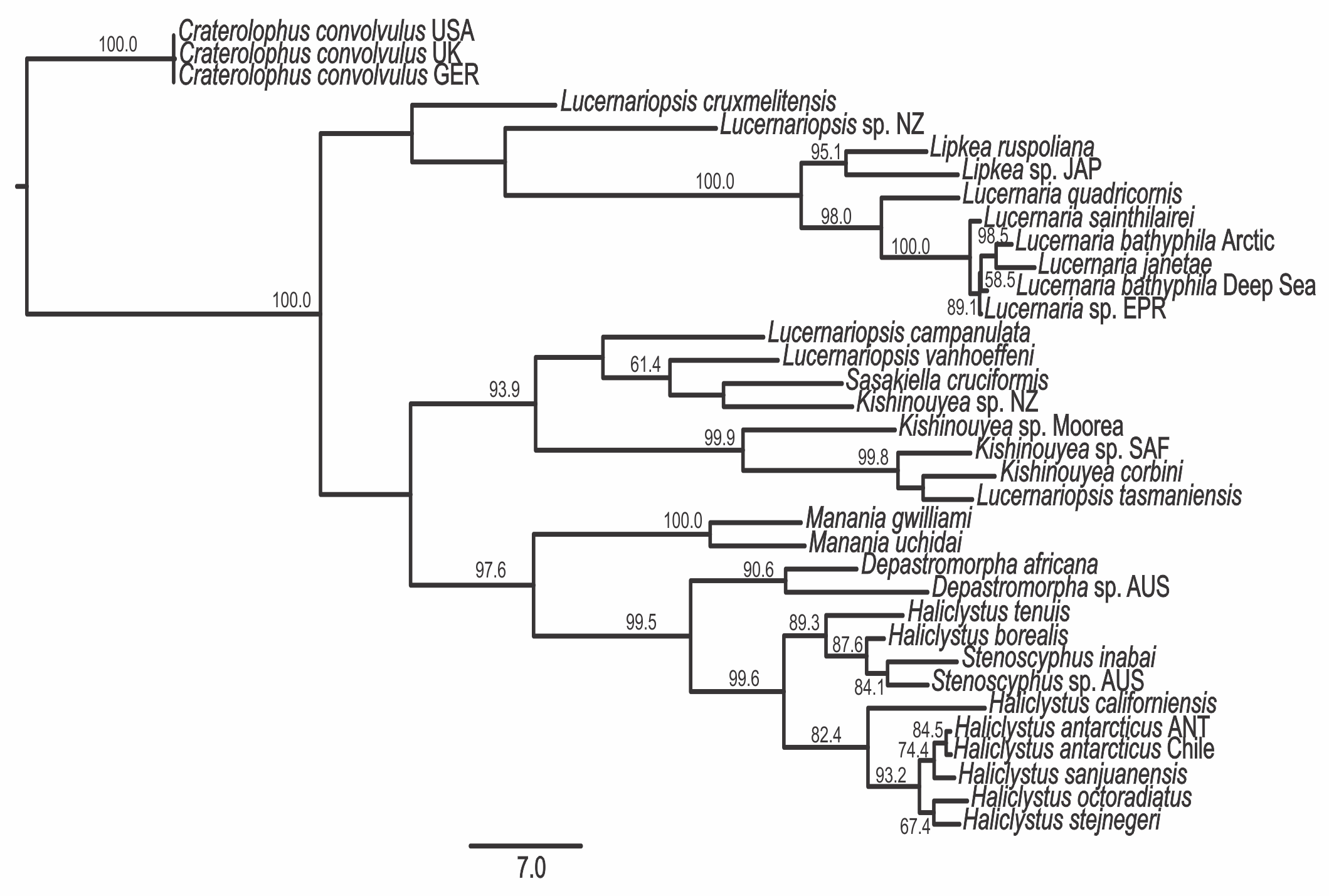{kind=link}
Consensus (50% majority-rule) of two equally most parsimonious trees, length: 122.18 steps. Bootstrap indices under parsimony at each node. ANT, Antarctica; AUS, Australia; EPR, East Pacific Rise; GER, Germany; JAP, Japan; NZ, New Zealand; SAF, South Africa; UK, the United Kingdom; USA, the United States of America.
{kind=link}
Consensus (50% majority-rule) of five equally most parsimonious trees, length: 312.48 steps. Bootstrap indices under parsimony at each node. ANT, Antarctica; AUS, Australia; EPR, East Pacific Rise; GER, Germany; JAP, Japan; NZ, New Zealand; SAF, South Africa; USA, the United States of America.
{kind=link}
Single most parsimonious tree, length: 783.04 steps. Bootstrap indices under parsimony at each node. ANT, Antarctica; AUS, Australia; EPR, East Pacific Rise; GER, Germany; JAP, Japan; NZ, New Zealand; SAF, South Africa; USA, the United States of America.
{kind=link}
Bootstrap indices under maximum likelihood at each node. ANT, Antarctica; AUS, Australia; EPR, East Pacific Rise; GER, Germany; JAP, Japan; NZ, New Zealand; SAF, South Africa; UK, the United Kingdom.
{kind=link}
Bootstrap indices under maximum likelihood at each node. ANT, Antarctica; AUS, Australia; EPR, East Pacific Rise; GER, Germany; JAP, Japan; NZ, New Zealand; SAF, South Africa; UK, the United Kingdom; USA, the United States of America.
{kind=link}
Bootstrap indices under maximum likelihood at each node. ANT, Antarctica; AUS, Australia; EPR, East Pacific Rise; GER, Germany; JAP, Japan; NZ, New Zealand; SAF, South Africa; UK, the United Kingdom; USA, the United States of America.
