
FIGURE 2.
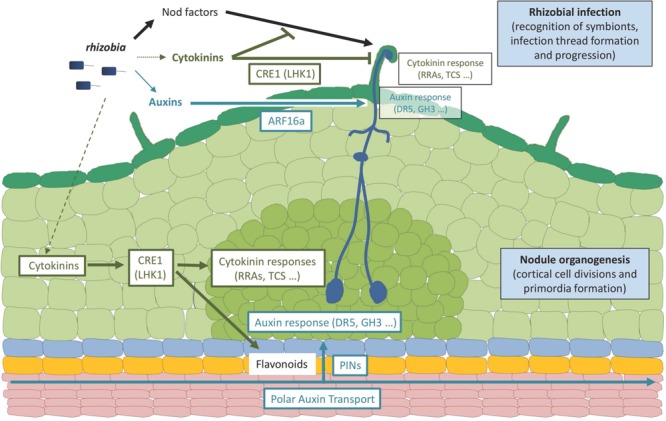
Roles of auxins and cytokinins in legume-rhizobia symbiotic interactions. The scheme represents an early rhizobial infection event in a legume (Medicago truncatula) root leading to the formation of an indeterminate nodule. Rhizobia produce specific Nod factor signaling molecules as well as auxin and cytokinin hormones. A major contribution of Nod factor signals in establishing rhizobial infection events (i.e., symbiont recognition, infection thread formation and progression) in root hairs has been demonstrated, whereas cytokinins produced by rhizobia may only have a minor contribution, and a role for a WT-level bacterial production of auxins remains to be established. Plant and/or bacterial hormones lead to the activation of cytokinin and auxin responses in infected epidermal cells, respectively, monitored by Type A RRs (RRA) genes or the TCS (Two Component output Sensor) cytokinin-response marker and the GH3 (Gretchen Hagen3) auxin responsive gene or the DR5 auxin-response marker. The activation of cytokinin responses in the root epidermis may lead to an inhibition of Nod factor signaling and/or rhizobial infections whereas the activation of auxin responses may promote rhizobial infections. The activation of these root epidermal cytokinin responses depends on the CRE1 (Cytokinin Response 1) receptor (LHK1, standing for Lotus Histidine Kinase1, being its functional homolog in Lotus japonicus) whereas the activation of the auxin epidermal responses depends on ARF16a (Auxin Response Factor 16a, a transcription factor). A cytokinin response is additionally activated in dividing cortical cells and nodule primordia, also depending on the CRE1 (LHK1) receptor, promoting nodule organogenesis even in the absence of symbiotic signals (rhizobia or Nod factors). An activation of auxin responses is observed in the cortex downstream of the CRE1-cytokinin pathway, involving local changes in polar auxin transport associated to the activation of specific flavonoid metabolite production and changes in PIN (PIN-formed) auxin-efflux carriers expression and accumulation. Ultimately, the activation of these auxin and cytokinin responses is linked to modulations of the cell cycle leading either to infection thread formation (epidermis) or cell divisions (cortex). It remains open if the auxin and/or cytokinin hormonal pools may, in addition to originate from plants and/or bacteria, accumulate upon rhizobial/Nod factors activation specifically in the infected epidermis, in the activated cortex, or in both root tissues. In addition, divergences in these hormonal responses are observed between determinate and indeterminate nodule types (e.g., L. japonicus versus M. truncatula model legumes), notably regarding the role of CRE1 versus LHK1 cytokinin receptors in regulating infections in the epidermis, the involvement of polar auxin transport in root inner tissues, and the contribution of a local auxin biosynthesis in these different root tissues. Dark green: root hairs and epidermis; pale green, cortex; middle green, nodule primordium; blue, endodermis; yellow, pericycle; pink, stele tissues including vascular bundles.