

Abstract
Bcl-2 family proteins play a central role in regulating apoptosis. We previously reported that human Bcl-rambo, also termed BCL2L13, localized to mitochondria and induced apoptosis when overexpressed in human embryonic kidney 293T cells. However, the physiological function of Bcl-rambo currently remains unclear. In the present study, human Bcl-rambo was ectopically expressed in Drosophila melanogaster. Bcl-rambo mainly localized to the mitochondria of Drosophila Schneider 2 (S2) cells. The overexpression of Bcl-rambo, but not Bcl-rambo lacking a C-terminal transmembrane domain, induced apoptosis in S2 cells. Moreover, the ectopic expression of Bcl-rambo by a GAL4-UAS system induced aberrant morphological changes characterized by atrophied wing, split thorax, and rough eye phenotypes. Bcl-rambo induced the activation of effector caspases in eye imaginal discs. The rough eye phenotype induced by Bcl-rambo was partly rescued by the co-expression of p35, Diap1, and Diap2. By using this Drosophila model, we showed that human Bcl-rambo interacted genetically with Drosophila homologues of adenine nucleotide translocators and the autophagy-related 8 protein. The results of the present study demonstrated that human Bcl-rambo localized to mitochondria and at least regulated an apoptosis signaling pathway in Drosophila.
Introduction
Programmed cell death plays an essential role in the development and maintenance of tissue homeostasis in animals [1]. Apoptosis is one type of programmed cell death that is mainly regulated by a family of cysteinyl aspartate-specific proteinases (caspases) [2]. Caspases have been classified as initiator caspases (e.g., caspase-8, -9, and -10) and effector caspases (e.g., caspase-3, -6, and -7) [3]. Upon apoptotic stimuli, initiator caspases are activated in platforms referred to as an apoptosome or death-inducing signaling complex, and then cleave effector caspases into their active forms [4,5]. Active effector caspases have been shown to mediate the proteolytic cleavage of many target proteins, leading to the execution of apoptosis [6].
Mitochondria are central in the regulation of the intrinsic apoptosis pathway, which is regulated primarily by Bcl-2 family proteins [7]. Bcl-2 family proteins possess at least one of four Bcl-2 homology (BH) domains, and have been classified into three groups: anti-apoptotic (pro-survival) proteins (e.g., Bcl-2 and Bcl-xL), pro-apoptotic proteins (e.g., Bax and Bak), and BH3-only proteins (e.g., Bad, Bid, and Bim) [8]. Anti-apoptotic proteins interact with pro-apoptotic multidomain proteins, suppressing their activity, whereas BH3-only proteins either interact with anti-apoptotic proteins to inhibit their function or with pro-apoptotic multidomain proteins to stimulate their function [9]. Bcl-2 family proteins have been shown to regulate mitochondrial outer membrane permeabilization, which allows the release of pro-apoptotic proteins, including cytochrome c from the intermembrane space to the cytoplasm [10,11]. Released cytochrome c induces the formation of an apoptosome, thereby promoting the activation of the initiator caspase-9 [4].
We previously identified the widely expressed Bcl-2 family protein Bcl-rambo, also termed BCL2L13 (Fig 1A) [12], and showed that it was composed of four N-terminal BH domains (i.e., BH1, BH2, BH3, and BH4), a unique 250 amino acid extension termed the BHNo domain, and a C-terminal transmembrane domain (TM) [12]. Bcl-rambo localized to mitochondria and induced apoptosis when it was overexpressed in human embryonic kidney (HEK) 293T cells [12]. Bcl-rambo was previously shown to be strongly expressed in various cancer cells, i.e., childhood acute lymphoblastic leukemia, liposarcoma, gastric cancer, and glioblastoma [13–17]. It was also found to be constitutively expressed throughout human early embryonic development [18]. Moreover, it has been reported that Bcl-rambo regulates apoptosis [17,19–22] or induces mitochondrial fragmentation and mitophagy [23]. Although Bcl-rambo has been suggested to either positively or negatively regulate apoptosis, its physiological function remains unclear.
Fig 1. Bcl-rambo localized to mitochondria in Drosophila S2 cells.

(A) Structures of human Bcl-rambo and its mutants. (B) S2 cells were transfected with pMT-V5-His A, pMT-V5-His A/Bcl-rambo, or pMT-V5-His A/Bcl-rambo (ΔTM) and then incubated in the presence of CuSO4 (500 μM) and Z-VAD-fmk (20 μM) for 24 h. S2 cells were stained for Bcl-rambo (green) and with DAPI (blue) and MitoTracker® Red (red). The stained cells in at least five different fields were observed by confocal laser scanning microscopy. Optical sections containing single transfected cells are shown. Data were representative of two independent experiments. Scale bars indicate 10 μm. (C) S2 cells were transfected with pMT-V5-His A, pMT-V5-His A/Bcl-rambo, or pMT-V5-His A/Bcl-rambo (ΔTM) and incubated in the presence of CuSO4 (500 μM) for 24 h. S2 cells were homogenized and separated into PNL, P10, P100, and S100 fractions. The fractions were analyzed by Western blotting using anti-Bcl-rambo, anti-HSP60, and anti-α-tubulin antibodies. Data were representative of three independent experiments.
Drosophila melanogaster is commonly used as a model organism and is a powerful screening tool for investigating genetic interactions. Mammals and flies share some common features for apoptosis signaling pathways [24–26]. Drosophila possesses apoptosis regulators, such as Bcl-2 family proteins, caspases, and inhibitor of apoptosis (IAP) proteins [27–29]. In the present study, we ectopically expressed human Bcl-rambo using a GAL4-UAS expression system in Drosophila. The results obtained showed that Bcl-rambo localized to mitochondria. We also demonstrated that the ectopic expression of Bcl-rambo in transgenic flies induced apoptosis and caused aberrant morphological changes, including a rough eye phenotype.
Materials and Methods
Cells
Drosophila Schneider 2 (S2) cells were grown in Schneider's Drosophila medium (Life Technologies, Grand Island, NY, USA) containing heat-inactivated fetal calf serum (Nichirei Bioscience, Tokyo, Japan) and penicillin-streptomycin mixed solution (Nacalai Tesque, Kyoto, Japan) at 25°C. HEK 293T cells were maintained in DMEM medium (Life Technologies) containing heat-inactivated fetal calf serum and penicillin-streptomycin mixed solution at 37°C.
Plasmids
Human Bcl-rambo (1–485) (Fig 1A) was described previously [12]. Bcl-rambo (1–435, 454–459) designated Bcl-rambo (ΔTM), Bcl-rambo (441–485) designated Bcl-rambo (TM), Bcl-rambo (1–441), and Bcl-rambo (1–459) were generated by PCR amplification (Fig 1A and panel A in S1 Fig). Full-length Bcl-rambo, Bcl-rambo (ΔTM), Bcl-rambo (TM), Bcl-rambo (1–441), Bcl-rambo (1–459), and DsRed-monomer (TAKARA BIO, Shiga, Japan) were inserted into pUAST and/or pMT/V5-His A expression vectors. The genes encoding Drob-1/Debcl and Buffy were generated by PCR amplification using a cDNA library prepared from Drosophila early embryos. Full-length Drob-1/Debcl and Buffy were inserted into pCR3-based expression vectors containing N-terminal FLAG or VSV tags. FLAG-tagged Drob-1 was inserted into pMT/V5-His A expression vectors.
Establishment of transgenic flies
pUAST-Bcl-rambo and pUAST-Bcl-rambo (ΔTM) were injected into embryos to obtain stable transformant lines carrying UAS-Bcl-rambo and UAS-Bcl-rambo (ΔTM). P element-mediated germline transformation was accomplished as described [30]. F1 transformants were selected on the basis of white eye color rescue. Eight and six independent lines were established for UAS-Bcl-rambo and UAS-Bcl-rambo (ΔTM), respectively.
Fly stocks
Fly stocks were maintained at 25°C by standard food (0.7% agar, 5% glucose, and 7% dry yeast). The UAS-Bcl-rambo and UAS-Bcl-rambo (ΔTM) fly lines were established in the present study. The fly lines carrying glass multiple reporter (GMR)-GAL4 were described previously [31]. The engrailed (en)-GAL4 driver fly line was kindly provided by Dr. Nicholas Dyson (Massachusetts General Hospital, Boston, MA, USA). UAS-GFP, UAS-p35, UAS-Diap1, GMR-Diap2, pannier (pnr)-GAL4, decapentaplegic (dpp)-GAL4, salivary gland (sg)-GAL4, Drob-1E26, Drob-1W105, Drob-1E26 BuffyH37, Drob-1W105 BuffyH37, BuffyH37, sesBorg, Ant2G0247 sesBG0247, schlankG0061, and Atg8aEP362 fly lines were obtained from the Bloomington Drosophila stock center (Bloomington, IN, USA) and the Drosophila Genetic Resource Center (Kyoto, Japan). The GAL4 driver fly lines were crossed with the UAS fly lines, developed, and then reared at 28°C.
Antibodies
Antibodies reactive to Bcl-rambo (Rocky-1; Santa Cruz Biotechnology, Santa Cruz, CA, USA), cytochrome c (7H8.2C12; BD Biosciences, San Jose, CA, USA), FLAG (1E6, Wako Pure Chemical Industries, Osaka, Japan), HSP60 (insect) (Enzo Life Science, Farmingdale, NY, USA), α-tubulin (DM1A; Sigma-Aldrich, St. Louis, MI, USA), and VSV-G (P5D4; Santa Cruz Biotechnology) were commercially obtained. Antibodies reactive to Cut (2B10), Discs large (4F3), Elav (7E8A10), and Prospero (MR1A) were obtained from the Developmental Studies Hybridoma Bank (Iowa City, IA, USA). Secondary antibodies conjugated to horseradish peroxidase (HRP) were purchased from Jackson ImmunoResearch (West Grove, PA, USA).
Transfection
S2 cells were transfected with expression vectors by HilyMax transfection reagent (Dojindo, Tokyo, Japan) or siLentfect™ lipid reagent (Bio-Rad Laboratories, Hercules, CA, USA).
Immunostaining
S2 cells were incubated with MitoTracker® Red CMXRos (Lonza, Basel, Switzerland) for 30 min prior to fixation. The caspase inhibitor Z-Val-Ala-Asp(OMe)-fluoromethylketone (Z-VAD-fmk; Peptide Institute, Osaka, Japan) was used to prevent cell death. Tissues were dissected in phosphate-buffered saline (PBS). S2 cells and tissues were fixed with 4% paraformaldehyde (PFA)–PBS and treated with 0.1% and 0.3% Triton X-100–PBS, respectively. The permeabilized cells were blocked with normal goat serum and incubated with primary antibodies and secondary antibodies conjugated with Alexa Fluor 488 (Life Technologies). S2 cells and tissues were then stained with Hoechst 33342 and 4’,6-diamidino-2-phenylindole (DAPI) at final concentrations of 7.5 μM and 1.5 μg/ml, respectively. Samples were analyzed by the confocal laser scanning microscope FV10i (Olympus, Tokyo, Japan).