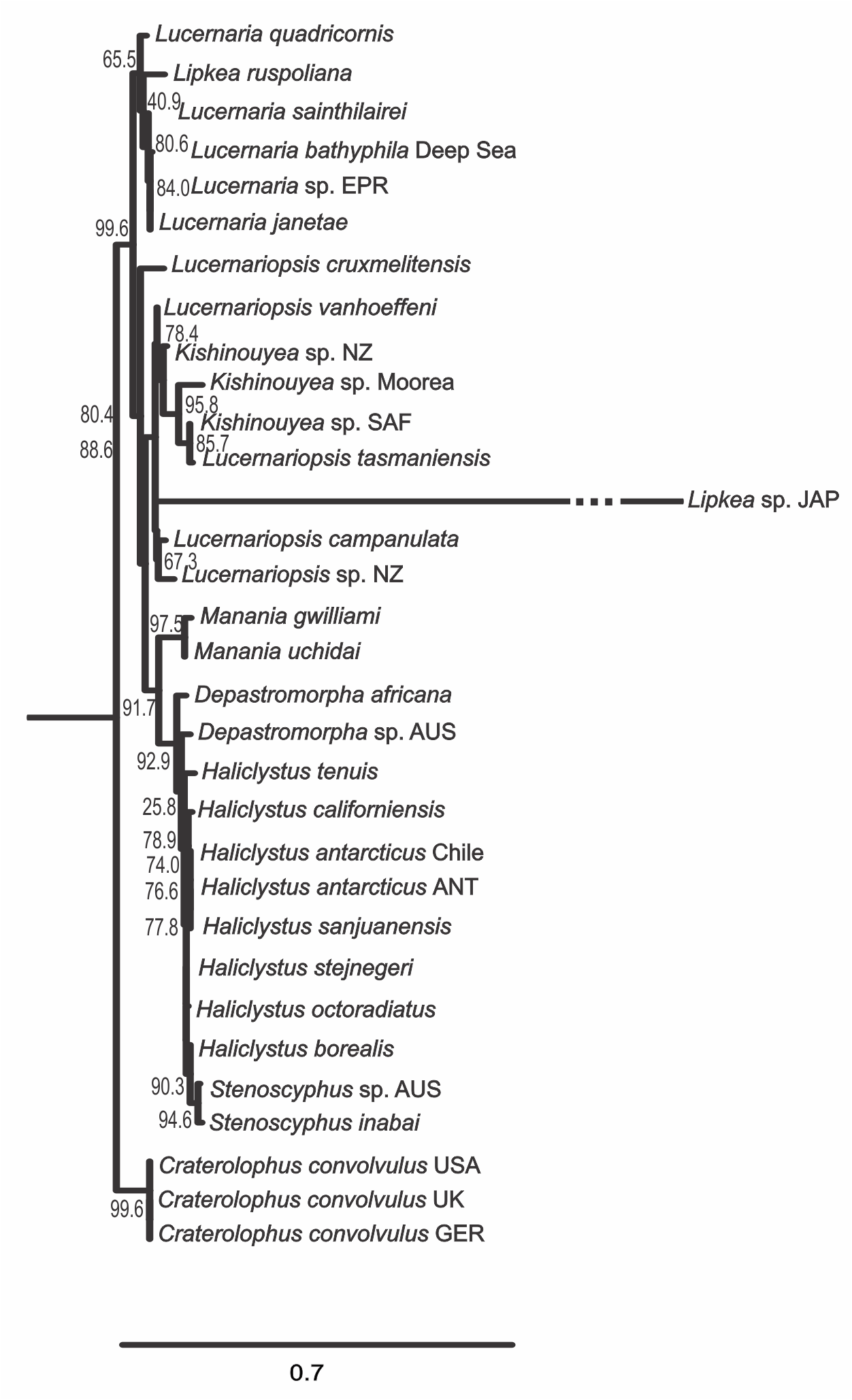{kind=link}
Bootstrap indices under maximum likelihood at each node. ANT, Antarctica; AUS, Australia; EPR, East Pacific Rise; GER, Germany; JAP, Japan; NZ, New Zealand; SAF, South Africa; USA, the United States of America.
{kind=link}
Bootstrap indices under maximum likelihood at each node. ANT, Antarctica; AUS, Australia; EPR, East Pacific Rise; GER, Germany; JAP, Japan; NZ, New Zealand; SAF, South Africa; USA, the United States of America.
{kind=link}
Posterior probability at each node. ANT, Antarctica; AUS, Australia; EPR, East Pacific Rise; GER, Germany; JAP, Japan; NZ, New Zealand; SAF, South Africa; UK, the United Kingdom.
{kind=link}
Posterior probability at each node. ANT, Antarctica; AUS, Australia; EPR, East Pacific Rise; GER, Germany; JAP, Japan; NZ, New Zealand; SAF, South Africa; UK, the United Kingdom; USA, the United States of America.
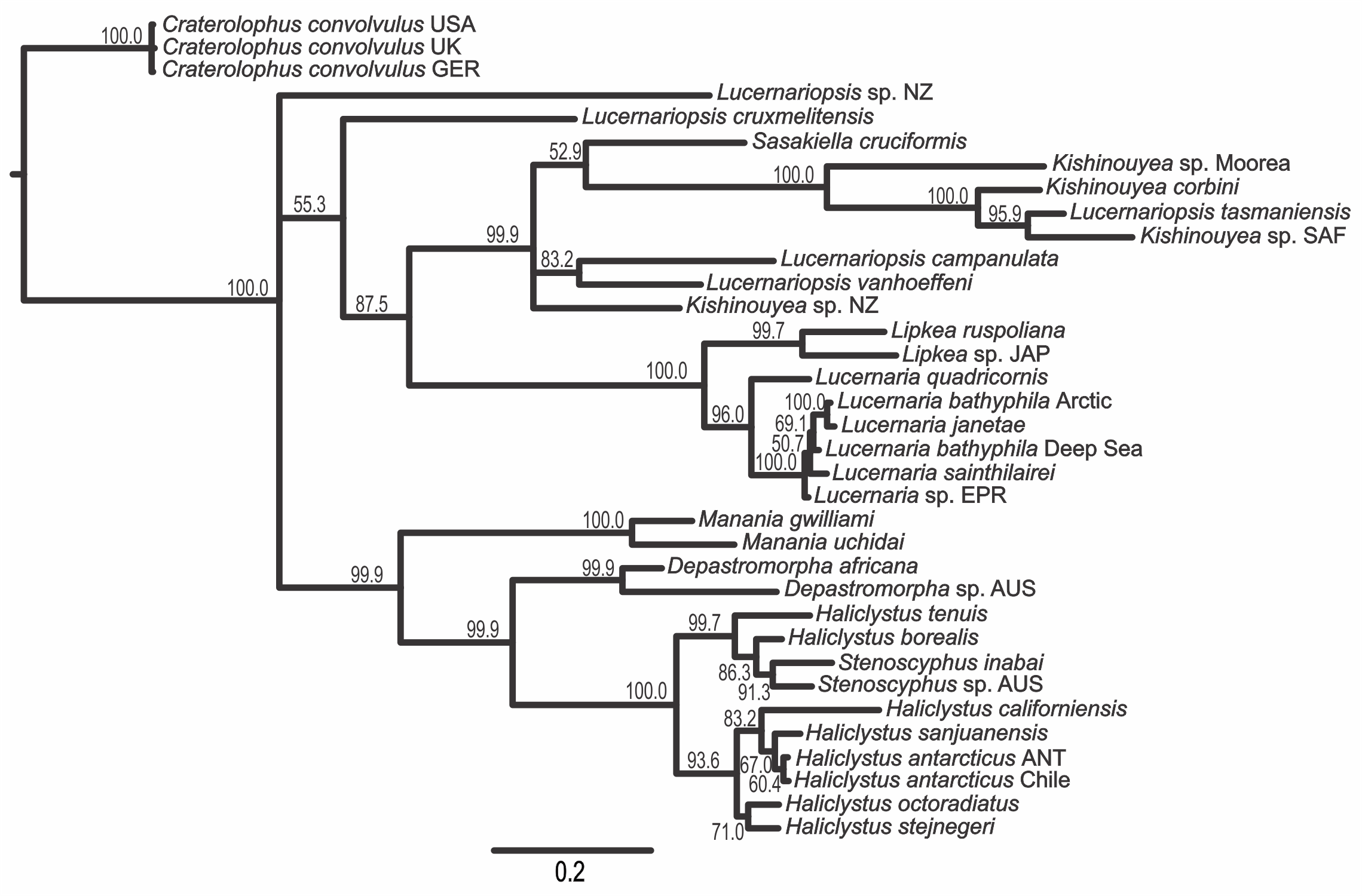{kind=link}
Posterior probability at each node. ANT, Antarctica; AUS, Australia; EPR, East Pacific Rise; GER, Germany; JAP, Japan; NZ, New Zealand; SAF, South Africa; UK, the United Kingdom; USA, the United States of America.
