
Figure 7. Model of how bridging of synaptic vesicles to the plasma membrane by the highly conserved C-terminal region of Munc13s can create a cage-like environment and facilitate the activity of the MUN domain in promoting the transition from the syntaxin-1-Munc18-1 complex to the SNARE complex, thus favoring SNARE complex assembly.
Syntaxin-1 (Habc domain, orange; SNARE motif and N-terminus, yellow) is shown in a closed conformation bound to Munc18-1 (purple). Synaptobrevin is shown in red, SNAP-25 in green and the C-terminal region of Munc13-1 in brown. The model is inspired by the ability of C1C2BMUNC2C to bridge V- to T-liposomes (Figure 6) and assumes that the C1-C2B region binds to the plasma membrane while the C2C domain binds to the vesicle membrane. See text and the legend of Figure 7—figure supplement 1 for additional details.

Figure 7—figure supplement 1. Speculative models of membrane bridging by C1C2BMUN and C1C2BMUNC2C.
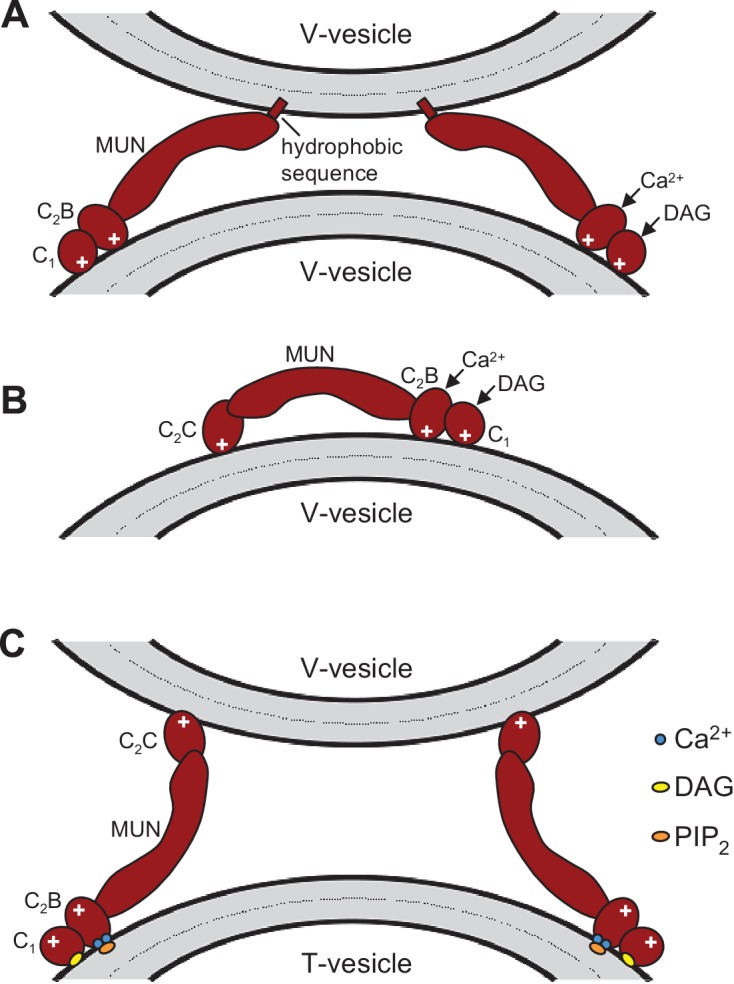
These models serve in part as a basis for the model proposed in Figure 7 and provide a rationalization for the liposomes clustering activities observed for C1C2BMUN and C1C2BMUNC2C. However, it is important to note that there are multiple potential explanations for these activities. The findings that PS is a major determinant of vesicle clustering by C1C2BMUN (Figure 2) without requiring Ca2+ (Figure 2—figure supplement 1), but C1C2BMUNC2C requires Ca2+ to cluster V-liposomes (Figure 6A,B), suggest that there are multiple membrane binding sites in these large protein fragments that can cooperate in cis to interact with a single membrane or in trans to bind to two membranes. Indeed, the MUN, C1 and C2B domains contain several positive patches, the C1 domain binds to DAG, and the C2B domain binds to PIP2 weakly in the absence of Ca2+ and more strongly in the presence of Ca2+ (Shen et al., 2005; Shin et al., 2010; Yang et al., 2015). The C2C domain is likely to have at least one lipid-binding site with moderate affinity that explains the stronger overall liposome clustering activity of C1C2BMUNC2C compared to C1C2BMUN (Figure 6—figure supplement 1) (see discussion). Moreover, the sequence spanning residues 1517–1531 at the C-terminus of C1C2BMUN does not form part of the MUN domain structure (Yang et al. 2015) and contains a highly hydrophobic sequence that could bind to membranes, but this sequence may become structured due to the presence of the C2C domain in C1C2BMUNC2C, which could render it unable to bind membranes. We speculate that this hydrophobic sequence together with positive patches in the C1-C2B region underlie the liposome clustering activity of Ca2+-free C1C2BMUN (A), while in C1C2BMUNC2C the C2C domain provides a PS-binding site that cooperates with the C1-C2B region to favor binding in cis to the same membrane (B). Ca2+ binding to the C2B domain may favor membrane binding of C1C2BMUNC2C in a different orientation that facilitates interaction of the C2C domain in trans with another membrane, which would explain why Ca2+-bound C1C2BMUNC2C can bridge V-liposomes; this orientation could also be favored by binding of the C2B domain to PIP2 and of the C1 domain to DAG in T-liposomes (C), leading to the overall notion that the C1-C2B region binds to the plasma membrane and the C2C domain to synaptic vesicle membrane (Figure 7). Extensive studies will be required to test this and other plausible models compatible with the liposome clustering data.