Subcellular fractionation
S2 cells were transfected with pMT/V5-His A expression vectors encoding Bcl-rambo and Bcl-rambo (ΔTM), and incubated with CuSO4 (500 μM) for 24 h. The cells were washed with PBS and disrupted by a dounce homogenizer (15 strokes) in Hepes-sucrose buffer containing 250 mM sucrose, 10 mM Hepes-KOH (pH 7.4), 1 mM EDTA, 1 mM EGTA, and the protease inhibitor cocktail Complete™ (Roche Diagnostics, Mannheim, Germany). Homogenates were centrifuged (500 x g, 10 min) to remove nuclei and unbroken cells. Supernatants were collected as post nuclear lysates (PNL) and further centrifuged (10,000 x g, 15 min) for separation into precipitates (P10 fraction) and supernatants. The supernatants were centrifuged (100,000 x g, 60 min) and separated into precipitates (P100 fraction) and supernatants (S100 fraction). Alternatively, S2 cells were washed with PBS and treated with digitonin lysis buffer (10 mM Hepes-KOH (pH 7.2), 100 μM digitonin, 250 mM sucrose, 50 mM NaCl, 5 mM EGTA, 2 mM MgCl2, 1 mM dithiothreitol, Complete™) on ice for 15 min. Cell lysates were centrifuged (15,300 x g, 5 min) to separate supernatants as cytosolic fractions. Precipitates were treated with Triton X-100 lysis buffer (50 mM Tris-HCl (pH 7.4), 1% Triton X-100, 2 mM dithiothreitol, 2 mM sodium orthovanadate, and Complete™) on ice for 15 min and centrifuged (15,300 x g, 5min) to remove insoluble materials. Supernatants were collected as organelle fractions containing mitochondria.
Western blotting
Subcellular fractions were prepared as described above. Salivary glands were taken from third instar larva and heated at 95°C for 2 min in 0.11 mM Tris-HCl (pH 7.5) and Complete™. They were homogenized with a dounce homogenizer in the presence of SDS sample buffer. After centrifugation (10, 000 x g, 10 min), supernatants were collected. Protein samples were separated by SDS-PAGE and transferred onto nitrocellulose membranes (Wako Pure Chemical Industries, Osaka, Japan). The membranes were incubated with primary antibodies and then HRP-conjugated secondary antibodies. Protein bands were visualized using ECL Western Blotting Detection Reagents (GE Healthcare, Piscataway, NJ, USA) and analyzed by ImageQuant LAS 4000 mini (GE Healthcare).
Apoptosis assay
S2 cells were stained with Hoechst 33342 (7.5 μM). Nuclear morphology was observed under a fluorescence light microscope (Axiovert 200M, Carl Zeiss, Jena, Germany). Apoptotic cells (%) were calculated as (condensed and/or fragmented nuclei / total nuclei) x 100. In parallel with the apoptosis assay, transfection efficiency was evaluated by transfection with expression vectors encoding DsRed-monomer.
Light microscopy and scanning electron microscopy (SEM)
Adult wings were dehydrated with isopropanol and mounted in Hoyer’s medium. Samples were observed under a light microscope SZX12 (Olympus) or SEM VE-7800 (Keyence, Osaka, Japan) in the low vacuum mode. The phenotypes of the eye and thorax from at least four adult female flies (3 to 5 d old) were observed. The phenotypes of the wings from 4–5 adult female flies (3 to 5 d old) were observed. In these experiments, no significant variations were observed in eye, thorax, and wing phenotypes among the individuals. The L3-L4 area of the wings was quantified by Image J software. The mean of the L3-L4 area was calculated from 4–5 flies and normalized by the mean of the control L3-L4 area. The L3-L4 area (%) was calculated as the mean ± S.E. of three independent experiments. The brightness and contrast of all images taken by the light microscope were adjusted uniformly using Microsoft Office Picture Manager, which was applied equally to all images in each figure.
Caspase assay
Third instar larvae were dissected in PBS, and imaginal discs were suspended in Grace’s insect medium in the presence of Cellevent™ Caspase-3/7 Green Detection Reagent (Life Technologies) at the final concentration of 5 μM for 1 h at 37°C. Eye imaginal discs were fixed with 4% PFA–PBS and permeabilized with 0.5% Triton X-100–PBS, followed by staining with Hoechst 33342 (5 μM). Samples were analyzed by confocal laser scanning microscopy.
Immunoprecipitation
HEK293T cells were transfected with expression vectors by the calcium phosphate method and incubated for in the presence of Z-VAD-fmk (20 μM) for 16 h. Cells were solubilized in Nonidet P-40 lysis buffer (20 mM Tris-HCl (pH 7.4), 150 mM NaCl, 0.5% Nonidet P-40, 10% glycerol, and 2 mM sodium vanadate, Complete™). Postnuclear lysates were precleared with Sepharose 6B for 1 h and then immunoprecipitated with anti-FLAG M2 affinity gels (Sigma-Aldrich) for 3 h. The immunoprecipitates were washed several times with Nonidet P-40 lysis buffer and analyzed by Western blotting.
Statistical analysis
Statistical analyses were performed using an analysis of variance followed by the Tukey’s test for multiple comparisons.
Results
Bcl-rambo localized to mitochondria in Drosophila S2 cells
Bcl-2 family proteins regulate the intrinsic apoptosis pathway via mitochondria. In HEK293T cells, full-length Bcl-rambo (1–485), but not Bcl-rambo lacking the C-terminal TM (1–459), localized to mitochondria [12]. Bcl-rambo, Bcl-rambo (ΔTM), Bcl-rambo (1–441), and Bcl-rambo (1–459) were transiently expressed in Drosophila S2 cells in order to examine whether the C-terminal TM of Bcl-rambo was necessary for localization to mitochondria in Drosophila. Confocal microscopy observations revealed that Bcl-rambo primarily co-localized with MitoTracker (Fig 1B). In contrast, Bcl-rambo (ΔTM) was uniformly distributed over the cytoplasm (Fig 1B). Consistent with this, Bcl-rambo (1–441) and Bcl-rambo (1–459) did not localize to the mitochondria and exhibited a cytoplasmic distribution (panel B in S1 Fig). The localization of Bcl-rambo and Bcl-rambo (ΔTM) was further analyzed by subcellular fractionation. The mitochondrial chaperone HSP60 was present in the P10 fraction and, to a lesser extent, in the S100 fraction, while α-tubulin was only present in the S100 fractions (Fig 1C), indicating that the P10 fraction was enriched with mitochondria. Similar to HSP60, Bcl-rambo was present mainly in the P10 fraction and marginally in the S100 fraction, whereas Bcl-rambo (ΔTM) was only present in the S100 fraction (Fig 1C). These results indicated that Bcl-rambo mainly localized to mitochondria in Drosophila S2 cells.
Bcl-rambo induced apoptosis in Drosophila S2 cells
Previous studies reported that the overexpression of Bcl-rambo induced apoptosis or promoted etoposide- and taxol-induced cell death in mammalian cells [12,19–22]. S2 cells were transiently transfected with pUAST-Bcl-rambo, pUAST-Bcl-rambo (ΔTM) or pUAST-Bcl-rambo (TM) together with pAct5C-GAL4 to investigate whether Bcl-rambo induced apoptosis in Drosophila. We found that Bcl-rambo significantly induced apoptosis, while Bcl-rambo (ΔTM) or Bcl-rambo (TM) only very weakly induced apoptosis (Fig 2A). Since transfection efficiency was calculated to be approximately 17%, it appeared that most S2 cells transfected with Bcl-rambo underwent apoptosis. Moreover, Bcl-rambo (1–441) and Bcl-rambo (1–459) did not induce apoptosis in S2 cells (S2 Fig).
Fig 2. Bcl-rambo induced apoptosis in Drosophila S2 cells.

(A) S2 cells were transfected with pAct5C-GAL4 together with pUAST, pUAST-Bcl-rambo, pUAST-Bcl-rambo (ΔTM), pUAST-Bcl-rambo (TM), or pUAST-DsRed-monomer for 24 h. Cells were stained with Hoechst 33342. Nuclear morphology was observed by fluorescent microscopy. Apoptotic cells (%) are shown as the mean ± S.E. of three independent experiments. **P<0.01, significantly different from the control. n.s., not significant. Transfection efficiency was measured by counting DsRed-monomer-expressing cells, and calculated to be 17.0 ± 0.7% (the mean ± S.E of three independent experiments). (B) S2 cells were transfected with (+) or without (–) pMT-V5-His A/Bcl-rambo (no tag), or pMT-V5-His A/Drob-1 (FLAG tag) for 20 h and then incubated in the presence of CuSO4 (500 μM) for the indicated times. The expression of Bcl-rambo and Drob-1 was analyzed by Western blotting using anti-Bcl-rambo and anti-FLAG antibodies, respectively. Data were representative of three independent experiments. (C) S2 cells were transfected with pMT-V5-His A (open circles), pMT-V5-His A/Bcl-rambo (filled circles), or pMT-V5-His A/Drob-1 (filled squares) for 20 h and then incubated in the presence of CuSO4 (500 μM) for the indicated times. Cells were stained with Hoechst 33342. Apoptotic cells (%) are shown as the mean ± S.E. of three independent experiments. **P<0.01, significantly different from the control. (D) S2 cells were transfected with (+) or without (–) pMT-V5-His A/Bcl-rambo or pMT-V5-His A/Drob-1 for 20 h and then incubated in the presence of CuSO4 (500 μM) for 8 h. The cytosolic fraction and organelle fraction containing mitochondria were both analyzed by Western blotting using anti-cytochrome c (Cyt. c), α-tubulin, and anti-HSP60 antibodies. Data are representative of two independent experiments.
We further investigated the kinetics of apoptosis induced by human Bcl-rambo and the Drosophila pro-apoptotic Bcl-2 family member Drob-1/Debcl/dBorg-1/DBok [32–35]. In S2 cells transiently transfected with expression vectors driven by the metallothionein promoter, the expression of Bcl-rambo and Drob-1 was induced within 4 h of the CuSO4 treatment, and their expression levels were maintained at least up to 16 h (Fig 2B). The ectopic expression of Bcl-rambo or Drob-1 initiated the induction of apoptosis during 4–8 h, and the number of apoptotic cells steadily increased up to 16 h (Fig 2C). Cytochrome c was detected in the cytosolic fraction within 8 h of the CuSO4 treatment in Bcl-rambo- or Drob-1-transfected S2 cells, but not control S2 cells (Fig 2D). These results suggest that Bcl-rambo induces the mitochondrial pathway of apoptosis in Drosophila.
Ectopic expression of Bcl-rambo, but not Bcl-rambo (ΔTM), induced morphological aberrations in Drosophila tissues
In order to gain an insight into the function of Bcl-rambo, it was ectopically overexpressed in several tissues of Drosophila by the GAL4-UAS expression system. GAL4 is expressed along the anterior-posterior compartment boundary under the control of the dpp (blk) promoter in the dpp-GAL4 driver line [36,37]. The ectopic expression of Bcl-rambo using the dpp-GAL4 driver fly lines induced morphological changes in a wing vein (Fig 3A). In particular, the L3-L4 area of Bcl-rambo flies became smaller than that of control (Fig 3B). In contrast, the overexpression of Bcl-rambo (ΔTM) did not cause obviously different phenotypes from those of control (Fig 3A and 3B). In the pnr-GAL4 driver line, GAL4 is expressed in the dorsal mesothorax [38,39]. The expression of Bcl-rambo using the pnr-GAL4 driver fly lines induced a split-thorax phenotype, while no significant differences were observed between control and Bcl-rambo (ΔTM) (Fig 3C). The en gene is expressed in the posterior compartment of each segment of the embryo and plays an essential role in the development stages of Drosophila. The overexpression of Bcl-rambo using the en-GAL4 driver fly lines induced embryonic lethality, and no adult flies expressing Bcl-rambo were observed (Table 1). In contrast, Bcl-rambo (ΔTM) did not affect the number of adult flies (Table 1). In Drosophila, salivary gland is suitable for preparing cell lysates without the contamination of other tissues. The amount of Bcl-rambo and Bcl-rambo (ΔTM) at the protein level was almost equivalent in the sg-GAL4 driver fly lines (Fig 3D and 3E). Thus, these results indicated that the ectopic expression of Bcl-rambo interfered with the proper differentiation of the wing and thorax as well as embryonic development.
