

ABSTRACT
Many host- and vaccine-specific factors modulate an antibody response. Host genetic polymorphisms, in particular, modulate the immune response in multiple ways on different scales. This review article describes how information on host genetic polymorphisms and corresponding immune cascades may be used to generate personalized vaccine strategies to optimize the antibody response.
KEYWORDS: vaccine, genetic polymorphism, SNP, individualized vaccination, GWAS, immune response, antibodies, Influenza, Measles, systems biology
Introduction
In the last century, vaccines have become the primary prevention strategy of many infectious diseases and have saved millions of lives.1 On the other hand, the development of effective vaccines against global killers such as human immunodeficiency virus, Mycobacterium tuberculosis, and malaria parasites (Plasmodium spp.) remains a major challenge: Just recently, the European Medicines Agency gave a positive scientific opinion on the first malaria vaccine Mosquirix (RTS,S), although the vaccine's efficacy is limited.2 Existing vaccines for Mycobacterium tuberculosis are only partially effective.3 Also, vaccines against influenza or measles may fail in some cases. In particular, patients at the extremes of age,4 pregnant women,5 as well as patients with chronic diseases such as diabetes,6 autoimmune diseases,7,8 or after transplantation9-11 show lower vaccine response rates. A better understanding of the host-pathogen interaction and new insights into vaccine immune response modulating factors will help to improve current vaccination strategies and to develop novel types of vaccines.12,13
Vaccination effectiveness is influenced by various vaccine-, pathogen-, and host-related factors (see Fig. 1).4,14-16 Several studies have demonstrated that the host genetic background (genotype) has a strong influence on the immune response, e.g. to influenza, Hepatitis B or measles vaccination.17-19 Over the last few years, it has been proposed frequently that genetic information might be used to predict vaccine effectiveness and might help to develop more effective, individualized vaccination strategies.15,20,21 In this review, we summarize general concepts of how the genetic variations of the host can contribute to variability of vaccine-induced humoral immunity. Further, we discuss important clinical studies and how mathematical, mechanistic models help uncover therapeutic targets for personalized vaccination strategies.
Figure 1.
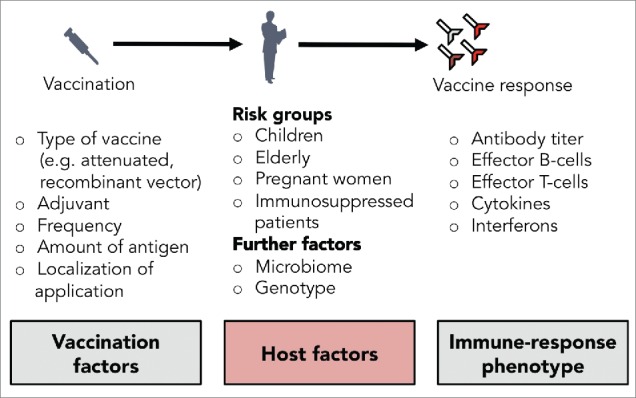
Selected factors for a successful vaccination.
The Vaccine Induced Immune Response: A Network of Networks
In general, vaccines aim to induce a significant level of neutralizing antibodies against specific viruses or bacteria, leading to protective immunity. In clinical studies seroprotection is normally defined as a specific antibody titer or antibody titer increase (seroconversion).22 However, vaccine-induced immunity is far more complex and can be understood as a dynamic network of molecular, cell-to-cell and tissue interactions that are organized in a hierarchical structure. Molecular interactions form networks and are themselves organized in networks on a higher cellular level: Dendritic cells detect the vaccine antigen and other components through pattern-recognition receptors, in particular Toll-like receptors (TLRs).23 Activated TLRs in turn initiate various signaling pathways through Toll/interleukin-1 receptor(TIR)-domain containing adaptor proteins such as myeloid differentiation primary response protein 88 (MyD88) and TIR domain containing adapter molecule 1 (TICAM1). This results in the expression of inflammatory genes, which are mainly regulated by a specific interferon regulatory factor or nuclear factor-κB (NF-κB), leading to a production of various cell surface receptors, cytokines, and chemokines.24 Activated dendritic cells mature to antigen-presenting cells and migrate to lymph nodes, where they present vaccine epitopes to T-cell receptors through human leukocyte antigen (HLA) molecules on their cell surface.25 This in turn initiates the maturation of naïve CD4+ T-cells to effector T-cells. The additional secretion of cytokines such as interleukin (IL) 12 and interferon (IFN) γ leads to proliferation of type 1 T helper cells (Th1), while the secretion of cytokines such as IL-4, IL-6, and IL-10 leads to proliferation of type 2 T helper cells (Th2). Th2 cells in turn support the proliferation of B-cells and their differentiation to antibody-secreting plasma cells and are thus an important factor for a successful vaccination.26-28 Due to its complexity, the vaccine-induced immune response is a focus of on-going research and further processes involved in the humoral immune response have been reported.29,30
Immunological network structure and robustness
Basically, intercellular signaling processes of immune cells are orchestrated by cytokines, chemokines, and cell surface receptors, while intracellular signaling processes are conducted by various signaling pathways (e.g., TLR or Janus Kinase (JAK)/Signal transducer and activator of transcription (STAT) signaling pathway). Gene regulatory networks control both intra- and intercellular processes. As often described for biological interaction networks,31-33 one can also assume that in an immune response only few components such as NF-κB regulate many processes (so-called key regulators) while most components regulate only a few processes. An advantage of such a network topology is that a single mutation in a random gene hardly affects the global immune response, because the failure does not propagate.34 In addition, gene redundancy, overlapping functions of genes as well as regulatory feedback mechanisms are able to compensate for gene perturbations. The concept underlying these phenomena is known as biological robustness and is a key property of living systems.35,36 The immune system shows features of robust systems such as functional redundancy of genes and proteins.37-39 For example, a mutation in IFN-α1 (IFNA1) may affect its binding affinity to a receptor, but it has many paralogues, which are themselves potent alpha interferons (IFN-α2-14).37 On the other hand, mutations in the NF-κB signaling pathway or several mutations in HLA molecules has been linked to diseases such as Crohn's disease and other autoimmune disorders.40-42 We assume that in immunosuppressed patients, e.g. transplant recipients, the level of robustness is reduced due to immunosuppressive drugs, which decreases the compensatory mechanisms. Immunosuppressive drugs used in transplant patients mainly affect different signals of T-cell activation, e.g., Calcineurin inhibitors, as well as proliferation capacities of T- and B-cells such as mycophenolic acid, rapamycin or methotrexate.43 Other immunosuppressive drugs affect signaling pathways such as JAK/STAT or TNF-α inhibitors.44 Therefore in such risk groups, genetic polymorphisms affecting the vaccine outcome may be easier to unmask.
Genetic polymorphisms in vaccine response
Evolution acts on many levels
The immune response is continuously shaped by evolutionary adaptations on the genome level of both host and pathogen, although with different rates. The development of vaccines against pathogens with dynamic antigen variation (e.g., human immunodeficiency virus (HIV) and other RNA viruses) remains a major challenge.45 Knowledge of genetically stable regions in the pathogen genome helps to develop new vaccination strategies.46 Similarly, the knowledge of genetic variations in the host, which lead to an increased antibody response, may help to predict vaccine efficiency and uncover important factors of the immune response. For instance, HLA molecules show high genetic variation among individuals (more than 13,000 alleles in the IMGT/HLA database, release 3.21.0) and HLA polymorphisms have been used to reconstruct human migration events.47 Due to the key role of HLA in the self/non-self immune recognition, it is not surprising that vaccine response shows variability between individuals.
