

Abstract
The new model presented here can be used to understand the influence of hemodynamics on specific cardiac developmental processes, at the cellular and molecular level. To alter intracardiac hemodynamics, fertilized chicken eggs are incubated in a humidified chamber to obtain embryos of the desired stage (HH17). Once this developmental stage is achieved, the embryo is maintained ex ovo and hemodynamics in the embryonic heart are altered by partially constricting the outflow tract (OFT) with a surgical suture at the junction of the OFT and ventricle (OVJ). Control embryos are also cultured ex ovo but are not subjected to the surgical intervention. Banded and control embryos are then incubated in a humidified incubator for the desired period of time, after which 2D ultrasound is employed to analyze the change in blood flow velocity at the OVJ as a result of OFT banding. Once embryos are maintained ex ovo, it is important to ensure adequate hydration in the incubation chamber so as to prevent drying and eventually embryo death. Using this new banded model, it is now possible to perform analyses of changes in the expression of key players involved in valve development and to understand the role of hemodynamics on cellular responses in vivo, which could not be achieved previously.
Keywords: Developmental Biology, Issue 111, Hemodynamics, chicken, outflow tract, embryonic heart, banding, ex ovo, shell-less chick embryo culture
Introduction
Abnormally formed outflow valves are the most common type of congenital heart defects 1. However, defective cardiac valve structure and function, even though present at birth, may become symptomatic only in adulthood. In fact, several adult valve diseases can be attributed to a congenital origin. Treatment of such patients often involves replacing defective valves, and, importantly, replaced aortic valves have been shown to have congenital anomalies 2. Given the fact that critical processes involved in valve development begin early during embryogenesis, the importance of better understanding the mechanisms that regulate these events is highlighted.
The primitive heart tube, which is the first functioning organ in an embryo, exhibits two distinct layers - an endothelial endocardium surrounded by myocardium - separated by extracellular matrix (cardiac jelly) which is mostly produced and secreted by the myocardium 3-5. As development continues, valve primordia (endocardial cushions) are formed, after rightward looping of the embryonic heart, by local expansion of the cardiac jelly at the atrioventricular (AV) canal and the outflow tract (OFT) 4,6. This expansion is mediated by the highly regulated process of epithelial-mesenchymal transition (EMT), during which the cardiac jelly becomes populated by endocardially-derived mesenchymal cells 6. In addition to the mesenchymal population derived through EMT, neural crest cells are also involved in valvulogenesis of the OFT 3.
Hemodynamic stimuli, such as shear stress, are important epigenetic factors that regulate heart development in the embryo 7,8. Using a 3D in vitro system, we have previously shown shear stress to be a factor influencing the expression and deposition of fibrous extracellular matrix (ECM) proteins in AV and OFT cushions 9,10. Moreover, studies carried out by several researchers have demonstrated that altered blood flow leads to improper valves and septa formation 11-16. Recently, using the novel banding procedure presented here, we have shown that changing hemodynamics in the embryonic chick heart affects the early processes involved in OFT valve formation 17.
The technique described here provides a novel model for altering hemodynamics in the developing chick heart by partially constricting the OFT ex ovo. This reproducible procedure is relatively quick and allows researchers to obtain a sufficient number of embryos/whole hearts/OFT tissue, etc. for downstream analyses including gene expression studies. Moreover, this new model can be used to study 'chronic' effects of altered hemodynamics on OFT valve development.
Protocol
Avian embryos are not considered vertebrate animals under IUCAC regulations.
1. Obtaining Embryos for Surgery
Incubate 80 - 90 fertilized Bovan chicken eggs (blunt end up) in a humidified (60%) rocker incubator at 40 °C for approximately 72 hr to obtain embryos at Hamilton and Hamburger (HH) stage 17. Determine exact number of eggs based on power analysis and survival rate of embryos 17. Use plastic egg trays for incubation to allow adequate air circulation.
Once this desired developmental stage is attained, pour approximately 20 ml of warm (37 °C) Tyrode's buffer (supplemented with sodium bicarbonate (1 g/L)) into a 100 mm x 26 mm petri dish.
Sterilize an egg shell with 70% ethanol.
