
Figure 3.
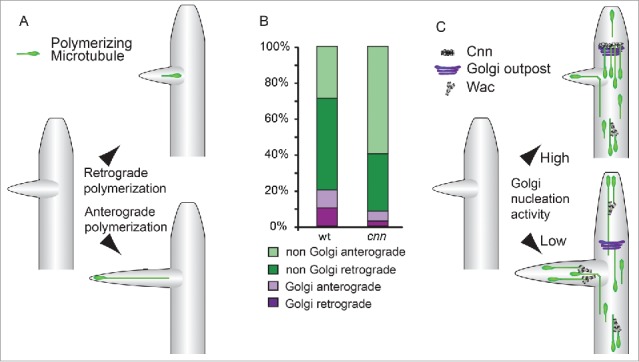
(A) Anterograde polymerizing microtubule invasion into nascent dendrite branches drives outgrowth. Microtubules nucleated within termini or at the branch site are more likely to drive an extension event.35 At a lower frequency, similar to the invasion of microtubules into the spines of mature dendrites,88 both retrograde- or anterograde-oriented microtubules in the main branch enter nascent dendrite branches, becoming anterograde-directed within the termini.35 (B) The relative contributions of Golgi- and non-Golgi-derived polymerizing microtubules to the dendrite termini. Golgi outpost-associated events contribute approximately equally to both anterograde and retrograde polymerizing microtubules. In cnn mutant neurons, there is an increase in non-Golgi microtubules that polymerize in the anterograde direction; this increase is not accounted for by the loss of Golgi outpost-associated retrograde polymerization events. These data suggest that nucleation machinery at outposts might counteract the activity of additional factors that promote anterograde bias. (C) A model for how arbor complexity can be regulated by modulating the balance between these distinct Golgi- and non-Golgi-associated pathways that orient microtubule polymerization. A single outpost is shown here; it should be noted, however, that throughout the arbor individual Golgi outposts can give rise to either repeated anterograde or retrograde nucleation events. During dendrite outgrowth, Cnn recruits microtubule nucleation to Golgi outposts to suppress an activity (that may be due to Augmin, here illustrated by the Drosophila Augmin component Wac62), which in turn promotes anterograde polymerization.