

Abstract
Categorization’s great debate has weighed single-system exemplar theory against the possibility of alternative processing systems. We take an evolutionary perspective toward this debate to illuminate it in a new way. Animals are crucial behavioral ambassadors to this area. They reveal the roots of human categorization, the basic assumptions of vertebrates entering category tasks, and the surprising weakness of exemplar memory as a category-learning strategy. These results have joined neuroscience results to prompt important changes in categorization theory. Categorization’s great debate is ending. Categorization is served by multiple systems of process and representation.
Keywords: learning, categorization, cognitive evolution, comparative cognition
1. INTRODUCTION
Categorization—the ability of humans and animals to learn psychological equivalence classes (categories)—is a crucial survival capacity. It lets us differentiate mushrooms from toadstools, garden snakes from rattlesnakes. It has been a vertebrate life preserver for hundreds of millions of years. For example, perhaps half the mortality among vervet monkeys (Cercopithecus aethiops) is due to predators (Cheney & Seyfarth, 1990), especially eagles. Vervets have developed an eagle warning call—it communicates the presence of an eagle to conspecifics and in this sense “names” a member of the category eagle (Figure 1).
Figure 1.

Eat, prey, love. The lives of vertebrates depend on identifying, categorizing, and naming natural kinds—foods, mates, especially predators. Photo credit: Melanie Stetson Freeman/The Christian Science Monitor via Getty Images (Top); Phineas Mufwaya (Middle); Dan Heller (Bottom).
For decades, the categorization literature had the dominant theoretical narrative that one representational/processing system serves all of categorization. The narrative was that organisms store exemplars as separate, individuated memory traces, refer new items to these stored exemplars, and include them in the category if they are similar enough. Thus, exemplar theory portrayed category representation as a slide carousel or Rolodex of instances. This narrative made categorization unitary and parsimonious. It made all categories potentially learnable (even random assemblages of objects would be memorizeable as instances). It made categorization science “easy,” with just one process in mind and memory to understand. Readers may see already how this article will unfold.
This determined parsimony produced our literature’s great debate—about alternative processes and multiple systems in categorization. Other literatures shared this unitarian impulse (e.g., imagery, which doggedly pursued unitary-code theory; memory, which sharply doubted dissociable memory systems). The search for parsimony runs deep in cognitive science, a crucial topic itself but not ours. Rather, this article summarizes recent research to make several points:
Animals are important behavioral ambassadors to categorization science. They illuminate the evolutionary roots of human categorization and they correct theory.
There are continuities in category processes—extending across millions of years in vertebrate evolution—that have profound theoretical implications.
The continuities could illuminate the structure of natural kinds.
The perspectives of fitness, adaptation, and evolution could explain the continuities.
The focus on categorization’s natural history has been part of a recent transformation in categorization theory.
Categorization’s great debate is ending. Broad-based converging evidence now makes it clear that the unitary exemplar view is insufficient. Categorization is served by multiple systems of process/representation.
2. EXEMPLAR PROCESSES INSUFFICIENT
Exemplar theory holds that categorizers store exemplars as separate memory traces spread out like a cloud in the mind’s psychological space. Then potential instances are endorsed as category members if they share sufficient summed similarity to these multiple cognitive reference points. Thus, exemplar theory makes this prediction: even a perfectly typical item (a category prototype) may not be maximally endorsed into the category, because it still cannot be similar to all the stored exemplars. It will always be close (similar) to some but far (dissimilar) from others (Smith, 2002).
Testing this prediction, Smith, Redford, and Haas (2008) let macaques (Macaca mulatta) learn categories containing central prototypes and related (typical) items. They adopted the influential dot-distortion task from human research (Knowlton & Squire, 1993; Posner & Keele, 1968; Smith & Minda, 2001). These categories were perceptually coherent, with category members mutually similar. Such categories are frequently called family-resemblance categories, recalling Rosch’s pioneering work (Rosch & Mervis, 1975).
Figure 2 (circles) shows the proportion of times a macaque endorsed different item types into the category. He maximally endorsed the prototype. An exemplar model instantiated exemplar processing by a hypothetical monkey mind, but its predictions (Es) were poor (e.g., it seriously underpredicted performance on the most typical category members). Exemplar theory’s psychology of categorization forced predictions that macaques strikingly disconfirmed.
Figure 2.

The proportion of times a macaque endorsed into a learned category items that were outside the category (Rand.), and low typical (Lo. Typical), typical, high typical (Hi. Typical), and prototypical category members (Prot.). Decreasing typicality was a function of increasing shape distortion from the prototype. Exemplar and prototype models fit the macaque’s performance as well as they could (E and P symbols, respectively). The models differed only in assuming multiple, specific-exemplar cognitive reference points for categorization or a single, central cognitive reference point. Adapted with permission from “Prototype abstraction by monkeys (Macaca mulatta),” by J. D. Smith, J. S. Redford, and S. M. Haas, Journal of Experimental Psychology: General, 137, 390–401. Copyright 2008 by the American Psychological Association.
Perhaps instead, macaques blended their exemplar experience into the category’s prototype. A prototype model instantiated prototype processing by a hypothetical monkey mind. It was equivalent to the exemplar model save only its representational assumption (i.e., the prototype as sole cognitive reference point). Its predictions (Ps) were so good that the P symbols in Figure 2 are difficult to see. Something like a prototype was the monkey’s cognitive reference point, a conclusion reinforced by other studies (Aydin & Pearce, 1994; Huber & Lenz, 1993; Jitsumori, 1996; von Fersen & Lea, 1990; White, Alsop, & Williams, 1993). Exemplar processing—missing in action here—is insufficient to explain all of what categorizing animals do, or humans (Smith, 2002; Smith & Minda, 2001, 2002), though still it could explain some of what they do.
3. THE FAMILY-RESEMBLANCE ASSUMPTION
One can show animals’ natural preference for something like prototype processing with a prototype-exception task (Figure 3). The shapes on the left are all Category As, though the last two (the exceptions) don’t look it. The same is true with the Category B shapes on the right. If animals store exemplars, they will memorize and categorize correctly typical and exception items. But not if they assume family-resemblance or prototype-based categories. Then they will call the leftward exceptions Bs and the rightward exceptions As. If we observe criss-crossed exception performance, we will know they made the family-resemblance assumption.
Figure 3.