{kind=link}
Posterior probability at each node. ANT, Antarctica; AUS, Australia; EPR, East Pacific Rise; GER, Germany; JAP, Japan; NZ, New Zealand; SAF, South Africa; USA, the United States of America.
{kind=link}
Posterior probability at each node. ANT, Antarctica; AUS, Australia; EPR, East Pacific Rise; GER, Germany; JAP, Japan; NZ, New Zealand; SAF, South Africa; USA, the United States of America.
{kind=link}
Acknowledgments
We are grateful to Charles Griffiths and George Branch (University of Cape Town) for providing specimens from South Africa (Depastromorpha africana and Calvadosia sp. 4 South Africa); to Dennis Gordon, Kareen Schnabel, and Sadie Mills (National Institute of Water and Atmospheric Research) for providing specimens from New Zealand (Calvadosia sp. 1 New Zealand and Calvadosia sp. 2 New Zealand); to Angélique Sygut for providing specimens of Lipkea ruspoliana from the Musée océanographique de Monaco to Alexander Plotkin and Hans Tore Rapp (University of Bergen) and Daniel Jones (National Oceanography Centre of the UK) for sharing specimens of Lucernaria spp.; and to Kensuke Yanagi (Coastal Branch of Natural History Museum and Institute, Chiba) for sharing information and providing specimens from Japan (Lipkea sp. JAP). We thank Nat Evans (University of Florida) and Robert Wilson (US Geological Survey) for assisting in the generation of some genetic sequences at earlier stages of their careers. We thank Ronald Shimek, Richard Lutz (Rutgers University), Mat Vestjens, and Anne Frijsinger for kindly providing images of stalked jellyfishes (Fig. 1). The Moorea Biocode Project is acknowledged for supporting fieldwork leading to the fortuitous discovery of the first staurozoans known from French Polynesia. We are also grateful to André Morandini for discussions about Medusozoa, and to Tim Collins, Marymegan Daly, and an anonymous reviewer who helped to improve the quality of the manuscript.
Funding Statement
This study was supported by FAPESP 2010/07362-7 (LSM), 2010/52324-6 (ACM), 2011/50242-5 (ACM), 2013/50484-4 (ACM); CNPq 474672/2007-7 (ACM), 142270/2010-5 (LSM), 563106/2010-7 (ACM), 562143/2010-6 (ACM), 477156/2011-8 (ACM), 305805/2013-4 (ACM), 165066/2014-8 (LSM), 445444/2014-2 (ACM); CAPES/PDSE: 16499/12-3 (LSM); and NSF Grant AToL EF-0531779 (to P. Cartwright, AGC, and D. Fautin). The funders had no role in study design, data collection and analysis, decision to publish, or preparation of the manuscript.
Additional Information and Declarations
Competing Interests
The authors declare that they have no competing interests.
Author Contributions
Lucília S. Miranda conceived and designed the experiments, performed the experiments, analyzed the data, contributed reagents/materials/analysis tools, wrote the paper, prepared figures and/or tables, reviewed drafts of the paper.
Yayoi M. Hirano contributed reagents/materials/analysis tools, wrote the paper, reviewed drafts of the paper.
Claudia E. Mills contributed reagents/materials/analysis tools, wrote the paper, reviewed drafts of the paper.
Audrey Falconer contributed reagents/materials/analysis tools, reviewed drafts of the paper.
David Fenwick contributed reagents/materials/analysis tools, reviewed drafts of the paper.
Antonio C. Marques conceived and designed the experiments, contributed reagents/materials/analysis tools, wrote the paper, prepared figures and/or tables, reviewed drafts of the paper.
Allen G. Collins conceived and designed the experiments, performed the experiments, analyzed the data, contributed reagents/materials/analysis tools, wrote the paper, prepared figures and/or tables, reviewed drafts of the paper.
DNA Deposition
The following information was supplied regarding the deposition of DNA sequences:
GenBank Accession numbers are provided in Table 1.