Single nucleotide polymorphisms (SNPs)
SNPs are single nucleotide variations of the genome sequence, which occur frequently within a population (by definition at least 1%).48 In genetic studies, particular SNPs, so-called tag SNPs, are used as markers for haplotypes. Haplotypes are regions with high linkage disequilibrium and are represented by a set of SNPs.49 To create a haplotype map of the considered genome, genotyping studies usually only need to examine a set of characteristic SNPs instead of sequencing the whole genome.50
Due to technical advances in genotyping techniques,51 the number of submitted SNPs in the NCBI dbSNP database has grown exponentially over the last 10 y. The human genome has about 3 billion base pairs.52 Early studies estimated an average SNP frequency within the human genome of about one SNP per 1000 base pairs (corresponds to 3 million SNPs).53,54 Today, there are almost 98 million SNPs listed in dbSNP (release build 144). However, the majority of SNPs must be regarded as candidate SNPs as the results of the studies that originally discovered them have not been reproduced. Some studies highlighted that a significant amount of SNPs in the dbSNP database is not reliable.55,56 In comparison, the SNP map provided by the international HapMap project reported about 10 million SNPs with a minor allele frequency (MAF) of at least 5%.57 Most SNPs, however, probably do not impact the phenotype due to biological robustness (as mentioned above).
In general, SNPs can be located within coding or non-coding sequences58: Within the coding region, a SNP may change the amino acid sequence of the respective protein (missense SNP), result in a stop-codon (nonsense SNP) or just have no effect on the protein sequence (synonymous SNP). SNPs within the non-coding region can affect the produced protein amount on the DNA or RNA level: At the DNA level, SNPs in the 5′ untranslated region (UTR) may affect the transcription factor binding, which leads to an up or down regulation of gene expression. SNPs in the 3′-UTR may affect microRNA binding and, as a result, gene silencing. At the RNA level, SNPs may affect mRNA degradation, RNA splicing, or the RNA sequence of non-coding RNA.
Impact of SNPs on the humoral immune response
Presuming an evolutionary pressure exerted by host-pathogen interaction, one can hypothesize that SNPs either have none, a positive or a negative, but never a fatal effect on the host. In fact, several independent studies identified a couple of SNPs to influence treatment outcome of infectious disease such as influenza, Hepatitis C virus (HCV), and cytomegalovirus (CMV) as well as the antibody response after vaccination (e.g., influenza, measles).19,59-62
A very prominent example concerns IFN-λ.63,64 Since 2009, SNPs in IFNL3/4 genes are associated with the HCV treatment outcome.60,61,65-70 In particular, SNPs in IFNL3 (rs8099917) and IFNL4 (rs368234815) have been proposed as predictors for spontaneous viral clearance and treatment success to pegylated interferon α/ribavirin (PEG-IFN-α/RBV) treatment.60,61 The IFNL4 SNP (rs368234815) has also been associated with CMV retinitis in HIV-infected risk patients (n=1134, p-value=7E-3),71 as well as with CMV replication in solid-organ transplant recipients at risk (n=455, p-value=4E-02).72 Moreover, an IFNL3 SNP (rs8099917) may modulate the humoral immune response after vaccination.62 Individuals carrying the minor allele in one or both alleles showed an increased seroconversion rate after influenza vaccination.62 Quantitative real-time PCR investigations have shown, that IFNL3 expression in individuals carrying the minor allele in one or both alleles was lower in PBMCs.70 In vitro studies showed that IFNL3 suppressesTh2-cytokines74-77 and modulates B-cell function.62,78 Although the impact of IFN-λ on immune cells is not yet understood, these studies indicate that IFNL3 is an important regulator of the Th1/Th2 balance and modulates viral clearance/antibody response.
Further SNPs that influence the humoral immune response to influenza vaccination have been reported (see Table 1): Gelder et al. investigated associations between HLA class II alleles and H1N1/H3N2 hemagglutination-inhibition (HAI) titers in an influenza risk group. They identified 4 alleles (n = 73, p-value range of 2.3E-03 to 1.6E-02, significance level p-value < 5.0E-2).79 Poland et al. were not able to reproduce the HLA class II associations to H1N1/H3N2 antibody titer (likely due to insufficient statistical power). Instead, they found association of HLA class I alleles and H1N1 antibody titer, as well as several SNPs in coding and non-coding regions of cytokines and cytokine receptors (n = 184, p-value range of 2.3E-03 to 9.0E-02, significance level p-value < 5.0E-2).80 Franco et al. combined genotype, gene expression and antibody titer information in order to identify genes whose genotype influences the antibody response through an alteration of gene expression. They identified 20 genes (n = 199, p-value < 5.0E-08, significance level p-value < 5.0E-2).17 Most of the identified genes are not specifically linked to the immune system, but to intracellular transport and membrane trafficking. Seven genes encode proteins involved in antigen transport and antigen processing, but these findings have to be confirmed through further studies with larger sample size.20
Table 1.
Genes with polymorphisms that influence the vaccine induced antibody level (selected studies).
| Vaccine | Gene(s) | Function | Study | n | Remarks | Reference |
|---|---|---|---|---|---|---|
| HBV | CD11a (ITGAL) | part of LFA-1 | 43 candidate SNPs across 133 genes1 | 662 | reproduced in second group (n=393) | Hennig et al. 2008 |
| HLA-DR, HLA-DP | HLA class II molecules | GWAS2 | 1683 | reproduced in second group (n=1931) | Png et al. 2011 | |
| HLA Class III | GWAS2 | 1683 | reproduced in second group (n=1931) | Png et al. 2011 | ||
| Measles | CD46 | co-receptor | 66 candidate SNPs across 3 genes3 | 744 | replication study, prev. identified | Ovsyannikova et al. 2011 |
| Influenza | HLA-DRB1, HLA-DQB1 | HLA class II molecules | HLA class II alleles4 | 73 | Gelder et al. 2002 | |
| HLA-A | HLA class I molecule | HLA class I and class II alleles5 | 184 | Poland et al. 2008 | ||
| IL6, IL12B, IFNB1 | cytokine | candidate SNPs in cytokines5 | 184 | SNPs in coding or regulatory region only | Poland et al. 2008 | |
| IL1R1,IL2RA, IL10RA,IL12RB2, IL1RN | cytokine receptor | candidate SNPs in cytokine receptors5 | 184 | SNPs in coding or regulatory region only | Poland et al. 2008 | |
| NAPSA, GLMP, GM2A, DYNL1, SNX29, TAP2,FGD2 | antigen transport and processing | GWAS and gene expression6 | 199 | reproduced in second group (n=125) | Franco et al. 2013 | |
| JUP, FBLN5 | cell junction and adhesion | genotype and gene expression6 | 199 | reproduced in second group (n=125) | Franco et al. 2013 | |
| OAS1 | antiviral response | genotype and gene expression6 | 199 | reproduced in second group (n=125) | Franco et al. 2013 | |
| LST1 | lymphocyte proliferation inhibition | genotype and gene expression6 | 199 | reproduced in second group (n=125) | Franco et al. 2013 | |
| CHST13, PAM | metabolism | genotype and gene expression6 | 199 | reproduced in second group (n=125) | Franco et al. 2013 | |
| RPL14, NAPSB, DIP2A, LRRC37A4, NSG1, HRC2 | various or unknown | genotype and gene expression6 | 199 | reproduced in second group (n=125) | Franco et al. 2013 | |
| IFNL3 | cytokine | 2 candidate SNPs in IFNL3 gene7 | 196 | reproduced in healthy volunteers (n=28) | Egli et al. 2014 |
infants
Indonesian population over 5 y of age
schoolchildren
influenza risk group according to ACIP
18–40 y old male Caucasians
18–40 y old males, ethnically homogeneous
immunosuppressed patients
Several genome wide association studies (GWAS) investigated the antibody response to various other vaccines, e.g. hepatitis B,81 smallpox,82 measles19,73,83,84 and rubella.85 These studies mainly identified SNPs in cytokines, cytokine receptors and co-receptors, but these findings are hard to interpret, because there are no replication studies by other research groups and none of the SNPs has been investigated on the RNA or protein level, which is required though for a biological interpretation.86 Similarly to influenza vaccination, variations in HLA molecules have been associated to hepatitis B vaccine response: Png et al. identified three independent variants in HLA class II and class III regions in an Indonesian population.87 Interestingly, polymorphisms associated to antibody response in cytokines and cytokine receptors have been found in genotyping studies of cytokine coding genes but not in genome-wide genotyping studies. Common caveats of GWAS are summarized in the next section.