Fig 3. Ectopic expression of Bcl-rambo, but not Bcl-rambo (ΔTM), induced aberrant morphological changes in Drosophila.

(A and B) Bcl-rambo and Bcl-rambo (ΔTM) were expressed using dpp-GAL4 driver fly lines. (a) w; +; dpp-GAL4/+, (b) w; UAS-Bcl-rambo/+; dpp-GAL4/+, (c) w; UAS-Bcl-rambo(ΔTM)/+; dpp-GAL4/+. The morphology of the wings was observed under light microscopy. The white arrow indicates morphological aberrations in the wing vein. The L3-L4 area was measured by Image J software (B). Data are shown as the mean ± S.E. of three independent experiments (n = 4–5 for each experiment). **P<0.05, significantly different from control. n.s., not significant. (C) Bcl-rambo and Bcl-rambo (ΔTM) were expressed using pnr-GAL4 driver lines. (a) w; +; pnr-GAL4/+, (b) w; UAS-Bcl-rambo/+; pnr-GAL4/+, (c) w; UAS-Bcl-rambo (ΔTM)/+; pnr-GAL4/+. The morphology of adult thoraxes was observed by SEM. Scale bars indicate 100 μm. Data were representative of three independent experiments. (D and E) Cell lysates of third instar larval salivary glands were prepared from sg-GAL4/w; +; +, sg-GAL4/w; UAS-Bcl-rambo/+; +, and sg-GAL4/w; UAS-Bcl-rambo (ΔTM)/+; + flies, and analyzed by Western blotting. The amount of Bcl-rambo and Bcl-rambo (ΔTM) was normalized to that of α-tubulin. The protein expression of Bcl-rambo (%) is shown as the mean ± S.E. of three independent experiments.
Table 1. Phenotypes of transgenic flies expressing Bcl-rambo or Bcl-rambo (ΔTM).
| GAL4 driver | Name of Transgene | Strain | Chromosome | Phenotype |
|---|---|---|---|---|
| dpp | Bcl-rambo | #77 | II | Atrophied wing |
| dpp | Bcl-rambo (ΔTM) | #45 | II | n.d. |
| pnr | Bcl-rambo | #77 | II | Split thorax |
| pnr | Bcl-rambo (ΔTM) | #45 | II | n.d. |
| GMR | Bcl-rambo | #1 | III | Rough eye |
| GMR | Bcl-rambo | #6 | X | Rough eye |
| GMR | Bcl-rambo | #46 | II | Rough eye |
| GMR | Bcl-rambo | #77 | II | Rough eye |
| GMR | Bcl-rambo (ΔTM) | #10 | II | n.d. |
| GMR | Bcl-rambo (ΔTM) | #26 | II | n.d. |
| GMR | Bcl-rambo (ΔTM) | #45 | II | n.d. |
| GMR | Bcl-rambo (ΔTM) | #148 | X | n.d. |
| en | Bcl-rambo | #4 | X | Lethal |
| en | Bcl-rambo | #7 | III | Lethal |
| en | Bcl-rambo | #46 | II | Lethal |
| en | Bcl-rambo | #77 | II | Lethal |
| en | Bcl-rambo (ΔTM) | #22 | X | n.d. |
| en | Bcl-rambo (ΔTM) | #45 | II | n.d. |
| en | Bcl-rambo (ΔTM) | #112 | III | n.d. |
n.d.: no detectable phenotype
Ectopic expression of Bcl-rambo, but not Bcl-rambo (ΔTM), induced a rough eye phenotype in Drosophila
The expression of Bcl-rambo using the GMR-GAL4 driver fly lines induced a rough eye phenotype, such as a reduction in eye size and the loss of ommatidia, bristles, and pigmentation (Fig 4A). In contrast, Bcl-rambo (ΔTM) did not exert any phenotypic changes from control and green fluorescent protein (GFP) (Fig 4A). Bcl-rambo and Bcl-rambo (ΔTM) were similarly expressed in eye imaginal discs (Fig 4B), excluding the possibility that Bcl-rambo (ΔTM) was not or insufficiently expressed. A rough eye phenotype was also observed in other independent transgenic lines for Bcl-rambo, but not Bcl-rambo (ΔTM) (Table 1 and S3 Fig), which ruled out position effects.
Fig 4. Ectopic expression of Bcl-rambo, but not Bcl-rambo (ΔTM), induced a rough eye phenotype in Drosophila.
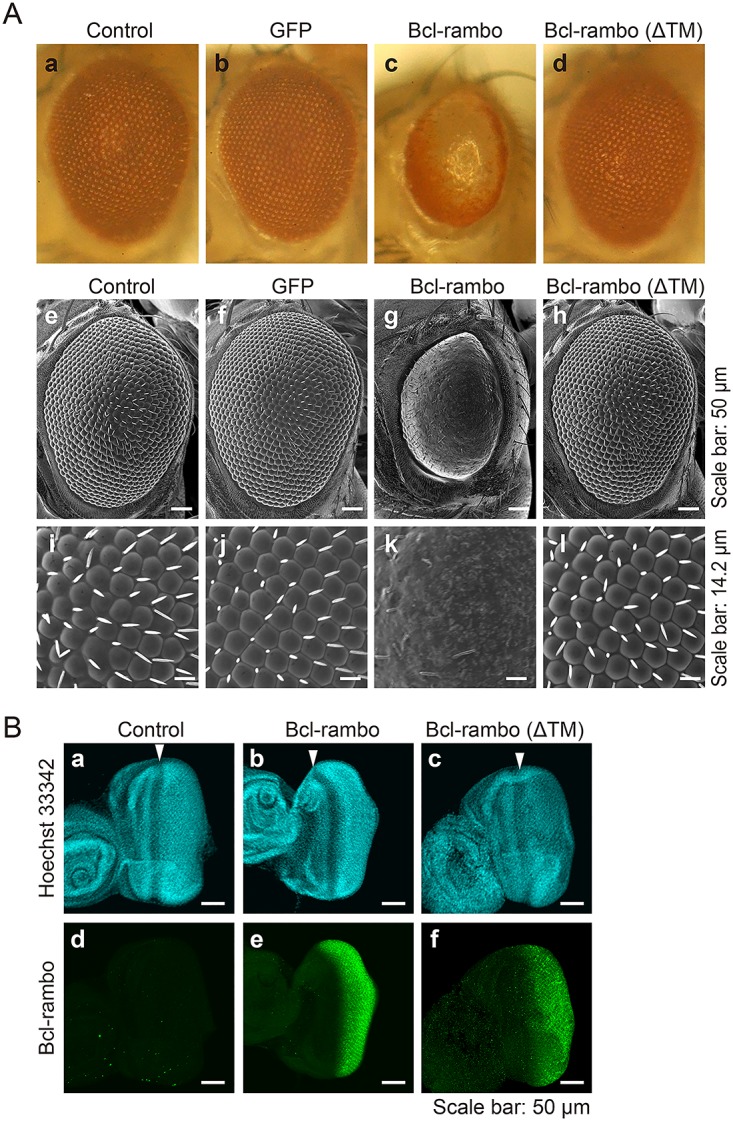
(A) Bcl-rambo and Bcl-rambo (ΔTM) were expressed using GMR-GAL4 driver fly lines. (a, e, i) GMR-GAL4/w; +/CyO; +, (b, f, j) GMR-GAL4/w; UAS-GFP/+; +, (c, g, k) GMR-GAL4/w; UAS-Bcl-rambo/CyO; +, (d, h, l) GMR-GAL4/w; UAS-Bcl-rambo (ΔTM)/CyO; +. The morphology of adult eyes was observed by light microscopy (a–d) and SEM (e–l). Scale bars in e–h and i–l indicate 50 μm and 14.2 μm, respectively. Light microscopy and SEM photographs were taken from different individuals. Data were representative of three independent experiments. (B) Eye imaginal discs were stained for Bcl-rambo (green) and with Hoechst 33342 (blue). (a and d) GMR-GAL4/w; +/CyO or Sp; +, (b and e) GMR-GAL4/w; UAS-Bcl-rambo/CyO or Sp; +, (c and f) GMR-GAL4/w; UAS-Bcl-rambo (ΔTM)/CyO or Sp; +. White arrows show the morphogenetic furrow. Scale bars indicate 50 μm. Data were representative of three independent experiments.
Ectopic expression of Bcl-rambo induced activation of effector caspases in Drosophila
Bcl-rambo was found to induce apoptosis in S2 cells (Fig 2). Drosophila effector caspases have been shown to exhibit DEVD-cleaving activity in a manner similar to mammalian effector caspases [40,41]. In order to determine whether the ectopic expression of Bcl-rambo caused caspase activation, a fluorogenic substrate for caspase-3/7 was used to visualize activated effector caspases. When Bcl-rambo was expressed in eye imaginal discs, the activated effector caspases greatly increased from the posterior region to the morphogenetic furrow (Fig 5A and 5B). However, no obvious signals of activated caspases were detected when Bcl-rambo (ΔTM) was expressed (Fig 5A and 5B). These results indicated that the ectopic expression of Bcl-rambo, but not Bcl-rambo (ΔTM), significantly induced the activation of effector caspases.
Fig 5. Ectopic expression of Bcl-rambo, but not Bcl-rambo (ΔTM), induced activation of effector caspases in eye imaginal discs.

(A and B) (a, d) GMR-GAL4/w; +/CyO or Sp; +, (b, e) GMR-GAL4/w; UAS-Bcl-rambo/CyO or Sp; +, (c, f) GMR-GAL4/w; UAS-Bcl-rambo (ΔTM)/CyO or Sp; +. The eye imaginal discs were labeled for activated caspase-3/7 (green) and with Hoechst 33342 (blue). Scale bars indicate 50 μm. White arrows indicate the morphogenetic furrow. The number of fluorescent cells harboring activated caspases from the morphogenetic furrow to the posterior region of imaginal eye discs was measured (B). Data are shown as the mean ± S.E. of three independent experiments (n = 6 for each experiment). **P<0.01, significantly different from control. n.s., not significant.
Reduced pigmentation induced by Bcl-rambo was rescued by p35, Diap-1, and Diap-2
The baculovirus p35 and Drosophila Diap1 and Diap2 have been shown to inhibit caspase activity and regulate caspase-dependent apoptosis [28,42]. We further investigated whether the rough eye phenotype induced by Bcl-rambo was rescued by p35, Diap1, and Diap2. The reduction in eye pigments induced by Bcl-rambo was rescued by the co-expression of p35, Diap1, and Diap2 (Fig 6A). In contrast, the co-expression of p35, Diap1, and Diap2 only weakly suppressed the abnormal formation of ommatidia and bristles as well as the reduction in eye size induced by Bcl-rambo (Fig 6A). Bcl-rambo (ΔTM) did not cause any changes with the co-expression of p35, Diap1, and Diap2 (Fig 6B). Moreover, additional UAS repeats in the UAS-GFP insertion did not rescue the rough eye phenotype induced by Bcl-rambo (S4 Fig), excluding the effect of diluting GMR-GAL4.