Data Deposition
The following information was supplied regarding data availability:
TreeBase (TB2:S18971)
References
- Antipa (1892).Antipa G. Die lucernariden der bremer expedition nach ostspitzbergen im jahre 1889. Zoologische Jahrbücher Abtheilung Für Systematik, Geographie und Biologie der Thiere. 1892;6:377–397. [Google Scholar]
- Antipa (1893).Antipa G. Eine neue stauromeduse (Capria n. sturdzii n.) Mittheilungen aus der Zoologischen Station zu Neapel. 1893;10(4):618–632. [Google Scholar]
- Arai (1997).Arai MN. A Functional Biology of Scyphozoa. London: Chapman & Hall; 1997. [Google Scholar]
- Berrill (1962).Berrill M. The biology of three New England stauromedusae, with a description of a new species. Canadian Journal of Zoology. 1962;40(7):1262–1249. doi: 10.1139/z62-101. [DOI] [Google Scholar]
- Berrill (1963).Berrill M. Comparative functional morphology of the stauromedusae. Canadian Journal of Zoology. 1963;41(5):741–752. doi: 10.1139/z63-046. [DOI] [Google Scholar]
- Broch (1907).Broch H. Hydroiden und medusen. Report of the Second Norwegian Arctic Expedition in the “Fram” 1898–1902. 1907;2:1–12. [Google Scholar]
- Browne (1910).Browne ET. Reports of the Natural History Results of the Voyage of the SS Discovery in the Antarctic Regions in 1901, under Captain R F Scott RN. Coelentera. V. Medusae. London: London British Museum (Natural History); 1910. [Google Scholar]
- Cairns et al. (2009).Cairns SD, Gershwin L-A, Brook FJ, Pugh P, Dawson EW, Ocaña VO, Vervoort W, Williams G, Watson JE, Opresko DM, Schuchert P, Hine PM, Gordon DP, Campbell HJ, Wright AJ, Sánchez JA, Fautin DG. Phylum Cnidaria: corals, medusae, hydroids, myxozoans. In: Gordon DP, editor. New Zealand Inventory of Biodiversity. Christchurch: Canterbury University Press; 2009. pp. 59–101. vol. 1. [Google Scholar]
- Carlgren (1930).Carlgren O. Die Lucernariden. Further Zoological Results of the Swedish Antarctic Expedition 1901–1903. 1930;2:1–18. [Google Scholar]
- Carlgren (1933).Carlgren O. Zur Kenntnis der Lucernariiden Lipkea, Capria und Brochiella. Kungliga Fysiografiska Sällskapets Handlingar. 1933;44:1–19. [Google Scholar]
- Carlgren (1935).Carlgren O. Über eine neue Südafrikanische Lucernariidae, Depastromorpha africana n. gen., n. sp., nebst Bemerkungen über den Bau und die Systematik dieser Tiergruppe. Kungliga Svenska Vetenskapsakademiens Handlingar. 1935;15:1–24. [Google Scholar]
- Carlgren (1938).Carlgren O. Eine neue südafrikanische Lucernariidae, Lucernariopsis capensis. Kungliga Fysiografiska Sällskapets i Lund Förhandlingar. 1938;8:1–6. [Google Scholar]
- Cartwright et al. (2008).Cartwright P, Evans NM, Dunn CW, Marques AC, Miglietta MP, Schuchert P, Collins AC. Phylogenetics of Hydroidolina (Hydrozoa: Cnidaria) Journal of the Marine Biological Association of the United Kingdom. 2008;88(8):1163–1672. doi: 10.1017/S0025315408002257. [DOI] [Google Scholar]
- Castresana (2000).Castresana J. Selection of conserved blocks from multiple alignments for their use in phylogenetic analysis. Molecular Biology and Evolution. 2000;17(4):540–552. doi: 10.1093/oxfordjournals.molbev.a026334. [DOI] [PubMed] [Google Scholar]
- Chapman (1978).Chapman DM. Microanatomy of the cubopolyp, Tripedalia cystophora (class Cubozoa) Helgoländer wissenschaftliche Meeresuntersuchungen. 1978;31(1–2):128–168. doi: 10.1007/BF02296994. [DOI] [Google Scholar]
- Chombard et al. (1997).Chombard C, Boury-Esnault N, Tillier A, Vacelet J. Polyphyly of “Sclerosponges” (Porifera, Demospongiae) supported by 28S ribosomal sequences. Biological Bulletin. 1997;193(3):359–367. doi: 10.2307/1542938. [DOI] [PubMed] [Google Scholar]
- Clark (1863).Clark HJ. Prodromus of the history, structure, and physiology of the order Lucernariae. Journal of the Boston Society of Natural History. 1863;7:531–567. [Google Scholar]
- Clark (1878).Clark HJ. Lucernariae and their allies. A memoir on the anatomy and physiology of Haliclystus auricula and other Lucernarians, with a discussion of their relations to other Acalephae; to beroids, and polypi. Smithsonian Contributions to Knowledge. 1878;242:1–130. [Google Scholar]
- Collins & Daly (2005).Collins AG, Daly M. A new deepwater species of Stauromedusae, Lucernaria janetae (Cnidaria, Staurozoa, Lucernariidae), and a preliminary investigation of stauromedusan phylogeny based on nuclear and mitochondrial rDNA data. Biological Bulletin. 2005;208(3):221–230. doi: 10.2307/3593154. [DOI] [PubMed] [Google Scholar]
- Collins et al. (2006).Collins AG, Schuchert P, Marques AC, Jankowski T, Medina M, Schierwater B. Medusozoan phylogeny and character evolution clarified by large and small subunit rDNA data and an assessment of the utility of phylogenetic mixture models. Systematic Biology. 2006;55(1):97–115. doi: 10.1080/10635150500433615. [DOI] [PubMed] [Google Scholar]
- Collins et al. (2008).Collins AG, Bentlage B, Lindner A, Lindsay D, Haddock SHD, Jarms G, Norenburg JL, Jankowski T, Cartwright P. Phylogenetics of Trachylina (Cnidaria: Hydrozoa) with new insights on the evolution of some problematical taxa. Journal of the Marine Biological Association of the United Kingdom. 2008;88(8):1673–1685. doi: 10.1017/S0025315408001732. [DOI] [Google Scholar]
- Corbin (1978).Corbin PG. A new species of the stauromedusan genus Lucernariopsis (Coelenterata: Scyphomedusae) Journal of the Marine Biological Association of the United Kingdom. 1978;58(2):285–290. doi: 10.1017/S0025315400027983. [DOI] [Google Scholar]
- Cunha, Genzano & Marques (2015).Cunha AF, Genzano GN, Marques AC. Reassessment of morphological diagnostic characters and species boundaries requires taxonomical changes for the genus Orthopyxis L. Agassiz, 1862 (Campanulariidae, Hydrozoa) and some related campanulariids. PLoS ONE. 2015;10(2):e1951. doi: 10.1371/journal.pone.0117553. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Cunningham & Buss (1993).Cunningham CW, Buss LW. Molecular evidence for multiple episodes of paedomorphosis in the family Hydractiniidae. Biochemical Systematics and Ecology. 1993;21(1):57–69. doi: 10.1016/0305-1978(93)90009-G. [DOI] [Google Scholar]