Toward personalized vaccination strategies: identification and investigation of SNP impacts
Gene-association studies propose the first hypothesis
In summary, the impact of genetic polymorphisms on the vaccine induced humoral immune response has been studied mostly in GWAS despite two main caveats: First, the replication of GWAS results proves to be difficult, and second, GWAS show at best statistical but not causal associations. In order to infer causal associations and to understand mechanistic details, the impact of SNPs has to be studied on several levels such as on the RNA or protein level as well as on the immune cell response and on the host antibody response12 (see Fig. 2). However, it is reasonable to infer the first hypothesis from a genetic association with the host antibody response, because not every SNP that affects the RNA or protein level has a physiological effect.
Figure 2.
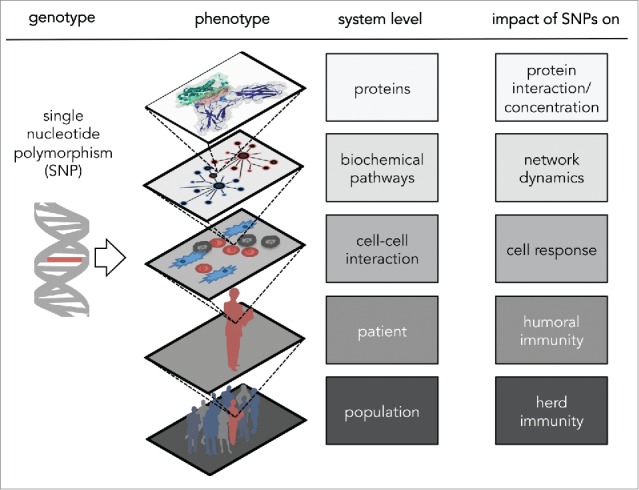
Multiscale impact of single nucleotide polymorphisms (SNPs).
Several factors complicate the replication of genetic association data: Many GWAS are insufficient for solid statistical analysis due to various reasons, e.g., an insufficient case/control ratio, an insufficient sample size or different case/control population groups. The linkage disequilibrium and thus the selected tag SNPs strongly influences the statistical power of a sample size and it is not clear to which extent tag and haplotype maps are transferable across population groups.86,88 Furthermore, the required sample size increases with the number of tested SNPs (when investigating small effects) and decreases for high minor allele frequencies and high prevalence of the investigated phenotype (e.g., weak antibody response to vaccination).86,89,90 This is the reason why GWAS that test few SNPs in risk groups need relatively low sample sizes. In contrast, a healthy group may show a very robust immune response such that a large group needs to be tested to find a particular geno-/phenotype association. In a less robust patient cohort, e.g., after transplantation, lower patient numbers may depict a particular phenotype. However, confounding effects such as differences in the level of immunosuppression must be ruled out thoroughly. A further caveat is that definitions of vaccine response phenotypes and tests are often not standardized. The surrogate marker “seroprotection” has an arbitrary titer for different pathogens, e.g. >1:40 for influenza. Conventional serological parameters can seriously over- or underestimate the clinical protection of an individual.91
However, if no prior knowledge of vaccination-related genes exists, a GWAS can be helpful to propose a first hypothesis86: A GWAS identifies a broad set of candidate genes and alleles associated to a vaccine induced antibody level, which then need to be replicated and associated to further vaccine response phenotypes, such as cytokine profiles. Several papers have highlighted that the careful design of a GWAS significantly improves its quality.86,89,90 Subsequent case-control studies are required to confirm the first findings and, finally, the exploration of SNP impacts on RNA, protein and immune cell level will complement GWAS results and help to understand the underlying mechanism behind genotype-associated vaccine response (see Fig. 2).
Although various genetic associations with vaccine response already have been proposed, only few replication studies and subsequent research studies have been performed. In contrast to vaccine research, the research of genetic factors for cancer susceptibility and cancer therapy is more advanced.92,93 A bibliometric analysis of recent publications in vaccine and/or SNP related research reflects the current scientific landscape (see Fig. 3; 94): Although immunobiological research (green cluster) shows close proximity to genetic studies (blue cluster), vaccine research and clinical studies (red cluster) are almost separated from genetic studies. In contrast, cancer research (yellow cluster) highly overlaps with both genetic studies and immunobiological research.
Figure 3.

Bibliometric map automatically created with Visualization of Similarity (VOS) viewer. Scientific terms are clustered with respect to their co-occurrence in title and abstract of PubMed references, which are published over the last 3 y (2013/01/01 - 2015/08/14) with ‘vaccination’, ‘vaccine’, ‘humoral immune response’, ‘single nucleotide polymorphism(s)’ or ‘SNP(s)' in title. Term proximity reflects co-occurrence and term size reflects occurrence frequency. Genetic terms (blue cluster) overlap with cancer terms (yellow cluster) and show close proximity to immunobiological terms (green cluster). In contrast, clinical and vaccination terms (red cluster) are almost separated from genetic terms. Bridging terms are for instance ‘liver transplantation’, ‘chronic hepatitis' and ‘ethnicity’. In total, 4,366 terms from 16,658 references are clustered.
Experimentally validated mathematical models unravel mechanistic details
Systems Biology applies mathematical models to test mechanistic hypotheses on biological processes: For instance, transport and recycling processes of cell surface recepors or signaling pathways are simulated and the simulation results are validated against experimental data.95,96 Such mechanistic models can be also applied to test hypotheses on how SNPs modulate the immunological signaling network and cause the observed variability in vaccine response: Cell signaling and effector cell communication can be formalized in mathematical models and simulated.97,98 The consequences of the investigated genetic polymorphism need to be implemented, for instance in the form of changes in protein concentration or changes in kinetic factors. If the simulation results do not match the observed vaccine response phenotype, further experiments are required to identify missing parts in the model such as crucial interactions. If simulation results match experimental results, perturbation experiments can be applied to test the predictive power of the model: for instance, antagonists can be used to block cytokine receptors. If the model is able to predict the impact of such perturbations, it can be further used to test modifications for improved antibody response. If the experimental tests do not show the same results as the model, the model has to be validated again. This iteration between experiments and mathematical modeling helps to unravel crucial interactions involved in vaccine response. Finally, such models can be used to predict the vaccine response based on patient data. Predictive models enable in silico experiments, for instance to determine potential adjuvant targets or to suggest personalized vaccination strategies (e.g., antigen amount and vaccination frequency).13,99,100
Conclusion
In summary, the association of SNPs and vaccine outcome has just begun. Important steps for a more profound understanding of these associations will be the mechanistic exploration of the impact of SNPs through an integrative analysis of gene expression, protein and immune cell data and their integration in mathematical models. In the near future we hope to use this knowledge to improve current vaccine strategies and develop new types of vaccines.