Fig 6. Reduced pigmentation induced by Bcl-rambo was rescued by co-expression of p35, Diap1, and Diap2.
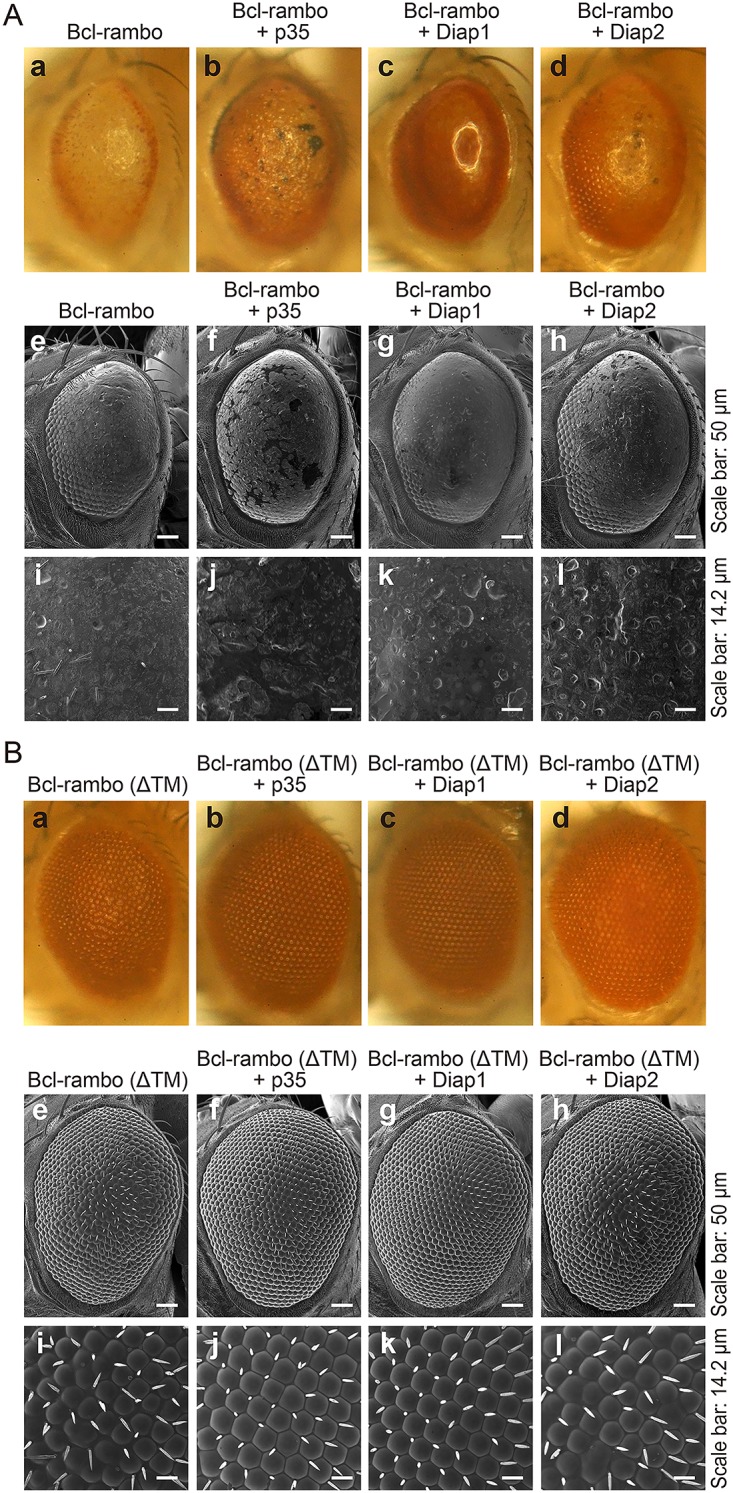
(A and B) Bcl-rambo and Bcl-rambo (ΔTM) were expressed using GMR-GAL4 driver fly lines. (A) (a, e, i) GMR-GAL4/w; UAS-Bcl-rambo/+; +, (b, f, j) GMR-GAL4/w; UAS-Bcl-rambo/+; UAS-p35/+, (c, g, k) GMR-GAL4/w; UAS-Bcl-rambo/+; UAS-Diap1/+, (d, h, l) GMR-GAL4/w; UAS-Bcl-rambo/+; GMR-Diap2/+. (B) (a, e, i) GMR-GAL4/w; UAS-Bcl-rambo (ΔTM)/+; +, (b, f, j) GMR-GAL4/w; UAS-Bcl-rambo (ΔTM)/+; UAS-p35/+, (c, g, k) GMR-GAL4/w; UAS-Bcl-rambo (ΔTM)/+; UAS-Diap1/+, (d, h, l) GMR-GAL4/w; UAS-Bcl-rambo (ΔTM)/+; GMR-Diap2/+. The morphology of adult eyes was observed by light microscopy (a–d) and SEM (e–l). Scale bars in e–h and i–l indicate 50 μm and 14.2 μm, respectively. Light microscopy and SEM photographs were taken from different individuals. Data were representative of three independent experiments.
Aberrant morphology of ommatidia induced by Bcl-rambo was partly rescued by p35
The Drosophila compound eye is composed of eight photoreceptor cells, four cone cells, two primary pigment cells, six secondary pigment cells, three tertiary pigment cells, and three mechanosensory bristles. The differentiation of these cells was completed ~42 h after pupal formation at 28°C. Since the loss-of-pigment phenotype induced by Bcl-rambo was effectively rescued by the co-expression of caspase inhibitors, we examined the morphology of the Drosophila pupal retina 42 h after pupal formation. When Bcl-rambo was expressed using the GMR-GAL4 driver fly lines, the shapes of the cone cells and pigment cells were markedly altered because four cone cells per ommatidium and the surrounding pigment cells were hardly distinguishable (Fig 7A and 7B). In contrast, the morphology of the pupal retina expressing Bcl-rambo (ΔTM) was not obviously changed, but showed a slight defect in the ommatidia rotation (Fig 7A). The aberrant morphology of ommatidia induced by Bcl-rambo was partly rescued by p35 (Fig 7A and 7B).
Fig 7. Aberrant morphological changes in ommatidia induced by Bcl-rambo were partly rescued by p35.

(A and B) Pupal retinae 42 h after pupal formation were stained with an anti-discs large antibody (green). (a) w/+; +; +, (b) GMR-GAL4/w; UAS-Bcl-rambo/+; +, (c) GMR-GAL4/w; UAS-Bcl-rambo/+; UAS-p35/+, (d) GMR-GAL4/w; UAS-Bcl-rambo (ΔTM)/+; +, (e) GMR-GAL4/w; UAS-Bcl-rambo (ΔTM)/+; UAS-p35/+. Scale bars indicate 10 μm. The numbers of normal and aberrant ommatidia were counted. Aberrant morphology (%) is shown as the mean ± S.E. of three independent experiments (n = 5 for each experiment). **P<0.01. n.s., not significant.
We further investigated whether Bcl-rambo induced apoptosis in the pupal retinae at the same period. The signals for activated caspase-3/7 were rarely detected in the control pupal retinae, and they were not increased in the pupal retinae when Bcl-rambo or Bcl-rambo (ΔTM) was expressed (data not shown). These results suggest that Bcl-rambo does not induce apoptosis in the pupal retinae at this period.
In order to determine whether Bcl-rambo affected eye development, eye imaginal discs were stained with Elav for a marker of all photoreceptor cells, Prospero for a marker of R7 cells, and Cut for a maker of cone cell precursors. Elav, Prospero, and Cut were expressed in eye imaginal discs even when Bcl-rambo or Bcl-rambo (ΔTM) was expressed (Fig 8A and 8B). These results suggested that Bcl-rambo did not affect the differentiation of photoreceptor cells in the eye discs of third instar larvae. Although the formation of Cut-positive cone cell precursors was not affected by Bcl-rambo (Fig 8B), the differentiation of cone cells and pigment cells were clearly affected by the expression of Bcl-rambo in pupal retinae (Fig 7A). The spacing of Elav clusters appeared to be abnormal (Fig 8A and 8B), and this may have been due to the death of interommatidial cells. Therefore, the primary effect of Bcl-rambo appears to be the induction of apoptosis, while defects in the differentiation of cone cells and pigment cells may be secondary effects during the later stage of eye development.
Fig 8. Bcl-rambo did not affect the expression of Cut, Elav, or Prospero.
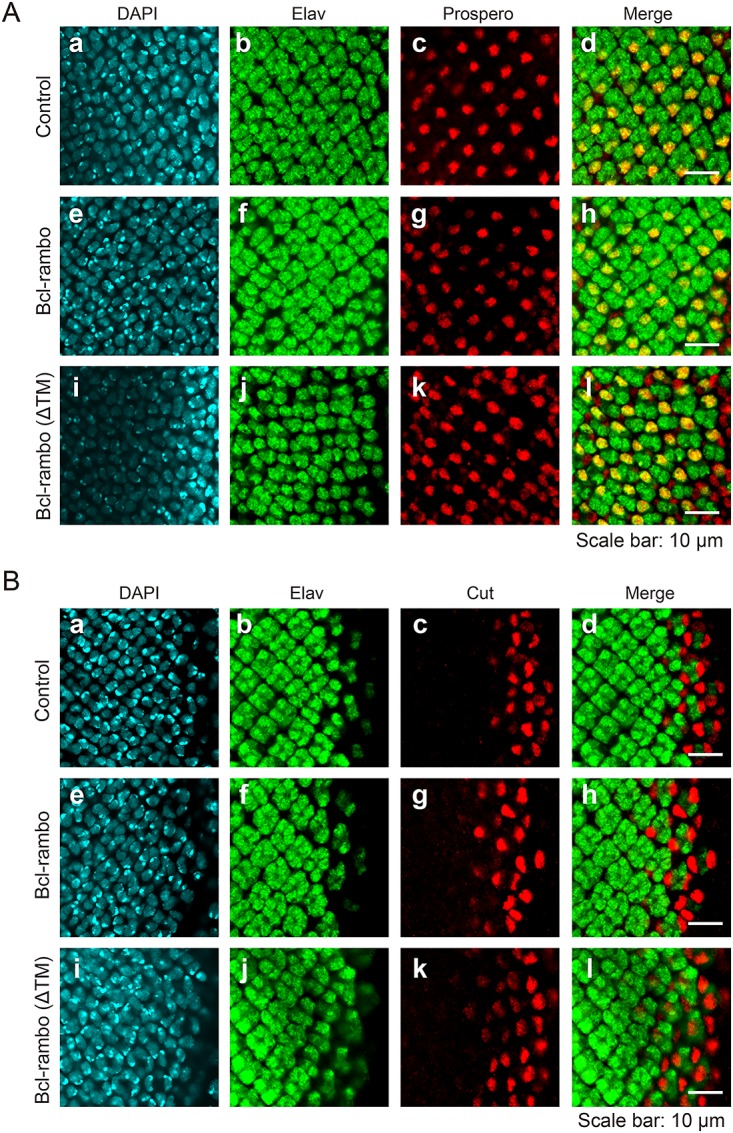
(A and B) Eye imaginal discs were stained for DAPI (blue) and Elav (green), together with Prospero (red) (A) or Cut (red) (B). (a–d) GMR-GAL4/w; +/CyO or Sp; +, (e–h) GMR-GAL4/w; UAS-Bcl-rambo/CyO or Sp; +, (i–l) GMR-GAL4/w; UAS-Bcl-rambo (ΔTM)/CyO or Sp; +. Scale bars indicate 10 μm. Data were representative of at least two independent experiments.