Gently crack the shell with a scalpel handle and carefully release its contents into the dish containing Tyrode's buffer. Discard embryos if (i) they visually appear abnormal (ii) they are not at the right developmental stage (iii) they are improperly oriented on the yolk and/or (iv) any bleeding occurs. Retain any embryos not subjected to surgery immediately after being placed ex ovo at 40 °C so as to not compromise development. Note: Staging is performed based on Hamilton and Hamburger 18. Abnormality of chick embryos is determined by the naked eye and under the dissection scope. Ensure that the embryo has appropriate folding and bending and vasculature.
2. OFT Banding
Tweeze out individual 1 cm long threads from a single 11/0 nylon surgical suture and form a loose knot. UV sterilize all preformed knots.
Under a dissection microscope, visually ensure that the heart is beating at a normal rate (~ 120 beats/min). If not, discard the embryo.
Just before surgery, apply 5 - 6 ml of warm (37 °C) Tyrode's buffer to the embryo surface using a transfer pipette.
Pass one free end of the preformed knot under the OFT, position the suture at the OFT/ventricle junction (OVJ) (or desired location), and pass the free end into the knot thereby constricting the OFT including the membranes surrounding the heart.
After surgery, wet the surface of the yolk with 5 - 6 ml of warm (37 °C) Tyrode's buffer using a transfer pipette.
Maintain control embryos ex ovo as the banded embryos; however do not subject them to the surgical procedure.
Incubate the embryos, for the desired amount of time, at 40 °C in a humidified (60%) incubator.
Once the desired time point is reached, excise the embryo from the yolk using straight scissors. Dissect the heart with fine forceps and use for several downstream studies such as changes in cardiac morphology due to altered hemodynamics 17.
3. Confirming that the Banding Intervention Causes a Change in Hemodynamics
Note: The partial constriction caused by the banding intervention leads to an increase in blood flow velocity at the OVJ. This hemodynamic parameter is conveniently assessed using 2D ultrasound imaging, which is performed at the desired time point of the experiment.
Since only a single embryo can be imaged at a time, maintain all other embryos designated for ultrasound imaging in a 40 °C incubator.
Place the petri dish containing the embryo to be imaged on a heating pad set at 40 °C.
Fill the dish, up to the brim, with warm (37 °C) Tyrode's buffer. Slowly pour the buffer solution into the dish so as to keep the yolk intact. If, however, the integrity of the yolk is compromised during this step, discard the embryo.
Orient the embryo such that the central axis of the embryo is perpendicular to the ultrasound probe which is suspended from an adjustable stand.
With the ultrasound machine operating in B-mode, obtain a 2D image of the beating heart on the screen and move the stage (heating pad) such that the OFT, ventricle and OVJ are clearly visible.
- Switch to the pulsed-wave (PW) mode, at the desired pulse repetition frequency (e.g., 20 kHz) and obtain velocity data exactly at the OVJ. A B-mode image of the beating heart is obtained on the screen.
- Move the stage to see the OFT, ventricle and the OVJ. Using the ultrasound machine software, obtain velocity measurements exactly at the OVJ.
Note: If the heart rate of an embryo decreases during imaging, velocity data thus obtained should not be used for analysis. All embryos used for velocity measurements should preferably not be used for any other experiments after ultrasound imaging.
Representative Results
Shown in Figure 1 are recommended instruments needed for OFT banding. The petri dish (Figure 1A) containing the embryo ex ovo should be deep enough so as to not destroy the embryo when covered with the lid. Deep petri dishes (Figure 1C) should also be used for ultrasound imaging to allow an adequate volume of Tyrode's buffer to be poured atop the yolk.
A single 1 cm thread (Figure 2A) is tweezed out from an 11 / 0 nylon suture (Figure 1A) and tied into a loose knot (Figure 1B). All preformed knots are UV sterilized. Figure 3A shows a humidified rocker chamber where fertilized chicken eggs are incubated until the desired stage. Figure 3B shows an embryo incubator, where banded and control embryos are maintained ex ovo for the desired amount of time. Both incubator types are maintained at 40 °C.
Shown in Figure 4A is a HH17 embryo on top of the yolk ex ovo. Figure 4B is a schematic representation showing the position of the suture (band) at the OVJ of a HH17 embryo. Figure 4C is a representative image of a banded embryo imaged 48 hr after surgery.