Abbreviations
- CD
Cluster of differentiation
- CMV
Cytomegalovirus
- GWAS
Genome-wide association study
- HAI
Hemagglutination inhibition assay
- HBV
Hepatitis B virus
- HCV
Hepatitis C virus
- HIV
Human immunodeficiency virus
- HLA
Human leukocyte antigen
- IFN
Interferon
- IL
Interleukin
- JAK
Janus kinase
- MAF
Minor allele frequency
- MyD88
Myeloid differentiation primary response protein 88
- NF-κB
Nuclear factor-κB
- PBMC
Peripheral blood mononuclear cell
- PCR
Polymerase chain reaction
- PEG-IFN
Pegylated interferon
- PRR
Pattern recognition receptor
- RBV
Ribavirin;
- RNA
Ribonucleic acid
- SNP
Single nucleotide polymorphism
- spp
Species pluralis
- STAT
Signal transducer and activator of transcription
- Th1
T helper cells type I
- Th2
T helper cells type II
- TICAM1
TIR domain containing adapter molecule 1
- TIR
Toll/interleukin-1 receptor
- TLR
Toll-like receptor
- UTR
Untranslated region
Disclosure of potential conflicts of interest
The authors have no conflict of interest and nothing to disclose.
Acknowledgments
We thank Jörg Stelling (ETH Zurich), Marco Kokic (University of Heidelberg), Julia Hartmann (University of Basel) and Mohammedyaseen Syedbasha (University of Basel) for critical reading of the manuscript.
Funding
A.E. was supported by a research grants from the “SNSF Ambizione Score” program (PZ00P3_154709), “Forschungsfond” University of Basel, Bangerter-Rhyner Stiftung, Stiftungsinfektionskrankheiten Basel, and “SystemsX” program (9th call). J.L. acknowledges support by an iPhD fellowship of the SystemsX.ch initiative in systems biology program (9th call).
References
- [1].Andre FE, Booy R, Bock HL, Clemens J, Datta SK, John TJ, Lee BW, Lolekha S, Peltola H, Ruff TA, et al.. Vaccination greatly reduces disease, disability, death and inequity worldwide. Bull World Health Organ 2008; 86:140-6; PMID:18297169; http://dx.doi.org/ 10.2471/BLT.07.040089 [DOI] [PMC free article] [PubMed] [Google Scholar]
- [2].Kaslow DC, Biernaux S. RTS,S: Toward a first landmark on the Malaria Vaccine Technology Roadmap. Vaccine 2015; 33(52):7425-32 [DOI] [PubMed] [Google Scholar]
- [3].Mangtani P, Abubakar I, Ariti C, Beynon R, Pimpin L, Fine PE, Rodrigues LC, Smith PG, Lipman M, Whiting PF, et al.. Protection by BCG vaccine against tuberculosis: a systematic review of randomized controlled trials. Clin Infect Dis 2014; 58:470-80; PMID:24336911; http://dx.doi.org/ 10.1093/cid/cit790 [DOI] [PubMed] [Google Scholar]
- [4].Siegrist CA, Aspinall R. B-cell responses to vaccination at the extremes of age. Nat Rev Immunol 2009; 9:185-94; PMID:19240757; http://dx.doi.org/ 10.1038/nri2508 [DOI] [PubMed] [Google Scholar]
- [5].Faucette AN, Unger BL, Gonik B, Chen K. Maternal vaccination: moving the science forward. Hum Reprod Update 2015; 21:119-35; PMID:25015234; http://dx.doi.org/ 10.1093/humupd/dmu041 [DOI] [PMC free article] [PubMed] [Google Scholar]
- [6].Smith SA, Poland GA, American Diabetes A. Influenza and pneumococcal immunization in diabetes. Diabetes Care 2004; 27 Suppl 1:S111-113; PMID:14693944 [DOI] [PubMed] [Google Scholar]
- [7].Gelinck LB, Teng YK, Rimmelzwaan GF, van den Bemt BJ, Kroon FP, van Laar JM. Poor serological responses upon influenza vaccination in patients with rheumatoid arthritis treated with rituximab. Ann Rheum Dis 2007; 66:1402-3; PMID:17881666; http://dx.doi.org/ 10.1136/ard.2007.071878 [DOI] [PMC free article] [PubMed] [Google Scholar]
- [8].van Assen S, Holvast A, Benne CA, Posthumus MD, van Leeuwen MA, Voskuyl AE, Blom M, Risselada AP, de Haan A, Westra J, et al.. Humoral responses after influenza vaccination are severely reduced in patients with rheumatoid arthritis treated with rituximab. Arthritis Rheum 2010; 62:75-81; PMID:20039396; http://dx.doi.org/ 10.1002/art.25033 [DOI] [PubMed] [Google Scholar]
- [9].Manuel O, Pascual M, Hoschler K, Giulieri S, Alves D, Ellefsen K, Bart PA, Venetz JP, Calandra T, Cavassini M. Humoral response to the influenza A H1N1/09 monovalent AS03-adjuvanted vaccine in immunocompromised patients. Clin Infect Dis 2011; 52:248-56; PMID:21288852; http://dx.doi.org/ 10.1093/cid/ciq104 [DOI] [PubMed] [Google Scholar]
- [10].Kunisaki KM, Janoff EN. Influenza in immunosuppressed populations: a review of infection frequency, morbidity, mortality, and vaccine responses. Lancet Infect Dis 2009; 9:493-504; PMID:19628174; http://dx.doi.org/ 10.1016/S1473-3099(09)70175-6 [DOI] [PMC free article] [PubMed] [Google Scholar]
- [11].Ljungman P. Vaccination of immunocompromised patients. Clin Microbiol Infect 2012; 18 Suppl 5:93-9; PMID:23051059; http://dx.doi.org/ 10.1111/j.1469-0691.2012.03971.x [DOI] [PubMed] [Google Scholar]
- [12].Hagan T, Nakaya HI, Subramaniam S, Pulendran B. Systems vaccinology: Enabling rational vaccine design with systems biological approaches. Vaccine 2015; 33(40):5294-301; PMID:25858860 [DOI] [PMC free article] [PubMed] [Google Scholar]
- [13].Germain RN. Vaccines and the future of human immunology. Immunity 2010; 33:441-50; PMID:21029956; http://dx.doi.org/ 10.1016/j.immuni.2010.09.014 [DOI] [PubMed] [Google Scholar]
- [14].Fish EN. The X-files in immunity: sex-based differences predispose immune responses. Nat Rev Immunol 2008; 8:737-44; PMID:18728636; http://dx.doi.org/ 10.1038/nri2394 [DOI] [PMC free article] [PubMed] [Google Scholar]
- [15].O'Shea D, Widmer LA, Stelling J, Egli A. Changing face of vaccination in immunocompromised hosts. Curr Infect Dis Rep 2014; 16:420; PMID:24992978; http://dx.doi.org/ 10.1007/s11908-014-0420-2 [DOI] [PubMed] [Google Scholar]
- [16].Egli A, Santer D, Barakat K, Zand M, Levin A, Vollmer M, Weisser M, Khanna N, Kumar D, Tyrrell L, et al.. Vaccine adjuvants–understanding molecular mechanisms to improve vaccines. Swiss Med Wkly 2014; 144:w13940; PMID:24844935 [DOI] [PubMed] [Google Scholar]
