

Abstract
Previous work has shown that wing wear increases mortality in bumblebees. Although a proximate mechanism for this phenomenon has remained elusive, a leading hypothesis is that wing wear increases predation risk by reducing flight manoeuvrability. We tested the effects of simulated wing wear on flight manoeuvrability in Bombus impatiens bumblebees using a dynamic obstacle course designed to push bees towards their performance limits. We found that removing 22% wing area from the tips of both forewings (symmetric wear) caused a 9% reduction in peak acceleration during manoeuvring flight, while performing the same manipulation on only one wing (asymmetric wear) did not significantly reduce maximum acceleration. The rate at which bees collided with obstacles was correlated with body length across all treatments, but wing wear did not increase collision rate, possibly because shorter wingspans allow more room for bees to manoeuvre. This study presents a novel method for exploring extreme flight manoeuvres in flying insects, eliciting peak accelerations that exceed those measured during flight through a stationary obstacle course. If escape from aerial predation is constrained by acceleration capacity, then our results offer a potential explanation for the observed increase in bumblebee mortality with wing wear.
Keywords: insect flight, bumblebees, wing damage, manoeuvrability, collisions
1. Introduction
Inadvertent collisions with obstacles are inevitable for many flying insects, especially pollinators that regularly weave through vegetation clutter. Insect wings are particularly vulnerable to collision damage. In foraging bumblebees, wing collisions occur once per second on average, and collision frequency is correlated with wing area loss [1], which increases mortality rate [2]. Surprisingly, a direct mechanistic link between wing wear and mortality has not yet been discovered. Simulated wing wear does not increase the energetic cost of flight in bumblebees [3], and has little effect on their foraging flight performance, including maximum acceleration and mean velocity [4]. However, previous studies have shown that wing area loss reduces maximum vertical aerodynamic force production in bumblebees [5–7], and it has been suggested that vertical force capacity can more generally predict an animal's ability to accelerate in any direction during flight manoeuvres [5,7,8].
We test the hypothesis that wing wear reduces bumblebee flight manoeuvrability, a performance trait that may constrain a bee's ability to escape from aerial predators or parasitoids. Combes et al. [9] addressed a similar question in dragonflies (from the perspective of the predator rather than the prey), and found that dragonflies with experimentally induced wing wear displayed lower flight accelerations, escape velocities and predation success, compared with those with intact wings. Although natural predator–prey experiments are a powerful method for examining extreme flight performance, such an assay would be difficult to implement with bumblebees and any of their natural predators. Instead, we challenged bumblebees to fly through a dynamic obstacle.
Although it is commonly assumed that higher acceleration capacity should improve flight performance in cluttered environments [10,11], a recent study showed that bumblebees flying through a static obstacle course do not regularly operate near their acceleration limits; rather, collision-avoidance behaviour appears to be a primary determinant of flight performance in clutter [10]. Motivated by these findings, we built a moving obstacle course—analogous to plants swaying in wind—designed to push bees closer to their maximum acceleration limits. We measured two performance metrics that have been used to infer flight manoeuvrability in previous studies: maximum aerodynamic acceleration [8] and the rate of collisions with obstacles [12].
2. Material and methods
(a). Experimental set-up
We cold-anaesthetized a colony of bumblebees (Bombus impatiens), randomly separated them into three treatment groups, and marked each individual with a paint colour according to group. In the symmetric group (SYM), we clipped the tips of both forewings to simulate natural wing wear, using a procedure similar to prior wing wear studies [3,6,7]. In the asymmetric group (ASYM), we clipped only the right forewing. In the intact group (INT), we left both forewings intact (figure 1a–c). The clipping procedure removed approximately 22% of the forewing area, which is within the range of naturally occurring damage [2]. The non-dimensional second moment of area for an intact wing was 0.59, and for a clipped wing was 0.57.
Figure 1.

Bees were randomly assigned to a treatment group in which (a) the wings were left intact (INT), (b) 22% of the right forewing was removed (ASYM) or (c) 22% of both forewings were removed (SYM). (d) Representative flight trajectory (see supplementary movies). (e) Time trace of two-dimensional (horizontal) and three-dimensional acceleration magnitude. Regions in grey were excluded from the analysis. (Online version in colour.)
The hive was placed in an indoor chamber that connected to the outdoors at the far end of a rectangular tunnel. Inside the tunnel was an array of 21 vertical posts (6.4 mm diameter, 6.35 cm spacing) mounted on an orbital shaker, which was oscillated at 2 Hz frequency and 2.5 cm amplitude. Foragers had to fly through the moving obstacle course en route to the outdoors and back (figure 1d).