The effect of the banding intervention on blood flow velocity through the constriction site is measured by 2D ultrasound imaging. As expected, the velocity at the OVJ is higher in the banded embryo relative to the control (Figure 5). We have recently performed computational fluid dynamics (CFD) analysis on banded and control hearts at the 24 hr time point showing the effects of OFT banding on blood flow velocity and wall shear stress 17.
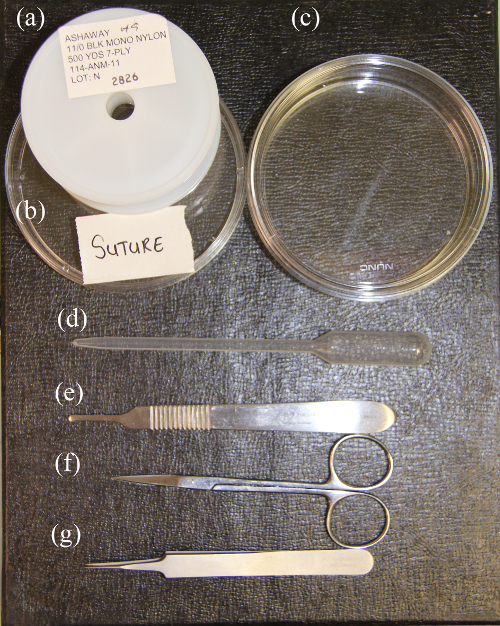 Figure 1. Surgical Instruments.(A) 11/0 nylon suture. (B) Petri plate containing pre-formed knots. (C) Deep petri dish for ex ovo embryo culture and ultrasound imaging. (D) Transfer pipette. (E) Scalpel handle to crack the egg shell. (F) Straight scissors for embryo excision for post-surgery studies. (G) Fine forceps Please click here to view a larger version of this figure.
Figure 1. Surgical Instruments.(A) 11/0 nylon suture. (B) Petri plate containing pre-formed knots. (C) Deep petri dish for ex ovo embryo culture and ultrasound imaging. (D) Transfer pipette. (E) Scalpel handle to crack the egg shell. (F) Straight scissors for embryo excision for post-surgery studies. (G) Fine forceps Please click here to view a larger version of this figure.
 Figure 2. Suture for OFT Banding. (A) A single 1 cm thread tweezed out from a suture. (B) Pre-formed knot. Please click here to view a larger version of this figure.
Figure 2. Suture for OFT Banding. (A) A single 1 cm thread tweezed out from a suture. (B) Pre-formed knot. Please click here to view a larger version of this figure.
 Figure 3. Incubation Chambers. (A) Humidified rocker incubator for fertilized eggs. (B) Humidified chamber for ex ovo culture of banded and control embryos. Please click here to view a larger version of this figure.
Figure 3. Incubation Chambers. (A) Humidified rocker incubator for fertilized eggs. (B) Humidified chamber for ex ovo culture of banded and control embryos. Please click here to view a larger version of this figure.
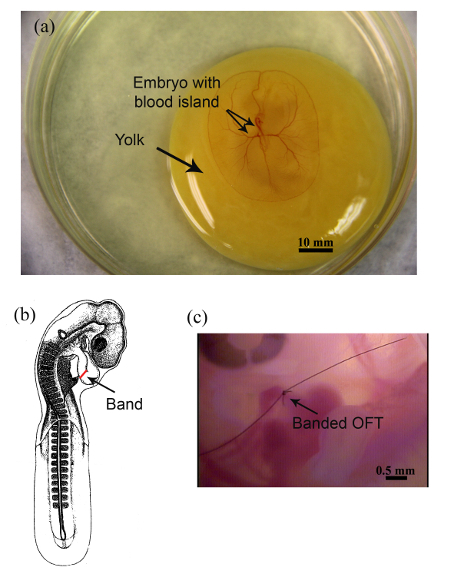 Figure 4. OFT Banding. (A) Whole HH17 chick embryo maintained ex ovo, scale bar = 10 mm (B) Schematic representation depicting the constriction site (red). (C) Banded embryo imaged 48 hr after OFT banding, scale bar = 0.5 mm. Please click here to view a larger version of this figure.
Figure 4. OFT Banding. (A) Whole HH17 chick embryo maintained ex ovo, scale bar = 10 mm (B) Schematic representation depicting the constriction site (red). (C) Banded embryo imaged 48 hr after OFT banding, scale bar = 0.5 mm. Please click here to view a larger version of this figure.