- [17].Franco LM, Bucasas KL, Wells JM, Nino D, Wang X, Zapata GE, Arden N, Renwick A, Yu P, Quarles JM, et al.. Integrative genomic analysis of the human immune response to influenza vaccination. Elife 2013; 2:e00299; PMID:23878721; http://dx.doi.org/ 10.7554/eLife.00299 [DOI] [PMC free article] [PubMed] [Google Scholar]
- [18].Tan PL, Jacobson RM, Poland GA, Jacobsen SJ, Pankratz VS. Twin studies of immunogenicity–determining the genetic contribution to vaccine failure. Vaccine 2001; 19:2434-9; PMID:11257374; http://dx.doi.org/ 10.1016/S0264-410X(00)00468-0 [DOI] [PubMed] [Google Scholar]
- [19].Ovsyannikova IG, Haralambieva IH, Vierkant RA, O'Byrne MM, Jacobson RM, Poland GA. The association of CD46, SLAM and CD209 cellular receptor gene SNPs with variations in measles vaccine-induced immune responses: a replication study and examination of novel polymorphisms. Hum Hered 2011; 72:206-23; PMID:22086389; http://dx.doi.org/ 10.1159/000331585 [DOI] [PMC free article] [PubMed] [Google Scholar]
- [20].Ferreira PG, Dermitzakis ET. Immune response is a personal matter. Elife 2013; 2:e00899; PMID:23878727; http://dx.doi.org/ 10.7554/eLife.00899 [DOI] [PMC free article] [PubMed] [Google Scholar]
- [21].Pulendran B. Systems vaccinology: probing humanity's diverse immune systems with vaccines. Proc Natl Acad Sci U S A 2014; 111:12300-6; PMID:25136102; http://dx.doi.org/ 10.1073/pnas.1400476111 [DOI] [PMC free article] [PubMed] [Google Scholar]
- [22].Nauta JJ, Beyer WE, Osterhaus AD. On the relationship between mean antibody level, seroprotection and clinical protection from influenza. Biologicals 2009; 37:216-21; PMID:19268607; http://dx.doi.org/ 10.1016/j.biologicals.2009.02.002 [DOI] [PubMed] [Google Scholar]
- [23].Kawai T, Akira S. The role of pattern-recognition receptors in innate immunity: update on Toll-like receptors. Nat Immunol 2010; 11:373-84; PMID:20404851; http://dx.doi.org/ 10.1038/ni.1863 [DOI] [PubMed] [Google Scholar]
- [24].Honda K, Taniguchi T. IRFs: master regulators of signalling by Toll-like receptors and cytosolic pattern-recognition receptors. Nat Rev Immunol 2006; 6:644-58; PMID:16932750; http://dx.doi.org/ 10.1038/nri1900 [DOI] [PubMed] [Google Scholar]
- [25].Chen L, Flies DB. Molecular mechanisms of T cell co-stimulation and co-inhibition. Nat Rev Immunol 2013; 13:227-42; PMID:23470321; http://dx.doi.org/ 10.1038/nri3405 [DOI] [PMC free article] [PubMed] [Google Scholar]
- [26].Kaplan MH, Schindler U, Smiley ST, Grusby MJ. Stat6 is required for mediating responses to IL-4 and for development of Th2 cells. Immunity 1996; 4:313-9; PMID:8624821; http://dx.doi.org/ 10.1016/S1074-7613(00)80439-2 [DOI] [PubMed] [Google Scholar]
- [27].Mari N, Hercor M, Denanglaire S, Leo O, Andris F. The capacity of Th2 lymphocytes to deliver B-cell help requires expression of the transcription factor STAT3. Eur J Immunol 2013; 43(6):1489-98; PMID:23504518 [DOI] [PubMed] [Google Scholar]
- [28].Randolph DA, Huang G, Carruthers CJ, Bromley LE, Chaplin DD. The role of CCR7 in TH1 and TH2 cell localization and delivery of B cell help in vivo. Science 1999; 286:2159-62; PMID:10591648; http://dx.doi.org/ 10.1126/science.286.5447.2159 [DOI] [PubMed] [Google Scholar]
- [29].MacLeod H, Wetzler LM. T cell activation by TLRs: a role for TLRs in the adaptive immune response. Sci STKE 2007; 2007:pe48; PMID:17785715; http://dx.doi.org/ 10.1126/stke.4022007pe48 [DOI] [PubMed] [Google Scholar]
- [30].Tartey S, Matsushita K, Imamura T, Wakabayashi A, Ori D, Mino T, Takeuchi O. Essential Function for the Nuclear Protein Akirin2 in B Cell Activation and Humoral Immune Responses. J Immunol 2015; 195:519-27; PMID:26041538; http://dx.doi.org/ 10.4049/jimmunol.1500373 [DOI] [PubMed] [Google Scholar]
- [31].Albert R. Scale-free networks in cell biology. J Cell Sci 2005; 118:4947-57; PMID:16254242; http://dx.doi.org/ 10.1242/jcs.02714 [DOI] [PubMed] [Google Scholar]
- [32].Han HW, Ohn JH, Moon J, Kim JH. Yin and Yang of disease genes and death genes between reciprocally scale-free biological networks. Nucleic Acids Res 2013; 41:9209-17; PMID:23935122; http://dx.doi.org/ 10.1093/nar/gkt683 [DOI] [PMC free article] [PubMed] [Google Scholar]
- [33].Wagner A. Robustness against mutations in genetic networks of yeast. Nat Genet 2000; 24:355-61; PMID:10742097; http://dx.doi.org/ 10.1038/74174 [DOI] [PubMed] [Google Scholar]
- [34].Albert R, Jeong H, Barabasi AL. Error and attack tolerance of complex networks. Nature 2000; 406:378-82; PMID:10935628; http://dx.doi.org/ 10.1038/35019019 [DOI] [PubMed] [Google Scholar]
- [35].Stelling J, Sauer U, Szallasi Z, Doyle FJ 3rd, Doyle J. Robustness of cellular functions. Cell 2004; 118:675-85; PMID:15369668; http://dx.doi.org/ 10.1016/j.cell.2004.09.008 [DOI] [PubMed] [Google Scholar]
- [36].Kitano H. Biological robustness. Nat Rev Genet 2004; 5:826-37; PMID:15520792; http://dx.doi.org/ 10.1038/nrg1471 [DOI] [PubMed] [Google Scholar]
- [37].Lavoie TB, Kalie E, Crisafulli-Cabatu S, Abramovich R, DiGioia G, Moolchan K, Pestka S, Schreiber G. Binding and activity of all human α interferon subtypes. Cytokine 2011; 56:282-9; PMID:21856167; http://dx.doi.org/ 10.1016/j.cyto.2011.07.019 [DOI] [PubMed] [Google Scholar]
- [38].Abdul-Sater AA, Majoros A, Plumlee CR, Perry S, Gu AD, Lee C, Shresta S, Decker T, Schindler C. Different STAT Transcription Complexes Drive Early and Delayed Responses to Type I IFNs. J Immunol 2015; 195:210-6; PMID:26019270; http://dx.doi.org/ 10.4049/jimmunol.1401139 [DOI] [PMC free article] [PubMed] [Google Scholar]
- [39].Kohler UA, Bohm F, Rolfs F, Egger M, Hornemann T, Pasparakis M, Weber A, Werner S. NF-kappaB/RelA and Nrf2 cooperate to maintain hepatocyte integrity and to prevent development of hepatocellular adenoma. J Hepatol 2016; 64(1):94-102; PMID:26348541. [DOI] [PubMed] [Google Scholar]
- [40].Bank S, Skytt Andersen P, Burisch J, Pedersen N, Roug S, Galsgaard J, Ydegaard Turino S, Broder Brodersen J, Rashid S, Kaiser Rasmussen B, et al.. Polymorphisms in the inflammatory pathway genes TLR2, TLR4, TLR9, LY96, NFKBIA, NFKB1, TNFA, TNFRSF1A, IL6R, IL10, IL23R, PTPN22, and PPARG are associated with susceptibility of inflammatory bowel disease in a Danish cohort. PLoS One 2014; 9:e98815; PMID:24971461; http://dx.doi.org/ 10.1371/journal.pone.0098815 [DOI] [PMC free article] [PubMed] [Google Scholar]