Bcl-rambo did not interact with Drob-1 or Buffy
Drosophila has two Bcl-2 family proteins: Drob-1/Debcl and Buffy/dBorg-2, both of which have been classified into pro-apoptotic multidomain proteins, and are closely related to mammalian Bok [32–35]. We investigated whether Bcl-rambo interacted genetically with Drob-1 and Buffy. The rough eye phenotype induced by Bcl-rambo was little if any influenced by the Drob-1E26 or Drob-1W105 mutation or by the Drob-1E26 BuffyH37 or Drob-1W105 BuffyH37 mutation (Fig 9A). Bcl-rambo (ΔTM) did not exhibit any phenotypic changes in Drob-1E26 BuffyH37, or Drob-1W105 BuffyH37 flies (Fig 9B). Moreover, the BuffyH37 mutation did not influence the rough eye phenotype induced by Bcl-rambo (S5 Fig).
Fig 9. Bcl-rambo did not interact genetically with Drob-1 or Buffy.
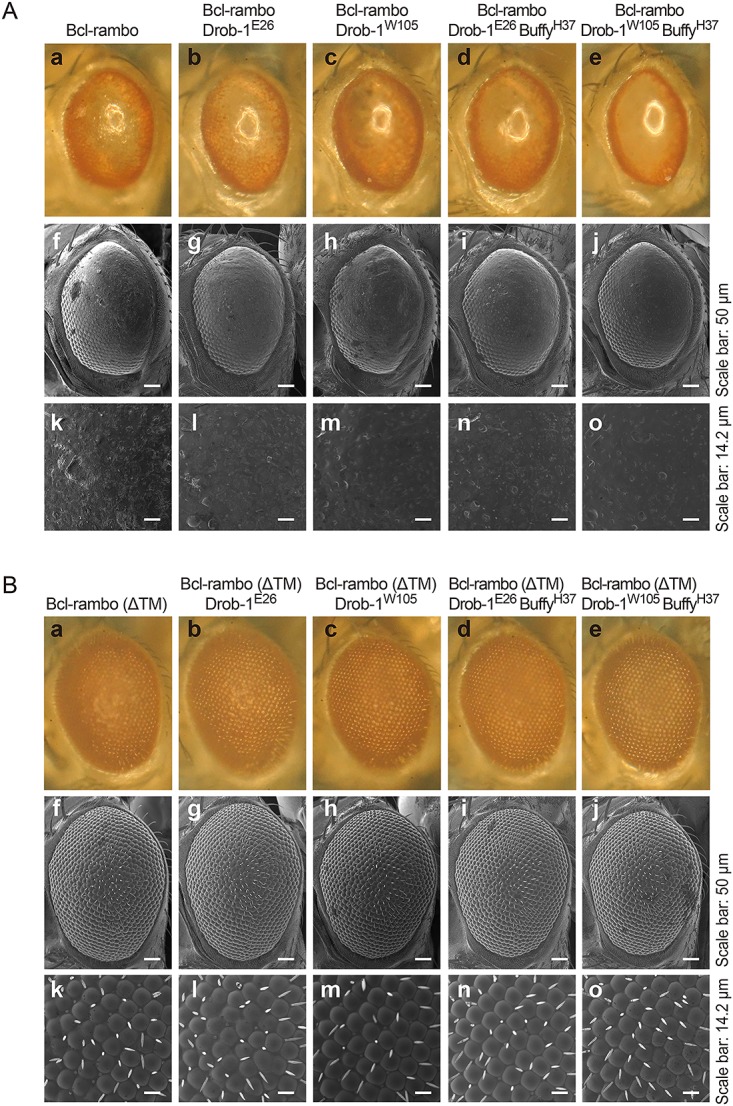
(A and B) Bcl-rambo and Bcl-rambo (ΔTM) were expressed using GMR-GAL4 driver fly lines. (A) (a, f, k) GMR-GAL4/w; UAS-Bcl-rambo/+; +, (b, g, l) GMR-GAL4/w; UAS-Bcl-rambo/Drob-1E26; +, (c, h, m) GMR-GAL4/w; UAS-Bcl-rambo/Drob-1W105; +, (d, i, n) GMR-GAL4/w; UAS-Bcl-rambo/Drob-1E26 BuffyH37; +, (e, j, o) GMR-GAL4/w; UAS-Bcl-rambo/Drob-1W105 BuffyH37; +. (B) (a, f, k) GMR-GAL4/w; UAS-Bcl-rambo (ΔTM)/+; +, (b, g, l) GMR-GAL4/w; UAS-Bcl-rambo (ΔTM)/Drob-1E26; +, (c, h, m) GMR-GAL4/w; UAS-Bcl-rambo (ΔTM)/Drob-1W105; +, (d, i, n) GMR-GAL4/w; UAS-Bcl-rambo (ΔTM)/Drob-1E26 BuffyH37; +, (e, j, o) GMR-GAL4/w; UAS-Bcl-rambo (ΔTM)/Drob-1W105 BuffyH37; +. The morphology of the adult eyes was observed by light microscopy (a–e) and SEM (f–o). Scale bars in f–j and k–o indicate 50 μm and 14.2 μm, respectively. Data were representative of three independent experiments.
An immunoprecipitation assay was performed to confirm the weak genetic interaction between Bcl-rambo and either Drob-1/Debcl or Buffy. FLAG-tagged Bcl-rambo and FLAG-tagged Buffy were transiently overexpressed together with VSV-tagged Drob-1/Debcl or VSV-tagged Buffy in HEK293T cells, and FLAG-tagged proteins were pulled down. Bcl-rambo did not bind to Drob-1/Debcl or Buffy in HEK293T cells, while Buffy bound to Drob-1/Debcl (Fig 10).
Fig 10. Bcl-rambo did not bind to Drob-1/Debcl or Buffy.

HEK293T cells were transfected with (+) or without (–) expression vectors for FLAG-tagged Bcl-rambo or FLAG-tagged Buffy together with VSV-tagged Buffy or VSV-tagged Drob-1/Debcl for in the presence of zVAD-fmk (20 μM) for 16 h. Cell lysates were immunoprecipitated with anti-FLAG antibody-conjugated beads. Immunoprecipitates (IP) were analyzed by Western blotting using an anti-VSV antibody. Cell lysates were analyzed by Western blotting using anti-FLAG, anti-VSV, and β-actin antibodies. Data were representative of three independent experiments.
Bcl-rambo interacted genetically with adenine nucleotide translocators and the autophagy-related 8a protein
Recent studies reported that Bcl-rambo bound to adenine nucleotide translocators [21], ceramide synthases 2 (CerS2) and 6 (CerS6) [17], and LC3, a mammalian homologue of the autophagy-related 8 (Atg8) protein [23]. We herein investigated whether Bcl-rambo interacted genetically with stress-sensitive B (SesB) and Ant2 (a major Drosophila homologue of adenine nucleotide translocators and the second isogene, respectively), Schlank (a Drosophila homologue of ceramide synthases), and Drosophila Atg8a. The loss-of-pigment phenotype induced by Bcl-rambo was effectively rescued by the sesBorg or Ant2G0247 sesBG0247 mutation (Fig 11A), while Bcl-rambo (ΔTM) did not exhibit any phenotypic changes in sesBorg or Ant2G0247 sesBG0247 flies (Fig 11B). The schlankG0061 mutation did not rescue the loss-of-pigment phenotype induced by Bcl-rambo (Fig 11C). The Atg8aEP362 mutation rescued the loss-of-pigmentation induced by Bcl-rambo, and partly suppressed the loss of bristles induced by Bcl-rambo, while Bcl-rambo (ΔTM) flies did not exhibit the clear effects observed in Atg8aEP362 flies (Fig 11D). Due to the insertion of UAS repeats in Atg8aEP362 flies, Atg8a may be overexpressed in the presence of the GMR-GAL4 driver.
Fig 11. Reduced pigmentation induced by Bcl-rambo was rescued by mutations in adenine nucleotide translocators and Atg8a.
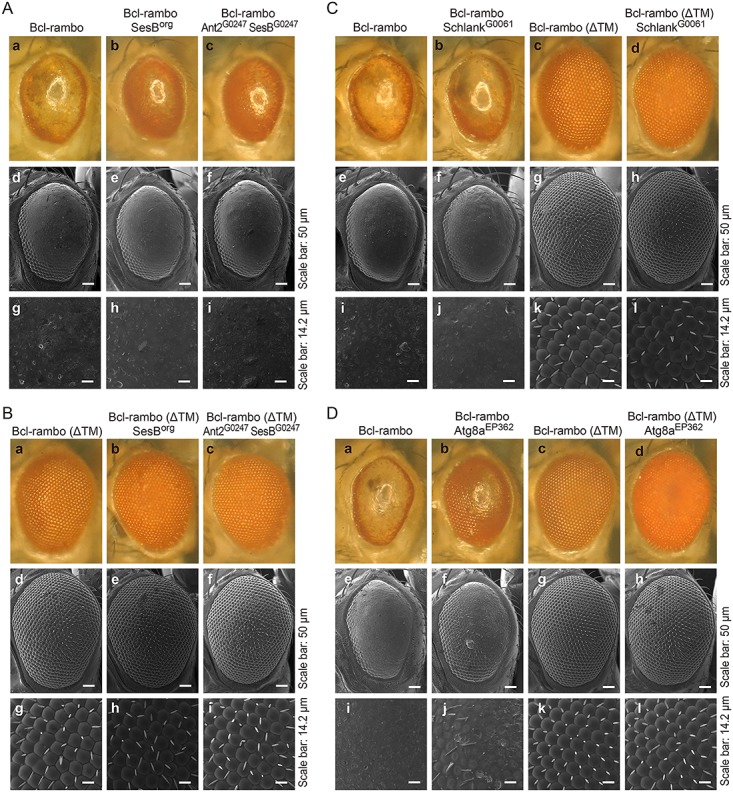
(A to C) (A) (a, d, g) GMR-GAL4/w; UAS-Bcl-rambo/+; +, (b, e, h) sesBorg/GMR-GAL4; UAS-Bcl-rambo/+; +, (c, f, i) w67c23 P{lacW}Ant2G0247 sesBG0247/GMR-GAL4; UAS-Bcl-rambo/+; +. (B) (a, d, g) GMR-GAL4/w; UAS-Bcl-rambo (ΔTM)/+; +, (b, e, h) sesBorg/GMR-GAL4; UAS-Bcl-rambo (ΔTM)/+; +, (c, f, i) w67c23 P{lacW}Ant2G0247 sesBG0247/GMR-GAL4; UAS-Bcl-rambo (ΔTM)/+; +. The morphology of adult eyes was observed by light microscopy (a–c) and SEM (d–i). (C) (a, e, i) GMR-GAL4/w; UAS-Bcl-rambo/+; +, (b, f, j) w67c23 P{lacW}schlankG0061 schlankG0061/GMR-GAL4; UAS-Bcl-rambo/+; + (c, g, k) GMR-GAL4/w; UAS-Bcl-rambo (ΔTM)/+; +, (d, h, l) w67c23 P{lacW}schlankG0061 schlankG0061/GMR-GAL4; UAS-Bcl-rambo (ΔTM)/+; +. The morphology of adult eyes was observed by light microscopy (a–d) and SEM (e–l). Scale bars in e–h and i–l indicate 50 μm and 14.2 μm, respectively. Data were representative of two independent experiments. (D) (a, e, i) GMR-GAL4/w; UAS-Bcl-rambo/+; +, (b, f, j) w1118 P{EP}Atg8aEP362/GMR-GAL4; UAS-Bcl-rambo/+; + (c, g, k) GMR-GAL4/w; UAS-Bcl-rambo (ΔTM)/+; +, (d, h, l) w1118 P{EP}Atg8aEP362/GMR-GAL4; UAS-Bcl-rambo (ΔTM)/+; +. The morphology of adult eyes was observed by light microscopy (a–d) and SEM (e–l). Scale bars in e–h and i–l indicate 50 μm and 14.2 μm, respectively. Data were representative of three independent experiments.
