
Figure 1.
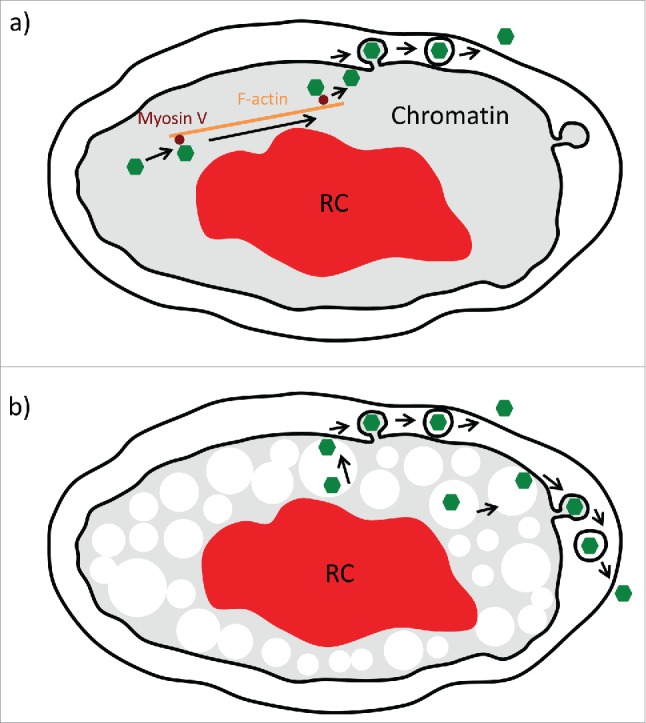
Models of nuclear capsid motility. a) Previous work suggested that herpesvirus infection induces nuclear F-actin (orange) that is used by viral capsids (green) for directed motility by recruiting Myosin V (brown), allowing transport through chromatin (gray) to the inner nuclear membrane (black inner circle). In this model, capsids attach and exit the nucleus by budding through the inner nuclear membrane and then fusing with the outer nuclear envelope (outer black circle). b) Our alternative model suggests that herpes infection enlarges nuclear corrals to an average of about 900 nm (white circles). Capsids form stochastically throughout the nucleus early in infection and can diffuse almost unobstructed in these enlarged corrals. In case of pseudorabies virus, the expansion of the viral replication compartment (RC, red) further concentrates capsids to the nuclear periphery. Capsids can reach the inner nuclear membrane by 2 pathways: Either diffusion in corrals located next to the membrane (left side) or by hopping between corrals by active diffusion (right side). In both cases, a high affinity/avidity binding system at the inner nuclear membrane is needed to provide a binding “sink” to capture the stochastic interactions for inner nuclear membrane budding.