- Cuvier (1817).Cuvier G. Le Règne animal distribué d’après son organisation, pour servir de base è l’histoire naturelle des animaux et d’introduction è l’anatomie comparée. Paris: Chez Déterville Libraire; 1817. Tome IV. [Google Scholar]
- Cuvier (1830).Cuvier G. Le Règne animal distribué d’après son organisation, pour servir de base è l’histoire naturelle des animaux et d’introduction è l’anatomie comparée. Paris: Chez Déterville Libraire; 1830. Tome III. [Google Scholar]
- Daly et al. (2007).Daly M, Brugler MR, Cartwright P, Collins AG, Dawson MN, Fautin DG, France SC, McFadden CS, Opresko DM, Rodriguez E, Romano SL, Stake JL. The phylum Cnidaria: a review of phylogenetic patterns and diversity 300 years after Linnaeus. Zootaxa. 2007;1668:127–182. [Google Scholar]
- Darriba et al. (2012).Darriba D, Taboada GL, Doallo R, Posada D. jModelTest 2: more models, new heuristics and parallel computing. Nature Methods. 2012;9(8):772. doi: 10.1038/nmeth.2109. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Dawson (2004).Dawson MN. Some implications of molecular phylogenetics for understanding biodiversity in jellyfishes, with emphasis on Scyphozoa. Hydrobiologia. 2004;530/531(1–3):249–260. doi: 10.1007/s10750-004-2659-3. [DOI] [Google Scholar]
- Dawson & Jacobs (2001).Dawson MN, Jacobs DK. Molecular evidence for cryptic species of Aurelia aurita (Cnidaria, Scyphozoa) Biological Bulletin. 2001;200(1):92–96. doi: 10.2307/1543089. [DOI] [PubMed] [Google Scholar]
- den Hartog (1976).den Hartog JC. A record of the stauromedusa Depastrum cyathiforme (M. Sars, 1846) in France. Travaux de la Station Biologique de Roscoff. 1976;23:3–4. [Google Scholar]
- Edmondson (1930).Edmondson CH. New Hawaiian medusae. Bernice P. Bishop Museum Occasional Papers. 1930;9:1–16. [Google Scholar]
- Elmhirst (1922).Elmhirst R. Notes on Lucernaria quadricornis, Müller, and related species. The Annals and Magazine of Natural History. 1922;10(56):221–224. doi: 10.1080/00222932308632766. [DOI] [Google Scholar]
- Evans et al. (2008).Evans NM, Lindner A, Raikova EV, Collins AG, Cartwright P. Phylogenetic placement of the enigmatic parasite, Polypodium hydriforme, within the phylum Cnidaria. BMC Evolutionary Biology. 2008;8(1):139. doi: 10.1186/1471-2148-8-139. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Fabricius (1780).Fabricius O. Fauna Groenlandica: systematice sistens animalia Groenlandiae occidentalis hactenus indagata, quoad nomen specificum, triviale, vernaculumque; synonyma auctorum plurium, descriptionem, locum, victum, generationem, mores, usum, capturamque singuli, prout detegendi occasio fuit, maximaque parte secundum proprias observations. Hafniae et Lipsiae: Gottlob Rothe; 1780. [Google Scholar]
- Falconer (2013).Falconer A. A stalked jellyfish Stenoscyphus inabai (Kishinouye, 1893) (Stauromedusae), found at the jawbone, port phillip bay, Victoria. Victoria Naturalist. 2013;130(5):202–207. [Google Scholar]
- Felsenstein (1985).Felsenstein J. Confidence limits on phylogenies: an approach using the bootstrap. Evolution. 1985;39(4):783–791. doi: 10.2307/2408678. [DOI] [PubMed] [Google Scholar]
- Franc (1994).Franc A. Classe des Scyphozoaires. In: Grassé PP, editor. Traité de Zoologie, Cnidaires, Ctenaires. Paris: Masson; 1994. pp. 597–1117. Tome III, Fascicule 2. [Google Scholar]
- Geller et al. (2013).Geller J, Meyer C, Parker M, Hawk H. Redesign of PCR primers for mitochondrial cytochrome c oxidase subunit I for marine invertebrates and application in all-taxa biotic surveys. Molecular Ecology Resources. 2013;13(5):851–861. doi: 10.1111/1755-0998.12138. [DOI] [PubMed] [Google Scholar]
- Goette (1887).Goette A. Abhandlungen zur Entwickelungsgeschichte der Tiere. Entwickelungsgeschichte der Aurelia aurita und Cotylorhiza tuberculata. Hamburg und Leipzig: Verlag von Leopold Voss; 1887. [Google Scholar]
- Goloboff, Farris & Nixon (2008).Goloboff P, Farris J, Nixon K. TNT, a free program for phylogenetic analysis. Cladistics. 2008;24(5):774–786. doi: 10.1111/cla.2008.24.issue-5. [DOI] [Google Scholar]
- Gosse (1858).Gosse PH. Synopsis of the families, genera, and species of the British Actiniae. The Annals and Magazine of Natural History. 1858;3:414–419. [Google Scholar]
- Gosse (1860).Gosse PH. On the Lucernaria cyathiformis of Sars. The Annals and Magazine of Natural History. 1860;5(30):480–481. doi: 10.1080/00222936008697258. [DOI] [Google Scholar]
- Grohmann, Magalhães & Hirano (1999).Grohmann PA, Magalhães MP, Hirano YM. First record of the order Stauromedusae (Cnidaria, Scyphozoa) from the tropical southwestern Atlantic, with a review of the distribution of Stauromedusae in the southern hemisphere. Species Diversity. 1999;4:381–388. [Google Scholar]
- Gross (1900).Gross I. Zur Anatomie der Lucernariden. Jenaischen Zeitschrift für Naturwissenschaft. 1900;33:611–624. [Google Scholar]
- Guindon et al. (2010).Guindon S, Dufayard JF, Lefort V, Anisimova M, Hordijk W, Gascuel O. New algorithms and methods to estimate maximum-likelihood phylogenies: assessing the performance of PhyML 3.0. Systematic Biology. 2010;59(3):307–321. doi: 10.1093/sysbio/syq010. [DOI] [PubMed] [Google Scholar]
- Gwilliam (1956).Gwilliam GF. Berkeley: University of California; 1956. Studies on West Coast Stauromedusae. Ph.D. Dissertation. [Google Scholar]
- Haeckel (1879).Haeckel E. Das system der medusen. I, 2: System der Acraspeden. Zweite Hälfte des Systems der Medusen. Jena: Gustav Fischer; 1879. [Google Scholar]
- Hirano (1986).Hirano YM. Species of Stauromedusae from Hokkaido, with notes on their metamorphosis. Journal of the Faculty of Science, Hokkaido University, Zoology. 1986;24:182–201. [Google Scholar]
- Hirano (1997).Hirano YM. A review of a supposedly circumboreal species of stauromedusa, Haliclystus auricula (Rathke, 1806). In: den Hartog JC, editor. Proceedings of the 6th International Conference on Coelenterate Biology; Leiden: National Naturhistorisch Museum, Noordwijkerhout; 1997. pp. 247–252. [Google Scholar]
