
Figure 2. Different programming protocols and shape-recovery pathways.

(a) Three different shape programming protocols produce identical deformation states of a dual network (uniaxial extension, ɛ=0.5): (i) Deformation at different strain rates as indicated (solid lines, T=25 °C), (ii) Stress relaxation for controlled times (dotted line, ɛ=0.5, 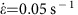 , 25 °C), and (iii) Deformation at a higher temperature (dashed line, 37 °C). Top panel: network re-arrangement at different stages of stress relaxation (–). The simple shear schematic is a conventional way for picturing deformation under constant volume conditions. The solid and dashed ovals correspond to the original and re-associated H-bonds, respectively. During shape recovery, the strained chemical network drives reverse dissociation/association of these transient H-bonds towards the original shape. (b) A triple hairpin demonstrates the ability to control both rate and pathway of shape recovery by adjusting the programming time (tp) of the individual angles. Upper panel: all three angles were programmed identically (Protocol 2: tp=100 min) and therefore, unfold simultaneously. Lower panel: The angles were held for different times ((1):1 min, (2):10 min, (3):100 min), which resulted in sequential unfolding with shorter tp resulting in faster recovery. Both programming and recovery were conducted under the same conditions (oil, T=22 °C). (c) Reversible shapeshifting of a hairpin was programmed in two steps: (i) folding and stress relaxation for 10 min, followed by (ii) unfolding and stress relaxation for 1 min. After release of the external force, the hairpin undergoes shape recovery through folding-unfolding motions (Supplementary Movie 1) under constant surrounding conditions (T=22 °C). All samples are 50:50 MAAc-co-DMAA gel (70 wt% water). To control water evaporation, all deformation and shape-recovery tests were conducted in either silicone oil or pH 3 water (Supplementary Fig. 10).
, 25 °C), and (iii) Deformation at a higher temperature (dashed line, 37 °C). Top panel: network re-arrangement at different stages of stress relaxation (–). The simple shear schematic is a conventional way for picturing deformation under constant volume conditions. The solid and dashed ovals correspond to the original and re-associated H-bonds, respectively. During shape recovery, the strained chemical network drives reverse dissociation/association of these transient H-bonds towards the original shape. (b) A triple hairpin demonstrates the ability to control both rate and pathway of shape recovery by adjusting the programming time (tp) of the individual angles. Upper panel: all three angles were programmed identically (Protocol 2: tp=100 min) and therefore, unfold simultaneously. Lower panel: The angles were held for different times ((1):1 min, (2):10 min, (3):100 min), which resulted in sequential unfolding with shorter tp resulting in faster recovery. Both programming and recovery were conducted under the same conditions (oil, T=22 °C). (c) Reversible shapeshifting of a hairpin was programmed in two steps: (i) folding and stress relaxation for 10 min, followed by (ii) unfolding and stress relaxation for 1 min. After release of the external force, the hairpin undergoes shape recovery through folding-unfolding motions (Supplementary Movie 1) under constant surrounding conditions (T=22 °C). All samples are 50:50 MAAc-co-DMAA gel (70 wt% water). To control water evaporation, all deformation and shape-recovery tests were conducted in either silicone oil or pH 3 water (Supplementary Fig. 10).