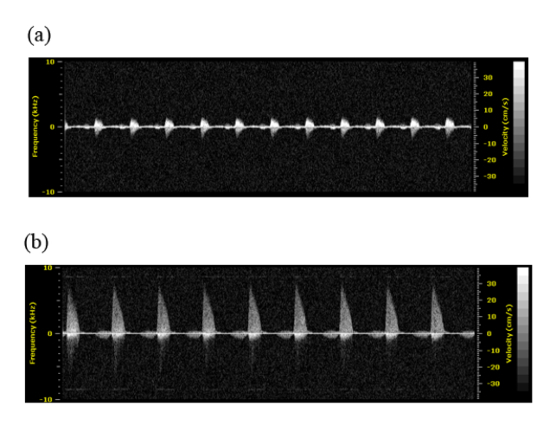 Figure 5. Ultrasound-derived Velocity Measurements. Representative velocity profiles of (A) control and (B) banded embryos obtained at a 1 hr time point. Please click here to view a larger version of this figure.
Figure 5. Ultrasound-derived Velocity Measurements. Representative velocity profiles of (A) control and (B) banded embryos obtained at a 1 hr time point. Please click here to view a larger version of this figure.
Discussion
This technique is relatively quick and easy to perform, however certain key points need to be kept in mind so as to get accurate downstream results. Embryos should be maintained ex ovo in a petri dish that contains Tyrode's buffer to provide adequate rehydration. It is also important to hydrate the yolk post-surgery with Tyrode's buffer and to make sure the incubation chamber is adequately hydrated. Surgeries should not be performed on an embryo if any bleeding is visible or the yolk is even slightly broken. This affects embryo survival, especially for long-term experiments. Another potential cause of lethality to embryos, is whether the band around the OFT is too tight when first applied, thus significantly compromising cardiac function.
At present, we use ultrasound as a confirmatory tool to determine the change in blood flow velocity upon OFT banding. In the future, we will confirm using micro-computed tomography (CT). We also visually determine the band tightness around the OFT in B-mode 2D images. Importantly, no differences in heart rate and volumetric flow (computed using CFD) are observed between embryos in the banded and control groups 17. These indicate that the physiology of the cardiovascular system is not compromised in the banded embryos, thus further validating the banding intervention.
At the desired time point, after banding, several analyses can be performed to understand the effect of altered hemodynamics on critical valve developmental processes. These include, but are not limited to, determining changes in transcript levels of key regulatory genes involved in valve development, determining localization of ECM components and determining the ultrastructure of the OFT.
Finally, this technique improves on the current method of banding the OFT in the embryo inside the windowed egg 19. That method is severely hampered by the inability to accumulate necessary embryos to do downstream molecular analyses. This new technique can be used in rescue experiments as well. This application allows for the banded OFT to be rescued and embryo allowed to recover from the insult. No current method can do this and still be able to perform statistically significant downstream analyses.
We have successfully employed this technique to change the hemodynamic stimuli in the heart of chick embryos at a very early developmental stage (HH 16 - 17). The banding protocol might present a challenge when performing the surgery at later stages of development due to the risk of nicking any of the extraembryonic blood vessels. Also, in late stages of development, it is often difficult to keep the yolk sac intact when the egg contents are transferred into the petri dish.
Disclosures
The authors have nothing to disclose.