- Humanson (1962).Humanson GL. Animal Tissue Techniques. San Francisco: W. H. Freeman and Company; 1962. [Google Scholar]
- Hutton (1880).Hutton FW. Contributions to the Coelenterate fauna of New Zealand. Transactions and Proceedings of the New Zealand Institute. 1880;12:274–276. [Google Scholar]
- Hyman (1940).Hyman LH. Observations and experiments on the physiology of medusae. Biological Bulletin. 1940;79(2):282–296. doi: 10.2307/1537823. [DOI] [Google Scholar]
- Ji, Zhang & He (2003).Ji Y-J, Zhang D-X, He L-J. Evolutionary conservation and versatility of a new set of primers for amplifying the ribosomal internal transcribed spacer regions in insects and other invertebrates. Molecular Ecology Notes. 2003;3(4):581–585. doi: 10.1046/j.1471-8286.2003.00519.x. [DOI] [Google Scholar]
- Johnston (1835).Johnston G. Illustrations in British Zoology. The Magazine of Natural History, and Journal of Zoology, Botany, Mineralogy, Geology, and Meteorology. 1835;8:59–61. [Google Scholar]
- Johnston (1847).Johnston G. A History of the British Zoophytes. Second Edition. London: John Van Voorst, Paternoster Row; 1847. vol. I. [Google Scholar]
- Kahn et al. (2010).Kahn AS, Matsumoto GI, Hirano YM, Collins AG. Haliclystus californiensis, a “new” species of stauromedusa (Cnidaria: Staurozoa) from the northeast Pacific, with a key to the species of Haliclystus. Zootaxa. 2010;2518:49–59. [Google Scholar]
- Katoh & Standley (2013).Katoh K, Standley DM. MAFFT multiple sequence alignment software version 7: improvements in performance and usability. Molecular Biology and Evolution. 2013;30(4):772–780. doi: 10.1093/molbev/mst010. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Kayal et al. (2013).Kayal E, Roure B, Phillippe H, Collins AG, Lavrov DV. Cnidarian phylogenetic relationships as revealed by mitogenomics. BMC Evolutionary Biology. 2013;13(1):5. doi: 10.1186/1471-2148-13-5. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Kikinger & Salvini-Plawen (1995).Kikinger R, Salvini-Plawen LV. Development from polyp to stauromedusa in Stylocoronella (Cnidaria: Scyphozoa) Journal of the Marine Biological Association of the United Kingdom. 1995;75(4):899–912. doi: 10.1017/S0025315400038236. [DOI] [Google Scholar]
- Kishinouye (1893).Kishinouye K. Mushi-kurage, Depastrum inabai n.sp. Zoological Magazine. 1893;5:416–419. [Google Scholar]
- Kishinouye (1899).Kishinouye K. Contributions to the natural history of the Commander Islands. XIII. A new species of stalked medusae, Haliclystus stejnegeri. Proceedings of the United States National Museum. 1899;22(1188):125–129. doi: 10.5479/si.00963801.22-1188.125. [DOI] [Google Scholar]
- Kishinouye (1902).Kishinouye K. Some new Scyphomedusae of Japan. Journal of the College of Science, Imperial University of Tokyo. 1902;17:1–17. [Google Scholar]
- Kishinouye (1910).Kishinouye K. Some medusae of Japanese water. Journal of the College of Science, Imperial University of Tokyo. 1910;27:1–35. [Google Scholar]
- Kramp (1914).Kramp PL. Meduser og Siphonophorer. Catalogue of the Medusae and Siphonophora of Greenland. Meddelelser om Grønland. 1914;23:381–456. [Google Scholar]
- Kramp (1943).Kramp PL. The Zoology of East Greenland. Medusae, Siphonophora and Ctenophora. Meddelelser om Grønland. 1943;121:1–20. [Google Scholar]
- Kramp (1961).Kramp PL. Synopsis of the medusae of the world. Journal of the Marine Biological Association of the United Kingdom. 1961;40(1):292–303. doi: 10.1017/S0025315400007438. [DOI] [Google Scholar]
- Lamouroux (1815).Lamouroux JVF. Mémoire sur la Lucernaire campanulée. Mémoirs du Muséum d’ Histoire Naturelle. 1815;II:460–473. [Google Scholar]
- Larson (1980).Larson RJ. A new stauromedusa, Kishinouyea corbini (Scyphozoa, Stauromedusae) from the tropical western Atlantic. Bulletin of Marine Science. 1980;30(1):102–107. [Google Scholar]
- Larson (1988).Larson RJ. Kyopoda lamberti gen.nov., sp.nov., an atypical stauromedusa (Scyphozoa, Cnidaria) from the eastern Pacific, representing a new family. Canadian Journal of Zoology. 1988;66(10):2301–2303. doi: 10.1139/z88-341. [DOI] [Google Scholar]
- Larson & Fautin (1989).Larson RJ, Fautin DG. Stauromedusae of the genus Manania (=Thaumatoscyphus) (Cnidaria, Scyphozoa) in the northeast Pacific, including descriptions of new species Manania gwilliami and Manania handi. Canadian Journal of Zoology. 1989;67(6):1543–1549. doi: 10.1139/z89-219. [DOI] [Google Scholar]
- Ling (1937).Ling SW. Studies on Chinese Stauromedusae. I. Stauromedusae from Tsingtao. Amoy Marine Biological Bulletin. 1937;3:1–35. [Google Scholar]
- Ling (1939).Ling SW. Studies on Chinese stauromedusae. II. Further studies on some stauromedusae from China. Lingnan Science Journal. 1939;18(3):4a95–503. [Google Scholar]
- Lutz et al. (2006).Lutz RA, Collins AG, Annis ER, Reed AJ, Bennett KF, Halanych KM, Vrijenhoek RC. Stauromedusan populations inhabiting deep-sea hydrothermal vents along the southern East Pacific Rise. Cahiers de Biologie Marine. 2006;47(4):409–413. [Google Scholar]
- Mahoney (1966).Mahoney R. Laboratory Techniques in Zoology. London: Butterworth & Co; 1966. [Google Scholar]
- Marques & Collins (2004).Marques AC, Collins AG. Cladistic analysis of Medusozoa and cnidarian evolution. Invertebrate Biology. 2004;123(1):32–42. doi: 10.1111/j.1744-7410.2004.tb00139.x. [DOI] [Google Scholar]
- Mayer (1910).Mayer AG. Medusae of the World. Washington, D.C.: Carnegie Institution Publishing; 1910. Scyphomedusae. vol. III. Publication 109. [Google Scholar]
- McInnes (1989).McInnes DE. A stalked jellyfish (Stauromedusae) found at Black Rock, Port Phillip Bay. A first recording in Australia. Victorian Naturalist. 1989;106(3):86–92. [Google Scholar]