Discussion
We previously identified human Bcl-rambo and showed that its overexpression induced apoptosis in HEK293T cells [12]. Consistent with these findings, previous studies reported that Bcl-rambo induced apoptosis in other cell lines [20–22]. However, Bcl-rambo has been also shown to inhibit pro-apoptotic ceramide synthases in glioblastoma [17]. Bcl-rambo is known to be strongly expressed in several cancer cells [13–17], implying that Bcl-rambo may inhibit apoptosis or promote growth. Thus, it currently remains unclear whether Bcl-rambo plays either a pro-apoptotic role or anti-apoptotic role or even both roles under physiological conditions. In order to gain an insight into this issue, we investigated the biological activity of human Bcl-rambo in Drosophila. The ectopic expression of Bcl-rambo induced apoptosis in Drosophila S2 cells. Bcl-rambo also induced apoptosis and morphological aberrations representative of the rough eye phenotype in transgenic flies.
Bcl-rambo, but not Bcl-rambo (ΔTM), mainly localized to mitochondria in Drosophila S2 cells. This result was consistent with our previous findings obtained using HEK293T cells [12], and indicated that the C-terminal TM was essential for the mitochondrial localization of Bcl-rambo. A previous study showed that the mitochondrial targeting of Bcl-xL required the C-terminal TM domain to be flanked at both ends by at least two basic amino acids [43]. This consensus sequence proposed for mitochondrial localization was found to be conserved in human Bcl-rambo [12]. Drosophila has two Bcl-2 family proteins Drob-1/Debcl and Buffy [32–35]. Drob-1/Debcl has been shown to predominantly localize to mitochondria, whereas Buffy localizes to the endoplasmic reticulum in Drosophila and mammalian cell lines [44]. Drob-1/Debcl, but not Buffy, has C-terminal positively-charged residues homologous with Bcl-xL [44]. Thus, Bcl-rambo localizes to mitochondria in a manner similar to Drob-1/Debcl in Drosophila.
In Drosophila, Bcl-2 family members were found to not play a central role in the regulation of apoptosis [9,26]. Drob-1/Debcl and Buffy are not required for normal development, but participate in stress-induced apoptosis [45]. The ectopic expression of Drob-1/Debcl induced apoptosis in cultured cells and transgenic flies [32–35], indicating that Drob-1/Debcl exhibits pro-apoptotic activity. In contrast, Buffy has an anti-apoptotic function and has been shown to interact genetically and physically with Drob-1/Debcl to suppress Drob-1/Debcl-induced cell death [46]. A previous study reported that Drob-1/Debcl protected neurons from polyglutamine-induced toxicity, while Buffy enhanced it [47]. These findings suggested that Drob-1/Debcl and Buffy possess both pro-apoptotic and anti-apoptotic activities. The ectopic expression of Bcl-rambo, but not Bcl-rambo (ΔTM), induced apoptosis in Drosophila S2 cells and caspase activation in eye imaginal discs. This is consistent with our previous findings in which Bcl-rambo induced caspase activation in HEK293T cells [12]. In S2 cells, Bcl-rambo and Drob-1/Debcl induced the release of cytochrome c into the cytosol. These results suggest that Bcl-rambo regulates the mitochondrial signaling pathway of apoptosis.
Human Bcl-2 and murine Bax have been shown to play roles in the regulation of apoptosis in the Drosophila eye and wing [48]. Human Bcl-2 is known to suppress murine Bax- and reaper-induced mitochondrial defects [49]. Furthermore, Drob-1/Debcl is required for cell death by murine Bax in the Drosophila eye [50]. These findings collectively suggest that the apoptosis pathway regulated by Bcl-2 family proteins is highly conserved between mammals and Drosophila. We showed that Bcl-rambo did not interact with anti-apoptotic or pro-apoptotic Bcl-2 family members in HEK293T cells [12]. Consistent with this result, Bcl-rambo interacted genetically with Drob-1/Debcl or Buffy only weakly in Drosophila. In addition, Bcl-rambo did not bind to Drob-1/Debcl or Buffy in HEK293T cells. Since the BHNo domain of Bcl-rambo was previously shown to induce caspase activation in HEK293T cells [12], the BHNo domain may be essential for the induction of apoptosis in Drosophila.
Bcl-rambo has been reported to regulate apoptosis by interacting with adenine nucleotide translocators [21], ceramide synthases 2 (CerS2) and 6 (CerS6) [17] and the Legionella protein SidF [20]. Adenine nucleotide translocators are components of the permeability transition pore complex in mitochondria, and play a key role in cell death [51]. In Drosophila, adenine nucleotide translocators are encoded by sesB as a major isoform and the second isogene Ant2 [52]. Ceramide synthases have been shown to regulate sphingolipid metabolism and also play a role in regulating cell death [53,54]. In subcellular localization, ceramide synthase activities have been detected in mitochondrial and microsomal fractions [54]. Schlank is the Drosophila member of the ceramide synthase family and controls growth and body fat [55]. Due to its localization to mitochondria, it was possible to postulate that Bcl-rambo may regulate the mitochondrial apoptosis pathway via adenine nucleotide translocators or ceramide synthases. The genetic interaction between Bcl-rambo and SesB, but not Schlank suggests that Bcl-rambo regulates the mitochondrial apoptosis pathway primarily via adenine nucleotide transporters in Drosophila.
The baculovirus p35 and Drosophila Diap1 and Diap2 have been shown to inhibit caspases in order to prevent apoptosis [28,42]. Bcl-rambo induced caspase activity in the eye imaginal discs of third instar larvae, but not the pupal retinae, suggesting that Bcl-rambo has a stronger effect on dividing cells. The ectopic expression of Bcl-rambo induced the rough eye phenotype in Drosophila, accompanied by a reduction in eye size and the loss of eye pigment. Although the reduced eye size caused by Bcl-rambo was only weakly rescued by p35, Diap1, and Diap2, reduced pigmentation was rescued by p35, Diap1 and Diap2. The shapes of cone cells and pigment cells were markedly altered by Bcl-rambo, and their morphologies were also partially rescued by p35 in the pupal retinae. Thus, the overexpression of Bcl-rambo may have multiple effects on cells, and the loss of cells may be secondary to defects rescued by p35. Previous studies reported that the ectopic expression of Drob-1/Debcl resulted in the rough eye phenotype [33,34], while the rough eye phenotype induced by Drob-1/Debcl was inhibited by p35 [33]. Drob-1/Debcl-induced cell death was not antagonized by p35 in Drosophila S2 cells [34], while it was rescued by p35 in CHO cells [32]. These findings suggest that the role of p35 in Drob-1/Debcl-induced cell death may be influenced by the cellular context. In addition to apoptosis, necrosis and autophagic cell death have been observed during Drosophila development [56,57]. Drob-1/Debcl and Buffy are associated with germinal cell death characterized with the mixed morphologies of apoptosis and necrosis [58]. Thus, the ectopic expression of Bcl-rambo may induce caspase-dependent apoptosis and caspase-independent cell death in Drosophila.
In conclusion, the ectopic expression of Bcl-rambo induced the rough eye phenotype in transgenic flies. Bcl-rambo induced the rough eye phenotype, which suggested that a functional homologue of Bcl-rambo may exist in Drosophila even though amino acid homology is small. A recent study showed that Bcl-rambo had a short WXXL/I motif and was a mammalian homologue of yeast Atg32 that mediated mitophagy and mitochondrial fragmentation [23]. Bcl-rambo has been reported to interact with adenine nucleotide translocators, ceramide synthases, and LC3 (a mammalian homologue of Atg8) [17,21,23], and homologues of these proteins are present in Drosophila. Thus, humans and Drosophila may have a similar pathway regulated by Bcl-rambo. By using Drosophila as a screening system, we revealed that human Bcl-rambo interacted genetically with Drosophila homologues of adenine nucleotide translocators and Atg8, suggesting that Bcl-rambo mediates at least two different pathways for apoptosis and mitophagy in Drosophila. These results indicate that the Drosophila model established in the present study can be a powerful screening tool for investigating genetic interactions. The further identification of gene(s) interacting with Bcl-rambo may help to address its physiological function in humans.
Supporting Information
(A) Structures of human Bcl-rambo and its mutants. (B) S2 cells were transfected with pMT-V5-His A, pMT-V5-His A/Bcl-rambo (1–441), or pMT-V5-His A/Bcl-rambo (1–459) and then incubated in the presence of CuSO4 (500 μM) and Z-VAD-fmk (20 μM) for 24 h. S2 cells were stained for Bcl-rambo (green) and with DAPI (blue) and MitoTracker® Red (red). The stained cells in at least five different fields were observed by confocal laser scanning microscopy. Optical sections containing a single transfected cell are shown. Data were representative of two independent experiments. Scale bars indicate 10 μm.
(TIF)
(A) S2 cells were transfected with pAct5C-GAL4 together with pMT-V5-His A, pMT-V5-His A/Bcl-rambo, pMT-V5-His A/Bcl-rambo (ΔTM), pMT-V5-His A/Bcl-rambo (1–441), pMT-V5-His A/Bcl-rambo (1–459), or pUAST-DsRed-monomer in the presence of CuSO4 (500 μM) for 24 h. Cells were stained with Hoechst 33342. Nuclear morphology was observed by fluorescent microscopy. Apoptotic cells (%) are shown as the mean ± S.E. of three independent experiments. **P<0.01, significantly different from the control. n.s., not significant. Transfection efficiency was measured by counting DsRed monomer-expressing cells, and calculated to be 16.5 ± 0.4% (the mean ± S.E of three independent experiments).