Flight sequences were filmed with two synchronized Photron SA3 cameras (1024 × 1024 pixel resolution) sampling at 500 fps, one positioned above the tunnel and the other positioned 30° from vertical, viewing down the long axis of the tunnel. Only sequences that exceeded 1 s of continuous flight within the obstacle course were analysed.
(b). Kinematic analysis
We digitized centre of body mass kinematics using DLTdv5 [13], and processed the data with a fifth-order Butterworth low-pass filter with a cut-off of 15 Hz. In three frames of each video, we also digitized the anterior tip of the head and the posterior tip of the abdomen to calculate an average body length for each individual.
Most trials featured at least one collision between a bee and a post. We quantified the collision rate by counting the number of collisions observed in each video and dividing by the total flight time. Because collisions often generated extreme body accelerations, we removed every segment of data that encompassed a collision event, including at least 30 ms before and after contact. We also removed any segments where the bee's body was obstructed from the top camera view by a post. We calculated flight velocity and acceleration by numerically differentiating the remaining trajectory data, and found the mean speed and maximum aerodynamic acceleration for each trial (figure 1e). We focused on the two-dimensional components of acceleration in the horizontal plane, because our dynamic obstacle course was designed to elicit sharp lateral manoeuvres. See supplementary material for additional details on the kinematic analysis.
(c). Statistical analysis
Statistical analyses were performed in R [14]. To determine how wing treatment and body length were associated with maximum two-dimensional acceleration, we used a multiple regression, first using maximum two-dimensional acceleration as a response to treatment, body length and the interaction of these covariates. We selected a final model by using a series of likelihood ratio tests to remove non-significant covariates (using α = 0.05 as the significance level). To find which treatments were different from the others, we used simultaneous tests for general linear hypotheses, using the glht function from the R package multcomp [15]. To determine how treatment and body length were associated with collision rate, we used a Poisson generalized linear model (using the glm function) with a natural log link function. We used the number of collisions as a response variable, and included flight time in the model as an offset. As above, we used a series of likelihood ratio tests to check for significant interactions and to remove non-significant terms. We conducted a similar analysis for mean flight speed through the obstacle course. See supplementary material for additional details on the statistical analysis.
3. Results
Wing treatment affected maximum two-dimensional acceleration (F2,142 = 5.07; p-value = 0.0075). Post hoc comparisons revealed that bees with symmetric wing wear produced significantly lower peak accelerations than those with intact wings (t142 = 3.17; p-value = 0.0019), but that peak two-dimensional accelerations did not differ significantly between the asymmetrically trimmed group and either of the other groups (figure 2a). An analysis of maximum three-dimensional accelerations yielded qualitatively similar results. Maximum two-dimensional acceleration was not associated with body length (figure 2b). We found no significant difference in collision rate among treatments (figure 2c), but did find a significant positive association between collision rate and body length (GLM: influence of body length, 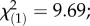 p-value = 0.0019, figure 2d). We found no differences in mean flight velocity through the obstacle course among the treatment groups (F2,142 = 0.19; p-value = 0.83). See supplementary material for additional information on the results of the statistical analysis.
p-value = 0.0019, figure 2d). We found no differences in mean flight velocity through the obstacle course among the treatment groups (F2,142 = 0.19; p-value = 0.83). See supplementary material for additional information on the results of the statistical analysis.
Figure 2.
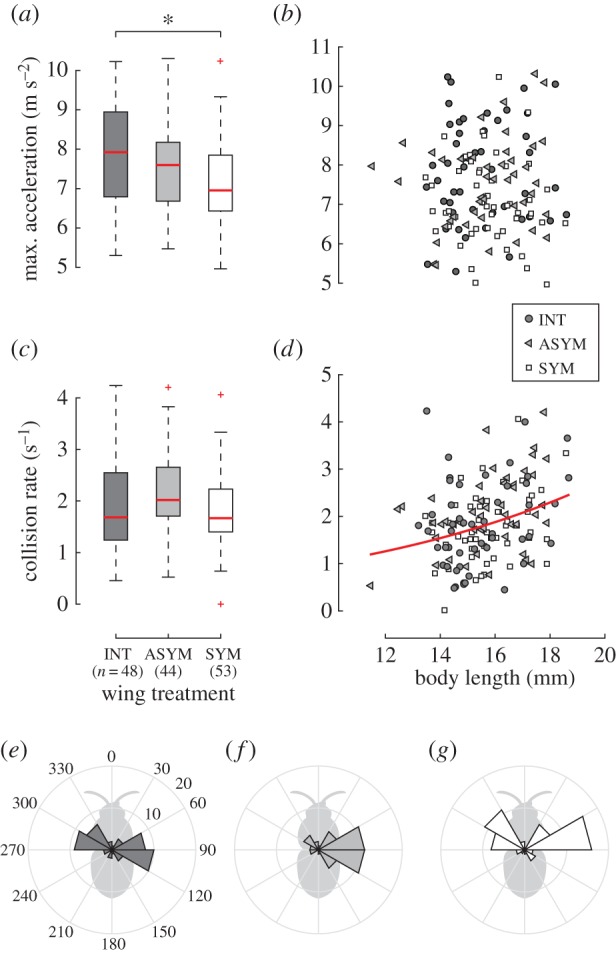
Maximum two-dimensional acceleration (a,b) and collision rate (c,d). Box and whisker plots show the median, quartiles and range of data points, with outliers plotted as plus signs. Asterisk indicates a significant difference. Line in (d) shows the predicted collision rate from the generalized linear model. (e–g) Angle histograms of maximum two-dimensional acceleration relative to body orientation for each treatment. (Online version in colour.)