- Medina et al. (2001).Medina M, Collins AG, Silberman JD, Sogin ML. Evaluating hypotheses of basal animal phylogeny using complete sequences of large and small subunit rRNA. Proceedings of the National Academy of Sciences of the United States of America. 2001;98(17):9707–9712. doi: 10.1073/pnas.171316998. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Miranda, Morandini & Marques (2009).Miranda LS, Morandini AC, Marques AC. Taxonomic review of Haliclystus antarcticus Pfeffer, 1889 (Stauromedusae, Staurozoa, Cnidaria), with remarks on the genus Haliclystus Clark, 1863. Polar Biology. 2009;32(10):1507–1519. doi: 10.1007/s00300-009-0648-8. [DOI] [Google Scholar]
- Miranda, Collins & Marques (2010).Miranda LS, Collins AG, Marques AC. Molecules clarify a cnidarian life cycle–the “hydrozoan” Microhydrula limopsicola is an early life stage of the staurozoan Haliclystus antarcticus. PLoS ONE. 2010;5(4):e1951. doi: 10.1371/journal.pone.0010182. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Miranda et al. (2012).Miranda LS, Haddad MA, Mills CE, Marques AC. Lucernariopsis capensis Carlgren, 1938 (Cnidaria, Staurozoa) in Brazil: first record outside its type locality in South Africa. Zootaxa. 2012;3158:60–64. [Google Scholar]
- Miranda, Collins & Marques (2013).Miranda LS, Collins AG, Marques AC. Internal anatomy of Haliclystus antarcticus (Cnidaria, Staurozoa) with a discussion on histological features used in staurozoan taxonomy. Journal of Morphology. 2013;274(12):1365–1383. doi: 10.1002/jmor.v274.12. [DOI] [PubMed] [Google Scholar]
- Müller (1776).Müller OF. Zoologiae Danicae Prodromus, seu animalium Daniae et Norvegiae indigenarum, Characteres, Nomina et Synonyma imprimis popularium. Havniae: Typis Hallageriis; 1776. [DOI] [Google Scholar]
- Naumov (1961).Naumov DV. Stsifoidnye meduzy morei SSSR (Scyphozoan medusae in the sea of USSR) Opredeliteli po Faune SSSR. 1961;75:1–98. [Google Scholar]
- Nawrocki et al. (2013).Nawrocki AM, Collins AG, Hirano YM, Schuchert P, Cartwright P. Phylogenetic placement of Hydra and relationships within Aplanulata (Cnidaria: Hydrozoa) Molecular Phylogenetics and Evolution. 2013;67(1):60–71. doi: 10.1016/j.ympev.2012.12.016. [DOI] [PubMed] [Google Scholar]
- Oka (1897).Oka A. Sur une nouvelle espèce Japonaise du genre Lucernaria. Annotationes Zoologicae Japonenses. 1897;1:141–145. [Google Scholar]
- Okubo (1917).Okubo T. Preliminary note on a new genus of Stauromedusae from Hokkaido. Zoological Magazine. 1917;29:317–322. [Google Scholar]
- Pfeffer (1889).Pfeffer G. Zur Fauna von Süd-Georgien. Jahrbuch der Hamburgischen Wissenschaftlichen Anstalten. 1889;6:37–55. [Google Scholar]
- Pisani et al. (2007).Pisani V, Otero-Ferrer F, Lotto S, Maurel P, Goy J. Lipkea ruspoliana Vogt, 1887 (Stauromedusae, Scyphozoa, Cnidaria) dans les aquariums du Musée Océanographique de Monaco. Bulletin de la Société Zoologique de France. 2007;132:183–190. [Google Scholar]
- Redikorzev (1925).Redikorzev V. Die Lucernose des Weißen Meeres. Zoologischer Anzeiger. 1925;62:155–157. [Google Scholar]
- Redmond et al. (2007).Redmond NE, van Soest RWM, Kelly M, Raleigh J, Travers SAA, McCormack GP. Reassessment of the classification of the Order Haplosclerida (Class Demospongiae, Phylum Porifera) using 18S rRNA gene sequence data. Molecular Phylogenetics and Evolution. 2007;43(1):344–352. doi: 10.1016/j.ympev.2006.10.021. [DOI] [PubMed] [Google Scholar]
- Ronquist & Huelsenbeck (2003).Ronquist F, Huelsenbeck JP. MRBAYES 3: Bayesian phylogenetic inference under mixed models. Bioinformatics. 2003;19(12):1572–1574. doi: 10.1093/bioinformatics/btg180. [DOI] [PubMed] [Google Scholar]
- Salvini-Plawen (1966).Salvini-Plawen LV. Zur Kenntnis der Cnidaria des nordadriatischen Mesopsammon. Veröffentlichungen des Instituts für Meeresforschung in Bremerhaven. 1966;2:165–186. [Google Scholar]
- Salvini-Plawen (1987).Salvini-Plawen LV. Mesopsammic Cnidaria from Plymouth (with systematic notes) Journal of the Marine Biological Association of the United Kingdom. 1987;67(3):623–637. doi: 10.1017/S0025315400027338. [DOI] [Google Scholar]
- Sars (1846).Sars M. Fauna littoralis Norvegiae oder Beschreibung und Abbildungen neuer oder wenig bekannter Seethiere, nebst Beobachtungen über die Organisation, Lebensweise u. Entwickelung derselben. Christiania: Druck und Verlag von Johann Dahk; 1846. [Google Scholar]
- Swofford (2002).Swofford DL. Version 4. Sunderland: Sinauer Associates; 2002. PAUP*. Phylogenetic Analysis Using Parsimony (*and Other Methods) Program and Documentation. [Google Scholar]
- Talavera & Castresana (2007).Talavera G, Castresana J. Improvement of phylogenies after removing divergent and ambiguously aligned blocks from protein sequence alignments. Systematic Biology. 2007;56(4):564–577. doi: 10.1080/10635150701472164. [DOI] [PubMed] [Google Scholar]
- Thiel (1966).Thiel H. The evolution of Scyphozoa: a review. In: Rees WJ, editor. The Cnidaria and their Evolution. London: Academic Press; 1966. pp. 77–118. [Google Scholar]
- Uchida (1929).Uchida T. Studies on the Stauromedusae and Cubomedusae, with special reference to their metamorphosis. Japanese Journal of Zoology. 1929;2:103–193. [Google Scholar]
- Uchida (1933).Uchida T. Eine neue Becherqualle aus Hokkaido. Proceedings of the Imperial Academy of Japan. 1933;9:450–452. [Google Scholar]
- Uchida (1973).Uchida T. The systematic position of the Stauromedusae. Publications of the Seto Marine Biological Laboratory. 1973;20:133–139. [Google Scholar]
- Uchida & Hanaoka (1933).Uchida T, Hanaoka K-I. On the morphology of a stalked medusa, Thaumatoscyphus distinctus Kishinouye. Journal of the Faculty of Science, Hokkaido Imperial University, Series VI, Zoology. 1933;2(3):135–153. [Google Scholar]
- Uchida & Hanaoka (1934).Uchida T, Hanaoka K-I. Anatomy of two stalked medusae with remarks on the distribution of the Stauromedusae in Japan. Journal of the Faculty of Science, Hokkaido Imperial University, Series VI, Zoology. 1934;2:209–219. [Google Scholar]