(TIF)
(A and B) Bcl-rambo and Bcl-rambo (ΔTM) were expressed using GMR-GAL4 driver fly lines. (A) (a, d, g) GMR-GAL4/w; +; UAS-Bcl-rambo/+ (strain #1), (b, e, h) GMR-GAL4/UAS-Bcl-rambo; +; + (strain #6), (c, f, i) GMR-GAL4/w; UAS-Bcl-rambo/+; + (strain #46) (B) (a, d, g) GMR-GAL4/w; UAS-Bcl-rambo (ΔTM)/+; + (strain #10), (b, e, h) GMR-GAL4/w; UAS-Bcl-rambo (ΔTM)/+; + (strain #26), (c, f, i) GMR-GAL4/UAS-Bcl-rambo (ΔTM); +; + (strain #148) The morphology of adult eyes was observed by light microscopy (a–c) and SEM (d–i). Scale bars in d–f and g–i indicate 50 μm and 14.2 μm, respectively. Data were representative of two independent experiments.
(TIF)
(A and B) Bcl-rambo and Bcl-rambo (ΔTM) were expressed using GMR-GAL4 driver fly lines. (A) (a, e, i) GMR-GAL4/+; UAS-Bcl-rambo/UAS-GFP; +, (b, f, j) GMR-GAL4/w; UAS-Bcl-rambo/+; UAS-p35/+, (c, g, k) GMR-GAL4/w; UAS-Bcl-rambo/+; UAS-Diap1/+, (d, h, l) GMR-GAL4/w; UAS-Bcl-rambo/+; GMR-Diap2/+. (B) (a, e, i) GMR-GAL4/+; UAS-Bcl-rambo (ΔTM)/UAS-GFP; +, (b, f, j) GMR-GAL4/w; UAS-Bcl-rambo (ΔTM)/+; UAS-p35/+, (c, g, k) GMR-GAL4/w; UAS-Bcl-rambo (ΔTM)/+; UAS-Diap1/+, (d, h, l) GMR-GAL4/w; UAS-Bcl-rambo (ΔTM)/+; GMR-Diap2/+. The morphology of adult eyes was observed by light microscopy (a–d) and SEM (e–l). Scale bars in e–h and i–l indicate 50 μm and 14.2 μm, respectively. Data were representative of two independent experiments.
(TIF)
Bcl-rambo and Bcl-rambo (ΔTM) were expressed using GMR-GAL4 driver fly lines. (A) (a, e, i) GMR-GAL4/w; UAS-Bcl-rambo/+; +, (b, f, j) GMR-GAL4/w; UAS-Bcl-rambo/BuffyH37; +, (c, g, k) GMR-GAL4/w; UAS-Bcl-rambo (ΔTM)/+; +, (d, h, l) GMR-GAL4/w; UAS-Bcl-rambo (ΔTM)/ BuffyH37; +. The morphology of adult eyes was observed by light microscopy (a–d) and SEM (e–l). Scale bars in e–h and i–l indicate 50 μm and 14.2 μm, respectively. Data were representative of two independent experiments.
(TIF)
Acknowledgments
We are very grateful to Ms. Akane Kyotani for technical assistance. This work was partly supported by a research grant from the NOVARTIS Foundation (Japan) for the Promotion of Science (to T.K.) and Grants-in-Aid from the Ministry of Education, Science, Sports and Culture of Japan, Japan Science and Technology Agency and JSPS Core-to-Core Program, B. Asia-Africa Science Platforms.
Data Availability
All relevant data are within the paper and its Supporting Information files.
Funding Statement
This work was partly supported by a research grant from the NOVARTIS Foundation (Japan) for the Promotion of Science (to TK) and Grants-in-Aid from the Ministry of Education, Science, Sports and Culture of Japan, Japan Science and Technology Agency and JSPS Core-to-Core Program, B. Asia-Africa Science Platforms. The funders had no role in study design, data collection and analysis, decision to publish, or preparation of the manuscript.
References
- 1.Fuchs Y, Steller H (2011) Programmed cell death in animal development and disease. Cell 147: 742–758. 10.1016/j.cell.2011.10.033 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 2.Nikoletopoulou V, Markaki M, Palikaras K, Tavernarakis N (2013) Crosstalk between apoptosis, necrosis and autophagy. Biochim Biophys Acta 1833: 3448–3459. 10.1016/j.bbamcr.2013.06.001 [DOI] [PubMed] [Google Scholar]
- 3.Siegel RM (2006) Caspases at the crossroads of immune-cell life and death. Nat Rev Immunol 6: 308–317. [DOI] [PubMed] [Google Scholar]
- 4.Bratton SB, Salvesen GS (2010) Regulation of the Apaf-1-caspase-9 apoptosome. J Cell Sci 123: 3209–3214. 10.1242/jcs.073643 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 5.Lavrik IN, Krammer PH (2012) Regulation of CD95/Fas signaling at the DISC. Cell Death Differ 19: 36–41. 10.1038/cdd.2011.155 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 6.Riedl SJ, Shi Y (2004) Molecular mechanisms of caspase regulation during apoptosis. Nat Rev Mol Cell Biol 5: 897–907. [DOI] [PubMed] [Google Scholar]
- 7.Newmeyer DD, Ferguson-Miller S (2003) Mitochondria: releasing power for life and unleashing the machineries of death. Cell 112: 481–490. [DOI] [PubMed] [Google Scholar]
- 8.Youle RJ, Strasser A (2008) The BCL-2 protein family: opposing activities that mediate cell death. Nat Rev Mol Cell Biol 9: 47–59. [DOI] [PubMed] [Google Scholar]
- 9.Martinou JC, Youle RJ (2011) Mitochondria in apoptosis: Bcl-2 family members and mitochondrial dynamics. Dev Cell 21: 92–101. 10.1016/j.devcel.2011.06.017 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 10.Chipuk JE, Bouchier-Hayes L, Green DR (2006) Mitochondrial outer membrane permeabilization during apoptosis: the innocent bystander scenario. Cell Death Differ 13: 1396–1402. [DOI] [PubMed] [Google Scholar]
- 11.Garrido C, Galluzzi L, Brunet M, Puig PE, Didelot C, Kroemer G (2006) Mechanisms of cytochrome c release from mitochondria. Cell Death Differ 13: 1423–1433. [DOI] [PubMed] [Google Scholar]
- 12.Kataoka T, Holler N, Micheau O, Martinon F, Tinel A, Hofmann K, et al. (2001) Bcl-rambo, a novel Bcl-2 homologue that induces apoptosis via its unique C-terminal extension. J Biol Chem 276: 19548–19554. [DOI] [PubMed] [Google Scholar]
- 13.Holleman A, den Boer ML, de Menezes RX, Cheok MH, Cheng C, Kazemier KM, et al. (2006) The expression of 70 apoptosis genes in relation to lineage, genetic subtype, cellular drug resistance, and outcome in childhood acute lymphoblastic leukemia. Blood 107: 769–776. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 14.Brill E, Gobble R, Angeles C, Lagos-Quintana M, Crago A, Laxa B, et al. (2010) ZIC1 overexpression is oncogenic in liposarcoma. Cancer Res 70: 6891–6901. 10.1158/0008-5472.CAN-10-0745 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 15.Yang YL, Lin SR, Chen JS, Lin SW, Yu SL, Chen HY, et al. (2010) Expression and prognostic significance of the apoptotic genes BCL2L13, Livin, and CASP8AP2 in childhood acute lymphoblastic leukemia. Leuk Res 34: 18–23. 10.1016/j.leukres.2009.07.023 [DOI] [PubMed] [Google Scholar]
- 16.Wang Z, Yan Z, Zhang B, Rao Z, Zhang Y, Liu J, et al. (2013) Identification of a 5-gene signature for clinical and prognostic prediction in gastric cancer patients upon microarray data. Med Oncol 30: 678 10.1007/s12032-013-0678-5 [DOI] [PubMed] [Google Scholar]
- 17.Jensen SA, Calvert AE, Volpert G, Kouri FM, Hurley LA, Luciano JP, et al. (2014) Bcl2L13 is a ceramide synthase inhibitor in glioblastoma. Proc Natl Acad Sci USA 111: 5682–5687. 10.1073/pnas.1316700111 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 18.Boumela I, Assou S, Haouzi D, Déchaud H, Aït-Ahmed O, Hamamah S (2014) Develpmental regulated expression of anti- and pro-apoptotic BCL-2 family genes during human early embryonic development. Curr Med Chem 21: 1361–1369. [DOI] [PubMed] [Google Scholar]
- 19.Yi P, Zhang W, Zhai Z, Miao L, Wang Y, Wu M (2003) Bcl-rambo beta, a special splicing variant with an insertion of an Alu-like cassette, promotes etoposide- and Taxol-induced cell death. FEBS Lett 534: 61–68. [DOI] [PubMed] [Google Scholar]
- 20.Banga S, Gao P, Shen X, Fiscus V, Zong WX, Chen L, et al. (2007) Legionella pneumophila inhibits macrophage apoptosis by targeting pro-death members of the Bcl2 protein family. Proc Natl Acad Sci USA 104: 5121–5126. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 21.Kim JY, So KJ, Lee S, Park JH (2012) Bcl-rambo induces apoptosis via interaction with the adenine nucleotide translocator. FEBS Lett 586: 3142–3149. 10.1016/j.febslet.2012.08.015 [DOI] [PubMed] [Google Scholar]
- 22.Schouten M, Fratantoni SA, Hubens CJ, Piersma SR, Pham TV, Bielefeld P, et al. (2015) MicroRNA-124 and -137 cooperatively controls caspase-3 activity through BCL2L13 in hippocampal neural stem cells. Sci Rep 5:12448 10.1038/srep12448 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 23.Murakawa T, Yamaguchi O, Hashimoto A, Hikoso S, Takeda T, Oka T, et al. (2015) Bcl-2-like protein 13 is a mammalian Atg32 homologue that mediates mitophagy and mitochondrial fragmentation. Nat Commun 6:7527 10.1038/ncomms8527 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 24.Kornbluth S, White K (2005) Apoptosis in Drosophila: neither fish nor fowl (nor man, nor worm). J Cell Sci 118: 1779–1787. [DOI] [PubMed] [Google Scholar]
- 25.Steller H (2008) Regulation of apoptosis in Drosophila. Cell Death Differ 15: 1132–1138. 10.1038/cdd.2008.50 [DOI] [PubMed] [Google Scholar]
- 26.Denton D, Aung-Htut MT, Kumar S (2013) Developmentally programmed cell death in Drosophila. Biochim Biophys Acta 1833: 3499–3506. 10.1016/j.bbamcr.2013.06.014 [DOI] [PubMed] [Google Scholar]
- 27.Kumar S, Doumanis J (2000) The fly caspases. Cell Death Differ 7: 1039–1044. [DOI] [PubMed] [Google Scholar]
- 28.Darding M, Meier P (2012) IAPs: guardians of RIPK1. Cell Death Differ. 19: 58–66. 10.1038/cdd.2011.163 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 29.Danial NN, Korsmeyer SJ (2004) Cell death: critical control points. Cell 116: 205–219. [DOI] [PubMed] [Google Scholar]
- 30.Spradling AC (1986) P-element-mediated transformation In: Drosophila: A practical approach (ed Robert D.B.), IRL Press, Oxford, pp. 175–197. [Google Scholar]
- 31.Takahashi Y, Hirose F, Matsukage A, Yamaguchi M (1999) Identification of three conserved regions in the DREF transcription factors from Drosophila melanogaster and Drosophila virilis. Nucleic Acids Res. 27: 510–516. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 32.Brachmann CB, Jassim OW, Wachsmuth BD, Cagan RL (2000) The Drosophila Bcl-2 family member dBorg-1 functions in the apoptotic response to UV-irradiation. Curr Biol 10: 547–550. [DOI] [PubMed] [Google Scholar]