- [41].Gough SC, Simmonds MJ. The HLA Region and Autoimmune Disease: Associations and Mechanisms of Action. Curr Genomics 2007; 8:453-65; PMID:19412418; http://dx.doi.org/ 10.2174/138920207783591690 [DOI] [PMC free article] [PubMed] [Google Scholar]
- [42].Smahi A, Courtois G, Rabia SH, Doffinger R, Bodemer C, Munnich A, Casanova JL, Israël A. The NF-kappaB signalling pathway in human diseases: from incontinentia pigmenti to ectodermal dysplasias and immune-deficiency syndromes. Hum Mol Genet 2002; 11:2371-5; PMID:12351572; http://dx.doi.org/ 10.1093/hmg/11.20.2371 [DOI] [PubMed] [Google Scholar]
- [43].Halloran PF. Immunosuppressive drugs for kidney transplantation. N Engl J Med 2004; 351:2715-29; PMID:15616206; http://dx.doi.org/ 10.1056/NEJMra033540 [DOI] [PubMed] [Google Scholar]
- [44].Scheinfeld N. A comprehensive review and evaluation of the side effects of the tumor necrosis factor α blockers etanercept, infliximab and adalimumab. J Dermatolog Treat 2004; 15:280-94; PMID:15370396; http://dx.doi.org/ 10.1080/09546630410017275 [DOI] [PubMed] [Google Scholar]
- [45].Rappuoli R, Aderem A. A 2020 vision for vaccines against HIV, tuberculosis and malaria. Nature 2011; 473:463-9; PMID:21614073; http://dx.doi.org/ 10.1038/nature10124 [DOI] [PubMed] [Google Scholar]
- [46].Prachi P, Donati C, Masciopinto F, Rappuoli R, Bagnoli F. Deep sequencing in pre- and clinical vaccine research. Public Health Genomics 2013; 16:62-8; PMID:23548719; http://dx.doi.org/ 10.1159/000345611 [DOI] [PubMed] [Google Scholar]
- [47].Buhler S, Sanchez-Mazas A. HLA DNA sequence variation among human populations: molecular signatures of demographic and selective events. PLoS One 2011; 6:e14643; PMID:21408106; http://dx.doi.org/ 10.1371/journal.pone.0014643 [DOI] [PMC free article] [PubMed] [Google Scholar]
- [48].Brookes AJ. The essence of SNPs. Gene 1999; 234:177-86; PMID:10395891; http://dx.doi.org/ 10.1016/S0378-1119(99)00219-X [DOI] [PubMed] [Google Scholar]
- [49].Snyder MW, Adey A, Kitzman JO, Shendure J. Haplotype-resolved genome sequencing: experimental methods and applications. Nat Rev Genet 2015; 16:344-58; PMID:25948246; http://dx.doi.org/ 10.1038/nrg3903 [DOI] [PubMed] [Google Scholar]
- [50].Daly MJ, Rioux JD, Schaffner SF, Hudson TJ, Lander ES. High-resolution haplotype structure in the human genome. Nat Genet 2001; 29:229-32; PMID:11586305; http://dx.doi.org/ 10.1038/ng1001-229 [DOI] [PubMed] [Google Scholar]
- [51].Kim S, Misra A. SNP genotyping: technologies and biomedical applications. Annu Rev Biomed Eng 2007; 9:289-320; PMID:17391067; http://dx.doi.org/ 10.1146/annurev.bioeng.9.060906.152037 [DOI] [PubMed] [Google Scholar]
- [52].Lander ES, Linton LM, Birren B, Nusbaum C, Zody MC, Baldwin J, Devon K, Dewar K, Doyle M, FitzHugh W, et al.. Initial sequencing and analysis of the human genome. Nature 2001; 409:860-921; PMID:11237011; http://dx.doi.org/ 10.1038/35057062 [DOI] [PubMed] [Google Scholar]
- [53].Sachidanandam R, Weissman D, Schmidt SC, Kakol JM, Stein LD, Marth G, Sherry S, Mullikin JC, Mortimore BJ, Willey DL, et al.. A map of human genome sequence variation containing 1.42 million single nucleotide polymorphisms. Nature 2001; 409:928-33; PMID:11237013; http://dx.doi.org/ 10.1038/35057149 [DOI] [PubMed] [Google Scholar]
- [54].Wang DG, Fan JB, Siao CJ, Berno A, Young P, Sapolsky R, Ghandour G, Perkins N, Winchester E, Spencer J, et al.. Large-scale identification, mapping, and genotyping of single-nucleotide polymorphisms in the human genome. Science 1998; 280:1077-82; PMID:9582121; http://dx.doi.org/ 10.1126/science.280.5366.1077 [DOI] [PubMed] [Google Scholar]
- [55].Musumeci L, Arthur JW, Cheung FS, Hoque A, Lippman S, Reichardt JK. Single nucleotide differences (SNDs) in the dbSNP database may lead to errors in genotyping and haplotyping studies. Hum Mutat 2010; 31:67-73; PMID:19877174; http://dx.doi.org/ 10.1002/humu.21137 [DOI] [PMC free article] [PubMed] [Google Scholar]
- [56].Mitchell AA, Zwick ME, Chakravarti A, Cutler DJ. Discrepancies in dbSNP confirmation rates and allele frequency distributions from varying genotyping error rates and patterns. Bioinformatics 2004; 20:1022-32; PMID:14764571; http://dx.doi.org/ 10.1093/bioinformatics/bth034 [DOI] [PubMed] [Google Scholar]
- [57].International HapMap C, Altshuler DM, Gibbs RA, Peltonen L, Altshuler DM, Gibbs RA, Peltonen L, Dermitzakis E, Schaffner SF, Yu F, et al.. Integrating common and rare genetic variation in diverse human populations. Nature 2010; 467:52-8; PMID:20811451; http://dx.doi.org/ 10.1038/nature09298 [DOI] [PMC free article] [PubMed] [Google Scholar]
- [58].Shastry BS. SNPs: impact on gene function and phenotype. Methods Mol Biol 2009; 578:3-22; PMID:19768584; http://dx.doi.org/ 10.1007/978-1-60327-411-1_1 [DOI] [PubMed] [Google Scholar]
- [59].Zhang YH, Zhao Y, Li N, Peng YC, Giannoulatou E, Jin RH, Yan HP, Wu H, Liu JH, Liu N, et al.. Interferon-induced transmembrane protein-3 genetic variant rs12252-C is associated with severe influenza in Chinese individuals. Nat Commun 2013; 4:1418; PMID:23361009; http://dx.doi.org/ 10.1038/ncomms2433 [DOI] [PMC free article] [PubMed] [Google Scholar]
- [60].Dong ZX, Zhou HJ, Xiang XG, Guo SM, Zhuang Y, Zhao GD, Xie Q. IL28B genetic variations are associated with treatment response of patients with chronic hepatitis C in a Chinese Han population. J Dig Dis 2015; 16:90-7; PMID:25312023; http://dx.doi.org/ 10.1111/1751-2980.12202 [DOI] [PubMed] [Google Scholar]
- [61].Fernandez-Carrillo C, Coto-Llerena M, Gonzalez P, Crespo G, Mensa L, Caro-Pérez N, Gambato M, Navasa M, Forns X, Pérez-del-Pulgar S. IFNL4 polymorphism predicts response to hepatitis C treatment after liver transplantation. J Clin Virol 2014; 61:282-5; PMID:25130512; http://dx.doi.org/ 10.1016/j.jcv.2014.07.015 [DOI] [PubMed] [Google Scholar]