4. Discussion
Clipping both wing tips significantly reduced maximum acceleration by 9% on average (figure 2a). Clipping only one wing tip also reduced maximum acceleration, but not significantly. These results are qualitatively consistent with a recent study showing that wing area loss was negatively correlated with load-lifting capacity in bumblebees, whereas asymmetric damage had no additional effect beyond the overall area loss [6].
Peak accelerations were primarily oriented laterally relative to the bee's body axis (figure 2e–g), consistent with roll-based flight manoeuvring. Although bees in the INT and SYM groups displayed a roughly equal frequency of left- and right-directed peak accelerations, the effect of clipping only the right wing in the ASYM group caused a dramatic reduction in the number of left-directed peak accelerations, indicating that asymmetric wear may have asymmetric effects on manoeuvrability.
Our results demonstrate that a moving obstacle course can push bees closer to their acceleration capacity. Peak three-dimensional accelerations for bees in our INT group exceeded those of B. impatiens bumblebees flying through a static obstacle course [10] by 0.7 m s−2, or 13% on average. Still, we do not know whether the peak accelerations measured here represent absolute maximum performance, and if not, whether bees of all sizes were under-performing to a similar degree. The lack of an association between peak acceleration and body length (figure 2b) would appear to argue against maximal performance across all body sizes, given that vertical aerodynamic force production has been found to decline with body size in bumblebees [5] and orchid bees [16]. While it is possible that vertical force capacity might not adequately predict maximum acceleration in three-dimensions, it is more likely that our assay was pushing larger bees closer to their acceleration capacity than smaller bees, which would be consistent with the body-size-dependent collision rate.
Although bees in the SYM group displayed significantly lower maximum accelerations, this did not increase their collision rate with posts (figure 2c). On the other hand, larger bees did collide with posts more frequently than smaller bees across all treatments (figure 2d), as has been seen in both bumblebees and bats flying through fixed obstacle courses [10,12]. Taken together, these results suggest that collision rate may be mediated by a combination of maximal acceleration and the spatial footprint of the bee (i.e. the volume of the body and the region swept by both wings). Small bees collide with posts less frequently because they have more room in which to enact directional changes. Similarly, bees with clipped wings have more room to manoeuvre owing to a shorter wingspan, perhaps compensating for their reduced accelerations.
The ability to accelerate is a critical component of manoeuvrability for flying insects, with potentially life-threatening consequences. This study presents an experimental assay that capitalizes on bumblebees' inherent collision-avoidance behaviour to push them closer to their flight performance limits. We show that wing damage decreases accelerations during manoeuvring flight, which may help explain why wing-worn bumblebees experience greater mortality. Future studies focused on bumblebee flight responses to aerial predators will clarify whether acceleration capacity does in fact limit escape ability.
Supplementary Material
Ethics
This work complies with ethical guidelines in the USA.
Data accessibility
Supplementary methods, results and movies are included in the electronic supplementary material. The dataset supporting this article is available on Dataverse: http://dx.doi.org/10.7910/DVN/4ZOIZL.
Authors' contributions
A.M.M. conceived and designed the experiment, collected and analysed the data and drafted the manuscript; T.M.A. participated in the experimental design and data collection; C.M.S. carried out the statistical analyses; S.A.C. helped analyse and interpret the data. T.M.A., C.M.S. and S.A.C. revised the paper critically for important intellectual content. All authors gave final approval of the manuscript for publication and acknowledge joint accountability for its content.
Competing interests
We have no competing interests.
Funding
This work was supported by CCF-0926148 and IOS-1253677.