- 33.Colussi PA, Quinn LM, Huang DCS, Coombe M, Read SH, Richardson H, et al. (2000) Debcl, a proapoptotic Bcl-2 homologue, is a component of the Drosophila melanogaster cell death machinery. J Cell Biol 148: 703–714. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 34.Igaki T, Kanuka H, Inohara N, Sawamoto K, Núñez G, Okano H, et al. (2000) Drob-1, a Drosophila member of the Bcl-2/CED-9 family that promotes cell death. Proc Natl Acad Sci USA 97: 662–667. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 35.Zhang H, Huang Q, Ke N, Matsuyama S, Hammock B, Godzik A, et al. (2000) Drosophila pro-apoptotic Bcl-2/Bax homologue reveals evolutionary conservation of cell death mechanisms. J Biol Chem 275: 27303–27306. [DOI] [PubMed] [Google Scholar]
- 36.Moriyama S, Maves L, Chen Y, Hoffmann FM. (1996) decapentaplegic overexpression affects Drosophila wing and leg imaginal disc development and wingless expression. Dev Biol 177:136–151. [DOI] [PubMed] [Google Scholar]
- 37.Tanimoto H, Itoh S, ten Dijke P, Tabata T (2000) Hedgehog creates a gradient of DPP activity in Drosophila wing imaginal discs. Mol Cell 5:59–71. [DOI] [PubMed] [Google Scholar]
- 38.Heitzler P, Haenlin M, Ramain P, Calleja M, Simpson P (1996) A genetic analysis of pannier, a gene necessary for viability of dorsal tissues and bristle positioning in Drosophila. Genetics 143:1271–1286. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 39.Calleja M, Herranz H, Estella C, Casal J, Lawrence P, Simpson P, et al. (2000) Generation of medial and lateral dorsal body domains by the pannier gene of Drosophila. Development 127:3971–3980. [DOI] [PubMed] [Google Scholar]
- 40.Fraser AG, McCarthy NJ, Evan GI (1997) drICE is an essential caspase required for apoptotic activity in Drosophila cells. EMBO J 16: 6192–6199. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 41.Song Z, Guan B, Bergman A, Nicholson DW, Thornberry NA, Peterson EP, et al. (2000) Biochemical and genetic interactions between Drosophila caspases and the proapoptotic genes rpr, hid, and grim. Mol Cell Biol 20: 2907–2914. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 42.Callus BA, Vaux DL (2007) Caspase inhibitors: viral, cellular and chemical. Cell Death Differ 14: 73–78. [DOI] [PubMed] [Google Scholar]
- 43.Kaufmann T, Schlipf S, Sanz J, Neubert K, Stein R, Borner C (2003) Characterization of the signal that directs Bcl-xL, but not Bcl-2, to the mitochondrial outer membrane. J Cell Biol 160: 53–64. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 44.Doumanis J, Dorstyn L, Kumar S (2007) Molecular determinants of the subcellular localization of the Drosophila Bcl-2 homologues DEBCL and BUFFY. Cell Death Differ 14: 907–915. [DOI] [PubMed] [Google Scholar]
- 45.Sevrioukov EA, Burr J, Huang EW, Assi HH, Monserrate JP, Purves DC, et al. (2007) Drosophila Bcl-2 proteins participate in stress-induced apoptosis, but are not required for normal development. Genesis 45: 184–193. [DOI] [PubMed] [Google Scholar]
- 46.Quinn L, Coombe M, Mills K, Daish T, Colussi P, Kumar S, et al. (2003) Buffy, a Drosophila Bcl-2 protein, has anti-apoptotic and cell cycle inhibitory functions. EMBO J 22: 3568–3579. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 47.Senoo-Matsuda N, Igaki T, Miura M (2005) Bax-like protein Drob-1 protects neurons from expanded polyglutamine-induced toxicity in Drosophila. EMBO J 24: 2700–2713. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 48.Gaumer S, Guénal I, Brun S, Théodore L, Mignotte B (2000) Bcl-2 and Bax mammalian regulators of apoptosis are functional in Drosophila. Cell Death Differ 7: 804–814. [DOI] [PubMed] [Google Scholar]
- 49.Brun S, Rincheval V, Gaumer S, Mignotte B, Guenal I (2002) reaper and bax initiate two different apoptotic pathways affecting mitochondria and antagonized by bcl-2 in Drosophila. Oncogene 21: 6458–6470. [DOI] [PubMed] [Google Scholar]
- 50.Galindo KA, Lu WJ, Park JH, Abrams JM (2009) The Bax/Bak ortholog in Drosophila, Debcl, exerts limited control over programmed cell death. Development 136: 275–283. 10.1242/dev.019042 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 51.Zhivotovsky B, Galluzzi L, Kepp O, Kroemer G (2009) Adenine nucleotide translocase: a component of the phylogenetically conserved cell death machinery. Cell Death Differ 16: 1419–1425. 10.1038/cdd.2009.118 [DOI] [PubMed] [Google Scholar]
- 52.Vartiainen S, Chen S, George J, Tuomela T, Luoto KR, O’Dell KMC, et al. (2014) Phenotypic rescue of a Drosophila model of mitochondrial ANT1 disease. Dis Model Mech 7: 635–648. 10.1242/dmm.016527 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 53.Kraut R (2011) Roles of sphingolipids in Drosophila development and disease. J Neurochem 116: 764–778. 10.1111/j.1471-4159.2010.07022.x [DOI] [PubMed] [Google Scholar]
- 54.Mullen TD, Hannun YA, Obeid LM (2012) Ceramide synthases at the centre of sphingolipid metabolism and biology. Biochem J 441: 789–802. 10.1042/BJ20111626 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 55.Bauer R, Voelzmann A, Breiden B, Schepers U, Farwanah H, Hahn I, et al. (2009) Schlank, a member of the ceramide synthase family controls growth and body fat in Drosophila. EMBO J 28: 3706–3716. 10.1038/emboj.2009.305 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 56.Ryoo HD, Baehrecke EH (2010) Distinct death mechanisms in Drosophila development. Curr Opin Cell Biol 22: 889–895. 10.1016/j.ceb.2010.08.022 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 57.Jenkins VK, Timmons AK, McCall K (2013) Diversity of cell death pathways: insight from the fly ovary. Trends Cell Biol 23: 567–574. 10.1016/j.tcb.2013.07.005 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 58.Yacobi-Sharon K, Namdar Y, Arama E (2013) Alternative germ cell death pathway in Drosophila involves HtrA2/Omi, lysosomes, and a caspase-9 counterpart. Dev Cell 25: 29–42. 10.1016/j.devcel.2013.02.002 [DOI] [PubMed] [Google Scholar]
Associated Data
This section collects any data citations, data availability statements, or supplementary materials included in this article.
Supplementary Materials
(A) Structures of human Bcl-rambo and its mutants. (B) S2 cells were transfected with pMT-V5-His A, pMT-V5-His A/Bcl-rambo (1–441), or pMT-V5-His A/Bcl-rambo (1–459) and then incubated in the presence of CuSO4 (500 μM) and Z-VAD-fmk (20 μM) for 24 h. S2 cells were stained for Bcl-rambo (green) and with DAPI (blue) and MitoTracker® Red (red). The stained cells in at least five different fields were observed by confocal laser scanning microscopy. Optical sections containing a single transfected cell are shown. Data were representative of two independent experiments. Scale bars indicate 10 μm.
(TIF)
(A) S2 cells were transfected with pAct5C-GAL4 together with pMT-V5-His A, pMT-V5-His A/Bcl-rambo, pMT-V5-His A/Bcl-rambo (ΔTM), pMT-V5-His A/Bcl-rambo (1–441), pMT-V5-His A/Bcl-rambo (1–459), or pUAST-DsRed-monomer in the presence of CuSO4 (500 μM) for 24 h. Cells were stained with Hoechst 33342. Nuclear morphology was observed by fluorescent microscopy. Apoptotic cells (%) are shown as the mean ± S.E. of three independent experiments. **P<0.01, significantly different from the control. n.s., not significant. Transfection efficiency was measured by counting DsRed monomer-expressing cells, and calculated to be 16.5 ± 0.4% (the mean ± S.E of three independent experiments).
(TIF)
(A and B) Bcl-rambo and Bcl-rambo (ΔTM) were expressed using GMR-GAL4 driver fly lines. (A) (a, d, g) GMR-GAL4/w; +; UAS-Bcl-rambo/+ (strain #1), (b, e, h) GMR-GAL4/UAS-Bcl-rambo; +; + (strain #6), (c, f, i) GMR-GAL4/w; UAS-Bcl-rambo/+; + (strain #46) (B) (a, d, g) GMR-GAL4/w; UAS-Bcl-rambo (ΔTM)/+; + (strain #10), (b, e, h) GMR-GAL4/w; UAS-Bcl-rambo (ΔTM)/+; + (strain #26), (c, f, i) GMR-GAL4/UAS-Bcl-rambo (ΔTM); +; + (strain #148) The morphology of adult eyes was observed by light microscopy (a–c) and SEM (d–i). Scale bars in d–f and g–i indicate 50 μm and 14.2 μm, respectively. Data were representative of two independent experiments.
(TIF)
(A and B) Bcl-rambo and Bcl-rambo (ΔTM) were expressed using GMR-GAL4 driver fly lines. (A) (a, e, i) GMR-GAL4/+; UAS-Bcl-rambo/UAS-GFP; +, (b, f, j) GMR-GAL4/w; UAS-Bcl-rambo/+; UAS-p35/+, (c, g, k) GMR-GAL4/w; UAS-Bcl-rambo/+; UAS-Diap1/+, (d, h, l) GMR-GAL4/w; UAS-Bcl-rambo/+; GMR-Diap2/+. (B) (a, e, i) GMR-GAL4/+; UAS-Bcl-rambo (ΔTM)/UAS-GFP; +, (b, f, j) GMR-GAL4/w; UAS-Bcl-rambo (ΔTM)/+; UAS-p35/+, (c, g, k) GMR-GAL4/w; UAS-Bcl-rambo (ΔTM)/+; UAS-Diap1/+, (d, h, l) GMR-GAL4/w; UAS-Bcl-rambo (ΔTM)/+; GMR-Diap2/+. The morphology of adult eyes was observed by light microscopy (a–d) and SEM (e–l). Scale bars in e–h and i–l indicate 50 μm and 14.2 μm, respectively. Data were representative of two independent experiments.
(TIF)
Bcl-rambo and Bcl-rambo (ΔTM) were expressed using GMR-GAL4 driver fly lines. (A) (a, e, i) GMR-GAL4/w; UAS-Bcl-rambo/+; +, (b, f, j) GMR-GAL4/w; UAS-Bcl-rambo/BuffyH37; +, (c, g, k) GMR-GAL4/w; UAS-Bcl-rambo (ΔTM)/+; +, (d, h, l) GMR-GAL4/w; UAS-Bcl-rambo (ΔTM)/ BuffyH37; +. The morphology of adult eyes was observed by light microscopy (a–d) and SEM (e–l). Scale bars in e–h and i–l indicate 50 μm and 14.2 μm, respectively. Data were representative of two independent experiments.
(TIF)
Data Availability Statement
All relevant data are within the paper and its Supporting Information files.