- van Iten et al. (2006).van Iten H, Leme JM, Simões MG, Marques AC, Collins AG. Reassessment of the phylogenetic position of Conulariids (?Ediacaran-Triassic) within the subphylum Medusozoa (phylum Cnidaria) Journal of Systematic Palaeontology. 2006;4(2):109–118. doi: 10.1017/S1477201905001793. [DOI] [Google Scholar]
- van Iten et al. (2014).van Iten H, Marques AC, Leme JM, Pacheco MLAF, Simões MG. Origin and early diversification of the phylum Cnidaria Verrill: major developments in the analysis of the taxon’s Proterozoic-Cambrian history. Palaeontology. 2014;57(4):677–690. doi: 10.1111/pala.2014.57.issue-4. [DOI] [Google Scholar]
- Vanhöffen (1908).Vanhöffen E. Die Lucernariden und Scyphomedusen der Deutschen Südpolar-Expedition 1901–1903. Deutsche Südpolar-Expedition. 1908;10:25–49. [Google Scholar]
- Vogt (1886).Vogt C. Sur une médusaire sessile, Lipkea ruspoliana. Archives des Sciences. 1886;16:356–362. [Google Scholar]
- Voigt et al. (2004).Voigt O, Collins AG, Pearse VB, Pearse JS, Ender A, Hadrys H, Schierwater B. Placozoa–no longer a phylum of one. Current Biology. 2004;14(22):R944–R945. doi: 10.1016/j.cub.2004.10.036. [DOI] [PubMed] [Google Scholar]
- von Lendenfeld (1884).von Lendenfeld R. The Schyphomedusae of the southern hemisphere. Proceedings of the Linnean Society of New South Wales. 1884;9:155–169. [Google Scholar]
- Wietrzykowski (1911).Wietrzykowski W. Seconde note sur le développement des Lucernaires. Archives de Zoologie Expérimentale et Générale, 5th Series. 1911;6:49–52. [Google Scholar]
- Wietrzykowski (1912).Wietrzykowski W. Recherches sur le développement des Lucernaires. Archives de Zoologie Expérimentale et Générale, 5th Series. 1912;10:1–95. [Google Scholar]
- Zagal et al. (2011).Zagal CJ, Hirano YM, Mills CE, Edgar GJ, Barrett NS. New records of Staurozoa from Australian coastal waters, with a description of a new species of Lucernariopsis Uchida, 1929 (Cnidaria, Staurozoa, Stauromedusae) and a key to Australian Stauromedusae. Marine Biology Research. 2011;7(7):651–666. doi: 10.1080/17451000.2011.558097. [DOI] [Google Scholar]
- Zapata et al. (2015).Zapata F, Goetz FE, Smith AS, Howison M, Siebert S, Church S, Sanders SM, Ames CL, McFadden CS, France SC, Daly M, Collins AG, Haddock SHD, Dunn C, Cartwright P. Phylogenomic analyses support traditional relationships within Cnidaria. PLoS ONE. 2015;10(10):e1951. doi: 10.1371/journal.pone.0139068. [DOI] [PMC free article] [PubMed] [Google Scholar]
Associated Data
This section collects any data citations, data availability statements, or supplementary materials included in this article.
Supplementary Materials
Single most parsimonious tree, length: 163.59 steps. Bootstrap indices under parsimony at each node. ANT, Antarctica; AUS, Australia; EPR, East Pacific Rise; GER, Germany; JAP, Japan; NZ, New Zealand; SAF, South Africa; UK, the United Kingdom.
Single most parsimonious tree, length: 322.30 steps. Bootstrap indices under parsimony at each node. ANT, Antarctica; AUS, Australia; EPR, East Pacific Rise; GER, Germany; JAP, Japan; NZ, New Zealand; SAF, South Africa; UK, the United Kingdom; USA, the United States of America.
Consensus (50% majority-rule) of two equally most parsimonious trees, length: 122.18 steps. Bootstrap indices under parsimony at each node. ANT, Antarctica; AUS, Australia; EPR, East Pacific Rise; GER, Germany; JAP, Japan; NZ, New Zealand; SAF, South Africa; UK, the United Kingdom; USA, the United States of America.
Consensus (50% majority-rule) of five equally most parsimonious trees, length: 312.48 steps. Bootstrap indices under parsimony at each node. ANT, Antarctica; AUS, Australia; EPR, East Pacific Rise; GER, Germany; JAP, Japan; NZ, New Zealand; SAF, South Africa; USA, the United States of America.
Single most parsimonious tree, length: 783.04 steps. Bootstrap indices under parsimony at each node. ANT, Antarctica; AUS, Australia; EPR, East Pacific Rise; GER, Germany; JAP, Japan; NZ, New Zealand; SAF, South Africa; USA, the United States of America.
Bootstrap indices under maximum likelihood at each node. ANT, Antarctica; AUS, Australia; EPR, East Pacific Rise; GER, Germany; JAP, Japan; NZ, New Zealand; SAF, South Africa; UK, the United Kingdom.
Bootstrap indices under maximum likelihood at each node. ANT, Antarctica; AUS, Australia; EPR, East Pacific Rise; GER, Germany; JAP, Japan; NZ, New Zealand; SAF, South Africa; UK, the United Kingdom; USA, the United States of America.
Bootstrap indices under maximum likelihood at each node. ANT, Antarctica; AUS, Australia; EPR, East Pacific Rise; GER, Germany; JAP, Japan; NZ, New Zealand; SAF, South Africa; UK, the United Kingdom; USA, the United States of America.
Bootstrap indices under maximum likelihood at each node. ANT, Antarctica; AUS, Australia; EPR, East Pacific Rise; GER, Germany; JAP, Japan; NZ, New Zealand; SAF, South Africa; USA, the United States of America.
Bootstrap indices under maximum likelihood at each node. ANT, Antarctica; AUS, Australia; EPR, East Pacific Rise; GER, Germany; JAP, Japan; NZ, New Zealand; SAF, South Africa; USA, the United States of America.
Posterior probability at each node. ANT, Antarctica; AUS, Australia; EPR, East Pacific Rise; GER, Germany; JAP, Japan; NZ, New Zealand; SAF, South Africa; UK, the United Kingdom.
Posterior probability at each node. ANT, Antarctica; AUS, Australia; EPR, East Pacific Rise; GER, Germany; JAP, Japan; NZ, New Zealand; SAF, South Africa; UK, the United Kingdom; USA, the United States of America.
Posterior probability at each node. ANT, Antarctica; AUS, Australia; EPR, East Pacific Rise; GER, Germany; JAP, Japan; NZ, New Zealand; SAF, South Africa; UK, the United Kingdom; USA, the United States of America.
Posterior probability at each node. ANT, Antarctica; AUS, Australia; EPR, East Pacific Rise; GER, Germany; JAP, Japan; NZ, New Zealand; SAF, South Africa; USA, the United States of America.
Posterior probability at each node. ANT, Antarctica; AUS, Australia; EPR, East Pacific Rise; GER, Germany; JAP, Japan; NZ, New Zealand; SAF, South Africa; USA, the United States of America.