References
- 1.Foster DJ, Cartar RV. 2011. What causes wing wear in foraging bumble bees? J. Exp. Biol. 214, 1896–1901. ( 10.1242/jeb.051730) [DOI] [PubMed] [Google Scholar]
- 2.Cartar RV. 1992. Morphological senescence and longevity: an experiment relating wing wear and life span in foraging wild bumble bees. J. Anim. Ecol. 61, 225–231. ( 10.2307/5525) [DOI] [Google Scholar]
- 3.Hedenstrom A, Ellington CP, Wolf TJ. 2001. Wing wear, aerodynamics and flight energetics in bumblebees (Bombus terrestris): an experimental study. Funct. Ecol. 15, 417–422. ( 10.1046/j.0269-8463.2001.00531.x) [DOI] [Google Scholar]
- 4.Haas CA, Cartar RV. 2008. Robust flight performance of bumble bees with artificially induced wing wear. Can. J. Zool. 86, 668–675. ( 10.1139/Z08-034) [DOI] [Google Scholar]
- 5.Buchwald R, Dudley R. 2010. Limits to vertical force and power production in bumblebees (Hymenoptera: Bombus impatiens). J. Exp. Biol. 213, 426–432. ( 10.1242/jeb.033563) [DOI] [PubMed] [Google Scholar]
- 6.Johnson SA, Cartar RV. 2014. Wing wear, but not asymmetry in wear, affects load-lifting capability in bumble bees Bombus impatiens. Can. J. Zool. 92, 179–184. ( 10.1139/cjz-2013-0229) [DOI] [Google Scholar]
- 7.Vance JT, Roberts SP. 2014. The effects of artificial wing wear on the flight capacity of the honey bee Apis mellifera. J. Insect Physiol. 65, 27–36. ( 10.1016/j.jinsphys.2014.04.003) [DOI] [PubMed] [Google Scholar]
- 8.Dudley R. 2002. Mechanisms and implications of animal flight maneuverability. Integr. Comp. Biol. 42, 135–140. ( 10.1093/icb/42.1.135) [DOI] [PubMed] [Google Scholar]
- 9.Combes SA, Crall JD, Mukherjee S. 2010. Dynamics of animal movement in an ecological context: dragonfly wing damage reduces flight performance and predation success. Biol. Lett. 6, 426–429. ( 10.1098/rsbl.2009.0915) [DOI] [PMC free article] [PubMed] [Google Scholar]
- 10.Crall JD, Ravi S, Mountcastle AM, Combes SA. 2015. Bumblebee flight performance in cluttered environments: effects of obstacle orientation, body size and acceleration. J. Exp. Biol. 218, 2728–2737. ( 10.1242/jeb.121293) [DOI] [PubMed] [Google Scholar]
- 11.Norberg UM, Rayner JMV. 1987. Ecological morphology and flight in bats (Mammalia; Chiroptera): wing adaptations, flight performance, foraging strategy and echolocation. Phil. Trans. R. Soc. Lond. B 316, 335–427. ( 10.1098/rstb.1987.0030) [DOI] [Google Scholar]
- 12.Stockwell EF. 2001. Morphology and flight manoeuvrability in New World leaf-nosed bats (Chiroptera: Phyllostomidae). J. Zool. 254, 505–514. ( 10.1017/S0952836901001005) [DOI] [Google Scholar]
- 13.Hedrick TL. 2008. Software techniques for two- and three-dimensional kinematic measurements of biological and biomimetic systems. Bioinspir. Biomim. 3, 034001. ( 10.1088/1748-3182/3/3/034001) [DOI] [PubMed] [Google Scholar]
- 14.R Core Team. 2016. R: A language and environment for statistical computing. Vienna, Austria: R Foundation for Statistical Computing. https://www.R-project.org/ (accessed 10 March 2016). [Google Scholar]
- 15.Hothorn T, Bretz F, Westfall P. 2008. Simultaneous inference in general parametric models. Biomet. J. 50, 346–363. ( 10.1002/bimj.200810425) [DOI] [PubMed] [Google Scholar]
- 16.Dillon ME, Dudley R. 2004. Allometry of maximum vertical force production during hovering flight of neotropical orchid bees (Apidae: Euglossini). J. Exp. Biol. 207, 417–425. ( 10.1242/jeb.00777) [DOI] [PubMed] [Google Scholar]
Associated Data
This section collects any data citations, data availability statements, or supplementary materials included in this article.
Supplementary Materials
Data Availability Statement
Supplementary methods, results and movies are included in the electronic supplementary material. The dataset supporting this article is available on Dataverse: http://dx.doi.org/10.7910/DVN/4ZOIZL.