- [62].Egli A, Santer DM, O'Shea D, Barakat K, Syedbasha M, Vollmer M, Baluch A, Bhat R, Groenendyk J, Joyce MA, et al.. IL-28B is a key regulator of B- and T-cell vaccine responses against influenza. PLoS Pathog 2014; 10:e1004556; PMID:25503988; http://dx.doi.org/ 10.1371/journal.ppat.1004556 [DOI] [PMC free article] [PubMed] [Google Scholar]
- [63].Lazear HM, Nice TJ, Diamond MS. Interferon-lambda: Immune Functions at Barrier Surfaces and Beyond. Immunity 2015; 43:15-28; PMID:26200010; http://dx.doi.org/ 10.1016/j.immuni.2015.07.001 [DOI] [PMC free article] [PubMed] [Google Scholar]
- [64].Egli A, Santer MD, O'Shea D, Tyrrell DL, Houghton M. The impact of the interferon-lambda family on the innate and adaptive immune response to viral infections. Emerg Infect Dis 2014;3(7):e51. http://dx.doi.org/ 10.1038/emi.2014.51 [DOI] [PMC free article] [PubMed] [Google Scholar]
- [65].Chinnaswamy S, Chatterjee S, Boopathi R, Mukherjee S, Bhattacharjee S, Kundu TK. A single nucleotide polymorphism associated with hepatitis C virus infections located in the distal region of the IL28B promoter influences NF-kappaB-mediated gene transcription. PLoS One 2013; 8:e75495; PMID:24116050; http://dx.doi.org/ 10.1371/journal.pone.0075495 [DOI] [PMC free article] [PubMed] [Google Scholar]
- [66].Fischer J, Bohm S, Scholz M, Muller T, Witt H, George J, Sarrazin C, Susser S, Schott E, Suppiah V, et al.. Combined effects of different interleukin-28B gene variants on the outcome of dual combination therapy in chronic hepatitis C virus type 1 infection. Hepatology 2012; 55:1700-10; PMID:22234924; http://dx.doi.org/ 10.1002/hep.25582 [DOI] [PubMed] [Google Scholar]
- [67].Aparicio E, Parera M, Franco S, Perez-Alvarez N, Tural C, Clotet B, Martínez MA. IL28B SNP rs8099917 is strongly associated with pegylated interferon-α and ribavirin therapy treatment failure in HCV/HIV-1 coinfected patients. PLoS One 2010; 5:e13771; PMID:21048934; http://dx.doi.org/ 10.1371/journal.pone.0013771 [DOI] [PMC free article] [PubMed] [Google Scholar]
- [68].Resham S, Manzoor S, Imran M, Saalim M, Naseem S, Azam S. Interleukin- 28B: a prognostic marker in interferon based therapy of chronic HCV patients of the Pakistan with variable treatment response. APMIS 2015; 123(9):765-71 [DOI] [PubMed] [Google Scholar]
- [69].Suppiah V, Moldovan M, Ahlenstiel G, Berg T, Weltman M, Abate ML, Bassendine M, Spengler U, Dore GJ, Powell E, et al.. IL28B is associated with response to chronic hepatitis C interferon-α and ribavirin therapy. Nat Genet 2009; 41:1100-4; PMID:19749758; http://dx.doi.org/ 10.1038/ng.447 [DOI] [PubMed] [Google Scholar]
- [70].Tanaka Y, Nishida N, Sugiyama M, Kurosaki M, Matsuura K, Sakamoto N, Nakagawa M, Korenaga M, Hino K, Hige S, et al.. Genome-wide association of IL28B with response to pegylated interferon-α and ribavirin therapy for chronic hepatitis C. Nat Genet 2009; 41:1105-9; PMID:19749757; http://dx.doi.org/ 10.1038/ng.449 [DOI] [PubMed] [Google Scholar]
- [71].Bibert S, Wojtowicz A, Taffe P, Manuel O, Bernasconi E, Furrer H, Günthard HF, Hoffmann M, Kaiser L, Osthoff M, et al.. The IFNL3/4 DeltaG variant increases susceptibility to cytomegalovirus retinitis among HIV-infected patients. AIDS 2014; 28:1885-9; PMID:25259701; http://dx.doi.org/ 10.1097/QAD.0000000000000379 [DOI] [PubMed] [Google Scholar]
- [72].Manuel O, Wojtowicz A, Bibert S, Mueller NJ, van Delden C, Hirsch HH, Steiger J, Stern M, Egli A, Garzoni C, et al.. Influence of IFNL3/4 polymorphisms on the incidence of cytomegalovirus infection after solid-organ transplantation. J Infect Dis 2015; 211:906-14; PMID:25301956; http://dx.doi.org/ 10.1093/infdis/jiu557 [DOI] [PubMed] [Google Scholar]
- [73].Haralambieva IH, Ovsyannikova IG, Umlauf BJ, Vierkant RA, Shane Pankratz V, Jacobson RM, Poland GA. Genetic polymorphisms in host antiviral genes: associations with humoral and cellular immunity to measles vaccine. Vaccine 2011; 29:8988-97; PMID:21939710; http://dx.doi.org/ 10.1016/j.vaccine.2011.09.043 [DOI] [PMC free article] [PubMed] [Google Scholar]
- [74].Srinivas S, Dai J, Eskdale J, Gallagher GE, Megjugorac NJ, Gallagher G. Interferon-lambda1 (interleukin-29) preferentially down-regulates interleukin-13 over other T helper type 2 cytokine responses in vitro. Immunology 2008; 125:492-502; PMID:18547367; http://dx.doi.org/ 10.1111/j.1365-2567.2008.02862.x [DOI] [PMC free article] [PubMed] [Google Scholar]
- [75].Jordan WJ, Eskdale J, Srinivas S, Pekarek V, Kelner D, Rodia M, Gallagher G. Human interferon lambda-1 (IFN-lambda1/IL-29) modulates the Th1/Th2 response. Genes Immun 2007; 8:254-61; PMID:17361203; http://dx.doi.org/ 10.1038/sj.gene.6364382 [DOI] [PubMed] [Google Scholar]
- [76].Dai J, Megjugorac NJ, Gallagher GE, Yu RY, Gallagher G. IFN-lambda1 (IL-29) inhibits GATA3 expression and suppresses Th2 responses in human naive and memory T cells. Blood 2009; 113:5829-38; PMID:19346497; http://dx.doi.org/ 10.1182/blood-2008-09-179507 [DOI] [PubMed] [Google Scholar]
- [77].Koltsida O, Hausding M, Stavropoulos A, Koch S, Tzelepis G, Ubel C, Kotenko SV, Sideras P, Lehr HA, Tepe M, et al.. IL-28A (IFN-lambda2) modulates lung DC function to promote Th1 immune skewing and suppress allergic airway disease. EMBO Mol Med 2011; 3:348-61; PMID:21538995; http://dx.doi.org/ 10.1002/emmm.201100142 [DOI] [PMC free article] [PubMed] [Google Scholar]
- [78].Steck T, Albert P, Weppler M, Becker W, Spingler H, Würfel W, von Hertwig I. [The effect of an intrauterine device on passive sperm transport in the uterus and fallopian tube at the time of ovulation]. Gynakol Rundsch 1989; 29 Suppl 2:430-2; PMID:2613070; http://dx.doi.org/ 10.1159/000271144 [DOI] [PubMed] [Google Scholar]