Acknowledgments
The authors would like to acknowledge Dr. Robert Price and the staff of the Instrumentation Resource Facility at the University of South Carolina School of Medicine. This work was partially supported by a SPARC Graduate Research Grant from the Office of the Vice President for Research at the University of South Carolina (JDP/VM). In addition this work was supported by Cook Biotech research agreement (JDP) and by FirstString Research Inc (JDP) and NIH 2 P20-RR016434-06 (JDP). In addition, NIH IDeA Networks of Biomedical Research Excellence (INBRE) grant for South Carolina P20GM103499 (JE)
References
- Neeb Z, Lajiness J, Bolanis E, Conway S. Cardiac outflow tract anomalies. Wiley Interdiscip Rev Dev Biol. 2013;2(4):499–530. doi: 10.1002/wdev.98. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Combs M, Yutzey K. Heart valve development: regulatory networks in development and disease. Circ Res. 2009;105(5):408–421. doi: 10.1161/CIRCRESAHA.109.201566. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Hinton R, Yutzey K. Heart valve structure and function in development and disease. Annu Rev Physiol. 2011;73:29–46. doi: 10.1146/annurev-physiol-012110-142145. [DOI] [PMC free article] [PubMed] [Google Scholar]
- von Gise A, Pu W. Endocardial and epicardial epithelial to mesenchymal transitions in heart development and disease. Circ Res. 2012;110(12):1628–1645. doi: 10.1161/CIRCRESAHA.111.259960. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Person A, Klewer S, Runyan R. Cell biology of cardiac cushion development. Int Rev Cytol. 2005;243:287–335. doi: 10.1016/S0074-7696(05)43005-3. [DOI] [PubMed] [Google Scholar]
- de Vlaming A, et al. Atrioventricular valve development: new perspectives on an old theme. Differentiation. 2012;84(1):103–116. doi: 10.1016/j.diff.2012.04.001. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Butcher J, McQuinn T, Sedmera D, Turner D, Markwald R. Transitions in early embryonic atrioventricular valvular function correspond with changes in cushion biomechanics that are predictable by tissue composition. Circ Res. 2007;100(10):1503–1511. doi: 10.1161/CIRCRESAHA.107.148684. [DOI] [PubMed] [Google Scholar]
- Hove J, et al. Intracardiac fluid forces are an essential epigenetic factor for embryonic cardiogenesis. Nature. 2003;421(6919):172–177. doi: 10.1038/nature01282. [DOI] [PubMed] [Google Scholar]
- Tan H, et al. Fluid flow forces and rhoA regulate fibrous development of the atrioventricular valves. Dev Biol. 2013;374(2):345–356. doi: 10.1016/j.ydbio.2012.11.023. [DOI] [PubMed] [Google Scholar]
- Biechler S, et al. The impact of flow-induced forces on the morphogenesis of the outflow tract. Front Physiol. 2014;5 doi: 10.3389/fphys.2014.00225. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Hu N, Clark E. Hemodynamics of the stage 12 to stage 29 chick embryo. Circ Res. 1989;65(6):1665–1670. doi: 10.1161/01.res.65.6.1665. [DOI] [PubMed] [Google Scholar]
- Hogers B, DeRuiter M, Gittenberger-de Groot A, Poelmann R. Unilateral vitelline vein ligation alters intracardiac blood flow patterns and morphogenesis in the chick embryo. Circ Res. 1997;80(4):473–481. doi: 10.1161/01.res.80.4.473. [DOI] [PubMed] [Google Scholar]
- Hogers B, DeRuiter M, Gittenberger-de Groot A, Poelmann R. Extraembryonic venous obstructions lead to cardiovascular malformations and can be embryolethal. Cardiovasc Res. 1999;41(1):87–99. doi: 10.1016/s0008-6363(98)00218-1. [DOI] [PubMed] [Google Scholar]
- Reckova M, et al. Hemodynamics is a key epigenetic factor in development of the cardiac conduction system. Circ Res. 2003;93(1):77–85. doi: 10.1161/01.RES.0000079488.91342.B7. [DOI] [PubMed] [Google Scholar]
- Stekelenburg-de Vos S, et al. Acutely altered hemodynamics following venous obstruction in the early chick embryo. J Exp Biol. 2003;206(pt 6):1051–1057. doi: 10.1242/jeb.00216. [DOI] [PubMed] [Google Scholar]
- Lucitti J, Tobita K, Keller B. Arterial hemodynamics and mechanical properties after circulatory intervention in the chick embryo. J Exp Biol. 2005;208(pt 10):1877–1885. doi: 10.1242/jeb.01574. [DOI] [PubMed] [Google Scholar]
- Menon V, Eberth J, Goodwin R, Potts J. Altered Hemodynamics in the Embryonic Heart Affects Outflow Valve Development. J. Cardiovasc. Dev. Dis. 2015;2(2):108–124. doi: 10.3390/jcdd2020108. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Hamburger V, Hamilton H. A series of normal stages in the development of the chick embryo. J. Morphol. 1951;88:473–481. [PubMed] [Google Scholar]
- Midgett M, Goenezen S, Rugonyi S. Blood flow dynamics reflect degree of outflow tract banding in Hamburger-Hamilton stage 18 chicken embryos. J R Soc Interface. 2014;11(100) doi: 10.1098/rsif.2014.0643. [DOI] [PMC free article] [PubMed] [Google Scholar]