
Figure 2.
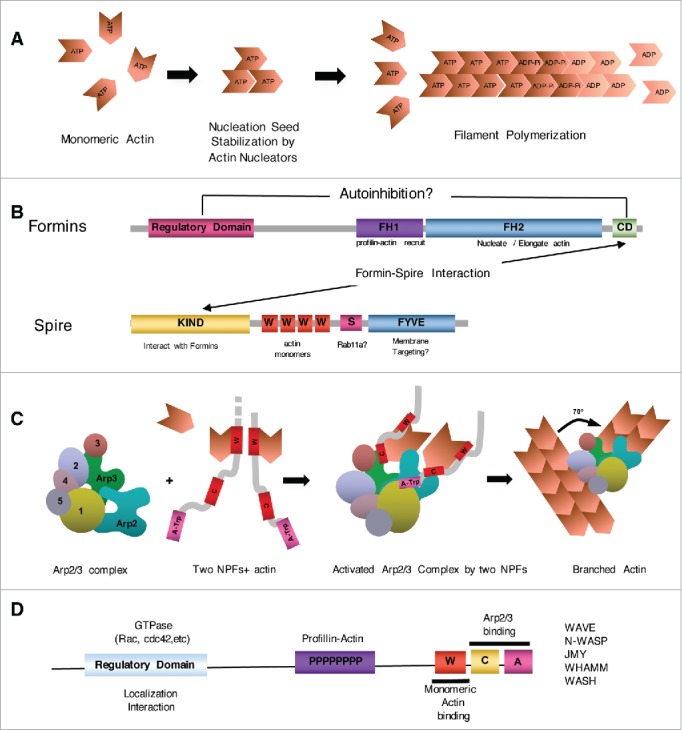
Actin nucleators and actin filament nucleation. (A) Actin polymerization is a kinetically unfavorable process and is accelerated by actin nucleators, which are families of proteins that stabilize unstable actin filament nucleation seeds. Whereas ATP-bound actin is mainly incorporated into fast-growing (also called barbed) ends of filaments, nucleotide hydrolysis occurs in the filaments, and ADP-bound actin is depolymerized at the slow-growing (also called pointed) ends of filaments. (B) Formin proteins and spire are important actin nucleators for the generation of actin filaments in oocytes. The domain architecture of canonical formin proteins (left). The proline-rich FH1 domain recruits profilin-actin complexes and directs them to the FH2 domain, which is responsible for the actin nucleation/elongation activity of the protein. Formins are usually regulated by an interaction between the C-terminal domain (CD) and the N-terminal regulatory domain. CD is essential for FMN2 catalytic activity and interaction with spire.154 The domain architecture of spire (right). Four tandem WH2 domains (marked as W) are responsible for binding with monomeric actin. Spire interacts with formin proteins via the N-terminal KIND.54 The putative Rab-binding domain56 (Spire-box, marked as S) and a modified FYVE domain,155 which usually target the protein to the membrane, are located in the CD. In mammalian oocytes, FMN2 interacts and cooperates with spire1/239 and is responsible for outer cortical actin and cytoplasmic actin mesh.12 (C) The ARP2/3 complex is a 7-protein complex responsible for the generation of branched actin.63 This complex contains 2 actin-related proteins (ARP2 and ARP3). Two NPFs recruit monomeric actin and bind to the ARP2/3 complex, activating ARP proteins.156,157 (D) Domain organization of typical NPFs. Regulatory domains can bind various GTPases, including Rac1 in the case of WAVEs158,159 and cdc42 in the case of n-WASP.160 Regulatory regions can also be involved in the localization of NPFs and actin polymerization, such as in the case of WHAMM161 or WASH.162 All known NPFs contain C-terminal ‘W-C-A’ motifs consisting of WH2 (marked as W), cofilin homology (C), and acidic (A) domains. The WH2 motif is responsible for monomeric actin recruitment, whereas ‘C-A’ motifs are required for ARP2/3 binding. Illustrations were adapted from Dominguez and Namgoong163 and modified to accommodate more recent findings.