- [79].Gelder CM, Lambkin R, Hart KW, Fleming D, Williams OM, Bunce M, Welsh KI, Marshall SE, Oxford J. Associations between human leukocyte antigens and nonresponsiveness to influenza vaccine. J Infect Dis 2002; 185:114-7; PMID:11756990; http://dx.doi.org/ 10.1086/338014 [DOI] [PubMed] [Google Scholar]
- [80].Poland GA, Ovsyannikova IG, Jacobson RM. Immunogenetics of seasonal influenza vaccine response. Vaccine 2008; 26 Suppl 4:D35-40; PMID:19230157; http://dx.doi.org/ 10.1016/j.vaccine.2008.07.065 [DOI] [PMC free article] [PubMed] [Google Scholar]
- [81].Hennig BJ, Fielding K, Broxholme J, Diatta M, Mendy M, Moore C, Pollard AJ, Rayco-Solon P, Sirugo G, van der Sande MA, et al.. Host genetic factors and vaccine-induced immunity to hepatitis B virus infection. PLoS One 2008; 3:e1898; PMID:18365030; http://dx.doi.org/ 10.1371/journal.pone.0001898 [DOI] [PMC free article] [PubMed] [Google Scholar]
- [82].Ovsyannikova IG, Kennedy RB, O'Byrne M, Jacobson RM, Pankratz VS, Poland GA. Genome-wide association study of antibody response to smallpox vaccine. Vaccine 2012; 30:4182-9; PMID:22542470; http://dx.doi.org/ 10.1016/j.vaccine.2012.04.055 [DOI] [PMC free article] [PubMed] [Google Scholar]
- [83].Haralambieva IH, Ovsyannikova IG, Kennedy RB, Vierkant RA, Pankratz VS, Jacobson RM, Poland GA. Associations between single nucleotide polymorphisms and haplotypes in cytokine and cytokine receptor genes and immunity to measles vaccination. Vaccine 2011; 29:7883-95; PMID:21875636; http://dx.doi.org/ 10.1016/j.vaccine.2011.08.083 [DOI] [PMC free article] [PubMed] [Google Scholar]
- [84].Kennedy RB, Ovsyannikova IG, Haralambieva IH, Lambert ND, Pankratz VS, Poland GA. Genome-wide SNP associations with rubella-specific cytokine responses in measles-mumps-rubella vaccine recipients. Immunogenetics 2014; 66:493-9; PMID:24811271; http://dx.doi.org/ 10.1007/s00251-014-0776-3 [DOI] [PMC free article] [PubMed] [Google Scholar]
- [85].Haralambieva IH, Lambert ND, Ovsyannikova IG, Kennedy RB, Larrabee BR, Pankratz VS, Poland GA. Associations between single nucleotide polymorphisms in cellular viral receptors and attachment factor-related genes and humoral immunity to rubella vaccination. PLoS One 2014; 9:e99997; PMID:24945853; http://dx.doi.org/ 10.1371/journal.pone.0099997 [DOI] [PMC free article] [PubMed] [Google Scholar]
- [86].Hattersley AT, McCarthy MI. What makes a good genetic association study? Lancet 2005; 366:1315-23; PMID:16214603; http://dx.doi.org/ 10.1016/S0140-6736(05)67531-9 [DOI] [PubMed] [Google Scholar]
- [87].Png E, Thalamuthu A, Ong RT, Snippe H, Boland GJ, Seielstad M. A genome-wide association study of hepatitis B vaccine response in an Indonesian population reveals multiple independent risk variants in the HLA region. Hum Mol Genet 2011; 20:3893-8; PMID:21764829; http://dx.doi.org/ 10.1093/hmg/ddr302 [DOI] [PubMed] [Google Scholar]
- [88].de Bakker PI, Yelensky R, Pe'er I, Gabriel SB, Daly MJ, Altshuler D. Efficiency and power in genetic association studies. Nat Genet 2005; 37:1217-23; PMID:16244653; http://dx.doi.org/ 10.1038/ng1669 [DOI] [PubMed] [Google Scholar]
- [89].Hong EP, Park JW. Sample size and statistical power calculation in genetic association studies. Genomics Inform 2012; 10:117-22; PMID:23105939; http://dx.doi.org/ 10.5808/GI.2012.10.2.117 [DOI] [PMC free article] [PubMed] [Google Scholar]
- [90].Spencer CC, Su Z, Donnelly P, Marchini J. Designing genome-wide association studies: sample size, power, imputation, and the choice of genotyping chip. PLoS Genet 2009; 5:e1000477; PMID:19492015; http://dx.doi.org/ 10.1371/journal.pgen.1000477 [DOI] [PMC free article] [PubMed] [Google Scholar]
- [91].Beyer WE, Palache AM, Luchters G, Nauta J, Osterhaus AD. Seroprotection rate, mean fold increase, seroconversion rate: which parameter adequately expresses seroresponse to influenza vaccination? Virus Res 2004; 103:125-32; PMID:15163500 [DOI] [PubMed] [Google Scholar]
- [92].King MC, Marks JH, Mandell JB, New York Breast Cancer Study G . Breast and ovarian cancer risks due to inherited mutations in BRCA1 and BRCA2. Science 2003; 302:643-6; PMID:14576434; http://dx.doi.org/ 10.1126/science.1088759 [DOI] [PubMed] [Google Scholar]
- [93].Weigelt B, Peterse JL, van 't Veer LJ. Breast cancer metastasis: markers and models. Nat Rev Cancer 2005; 5:591-602; PMID:16056258; http://dx.doi.org/ 10.1038/nrc1670 [DOI] [PubMed] [Google Scholar]
- [94].Van Eck NJ, Waltman L. Software survey: VOSviewer, a computer program for bibliometric mapping. Scientometrics 2010; 84:523-38; PMID:20585380; http://dx.doi.org/ 10.1007/s11192-009-0146-3 [DOI] [PMC free article] [PubMed] [Google Scholar]
- [95].Becker V, Schilling M, Bachmann J, Baumann U, Raue A, Maiwald T, Timmer J, Klingmüller U. Covering a broad dynamic range: information processing at the erythropoietin receptor. Science 2010; 328:1404-8; PMID:20488988; http://dx.doi.org/ 10.1126/science.1184913 [DOI] [PubMed] [Google Scholar]
- [96].Maiwald T, Schneider A, Busch H, Sahle S, Gretz N, Weiss TS, Kummer U, Klingmüller U. Combining theoretical analysis and experimental data generation reveals IRF9 as a crucial factor for accelerating interferon α-induced early antiviral signalling. FEBS J 2010; 277:4741-54; PMID:20964804; http://dx.doi.org/ 10.1111/j.1742-4658.2010.07880.x [DOI] [PubMed] [Google Scholar]
- [97].Kholodenko BN. Cell-signalling dynamics in time and space. Nat Rev Mol Cell Biol 2006; 7:165-76; PMID:16482094; http://dx.doi.org/ 10.1038/nrm1838 [DOI] [PMC free article] [PubMed] [Google Scholar]
- [98].Rangamani P, Iyengar R. Modelling cellular signalling systems. Essays Biochem 2008; 45:83-94; PMID:18793125; http://dx.doi.org/ 10.1042/bse0450083 [DOI] [PMC free article] [PubMed] [Google Scholar]
- [99].Kitano H. Computational systems biology. Nature 2002; 420:206-10; PMID:12432404; http://dx.doi.org/ 10.1038/nature01254 [DOI] [PubMed] [Google Scholar]
- [100].Butcher EC, Berg EL, Kunkel EJ. Systems biology in drug discovery. Nat Biotechnol 2004; 22:1253-9; PMID:15470465; http://dx.doi.org/ 10.1038/nbt1017 [DOI] [PubMed] [Google Scholar]