Examples of categories and stimuli used in Smith et al. (2010). The 8-shape groupings are two categories A and B. The 6 shapes in the top rows of each category are variations on the theme of the category prototype. The bottom row contains exception items that are variations on the theme of the opposing prototype. From “Stages of category learning in monkeys (Macaca mulatta) and humans (Homo sapiens),” by J. D. Smith, W. P. Chapman, and J. S. Redford, Journal of Experimental Psychology: Animal Behavior Processes, 36, 39–53. Copyright 2010 by the American Psychological Association. Reprinted with permission.
Figure 4a–c shows the macaques’ result (Smith, Chapman, & Redford, 2010). They persistently criss-crossed the exceptions, even after hundreds of tries at them. There were only four exceptions to learn! They remained at or below chance on the exceptions for about 12,000, 6,000, and 3,000 trials respectively. During this period, they showed an amazingly poor ability to remember and respond to individuated exemplars. If they were coping with exceptional eagles in the world, they might not last ‘til lunch. The macaques assumed these are coherent family-resemblance categories. They defaulted to something like prototype processing. To flip the exceptions is correct under the family-resemblance assumption. Humans show similar phenomena (Smith, Murray, & Minda, 1997; Smith & Minda, 1998; Smith et al., 2010). Where is the exemplar processing?
Figure 4.

A-C. The performance of monkeys Hank, Lou, and Murph by 64-trial block in the category task of Smith, Chapman, and Redford (2010). Curves T and E, respectively, show the proportion of correct responses made to the six typical and two exception items in each category. From “Stages of category learning in monkeys (Macaca mulatta) and humans (Homo sapiens),” by J. D. Smith, W. P. Chapman, and J. S. Redford, Journal of Experimental Psychology: Animal Behavior Processes, 36, 39–53. Copyright 2010 by the American Psychological Association. Reprinted with permission.
It is there. Two macaques finally somewhat learned the exceptions after thousands of trials (Figure 4b,c). We are not excluding exemplar processing from an overall theory. We are limiting its role. We are showing why a multiple-systems theoretical account is essential to the field’s development.
The animal’s assumption entering a task is a crucial theoretical fact about its mind, no matter what it does after thousands of trials. It shows what the animal assumes about how the world is. It limits the role of exemplar processing—here, to the task’s ultimate trials. But these trials have no relevance to categorization’s natural cognitive function. The reason?: if you make hundreds of mistakes on eagles early on, you never reach the ultimate trials. Though psychologists deem the ultimate trials “terminal performance,” they are too late. For nature has already deemed the early trials “terminal performance” in a deadlier sense. Categorization researchers must stop confusing ultimate performance with normal performance. The latter has deeper relevance.
4. BROAD EVOLUTIONARY CONTINUITIES
The family-resemblance assumption may reach far across vertebrate evolution. Cook and Smith (2006) gave pigeons (and humans) a prototype-exception task (Figure 5). The prototypes (Row 1) and typical items (Rows 2–6) have 6 and 5 typical colors, respectively. The exceptions (Row 7) look like they “should” belong across the aisle.
Figure 5.

Illustrating the prototype-exception task used to test pigeons and humans in Cook and Smith (2006). Each category contained a prototype (Row 1), five typical stimuli (Rows 2–6—5 features shared with their prototype), and an exception (Row 7—5 features shared with the opposing prototype). From “Stages of abstraction and exemplar memorization in pigeon category learning,” by R. G. Cook and J. D. Smith, 2006, Psychological Science, 17, p. 1059–1067. Copyright 2006 by the Association for Psychological Science. Reprinted with permission.
The black circles in Figure 6 (top panels) show early learning in this task. Both species showed strong prototype performance but criss-crossed exception performance. The triangles show how a hypothetical exemplar processor would perform. Its predictions are poor. The squares show how a hypothetical prototype processor would perform. Its predictions are good. Something like prototype processing is occurring. Wasserman, Kiedinger, and Bhatt (1988) showed a similar phenomenon in pigeons. Later on (Figure 6, bottom panels), dogged training let pigeons ultimately classify exceptions. Nonetheless, it is theoretically important that an initial family-resemblance assumption extends across many evolutionary lines, and probably back through many millions of years of vertebrate evolution. Why? (Sections 6,7).
Figure 6.

Humans’ and pigeons’ observed and predicted accuracy for prototypes, typical items, and exceptions during the early (top panels) and late (bottom panels) stages of learning within the prototype-exception task of Cook and Smith (2006). Observed performances are shown as filled, black circles. The best-fitting predictions of prototype and exemplar models are shown, respectively, as open squares and triangles. From “Stages of abstraction and exemplar memorization in pigeon category learning,” by R. G. Cook and J. D. Smith, 2006, Psychological Science, 17, p. 1059–1067. Copyright 2006 by the Association for Psychological Science. Reprinted with permission.
5. FAILED EXEMPLAR PROCESSING—STILL
Perhaps family-resemblance categories (Sections 3,4) tempt macaques toward family-resemblance assumptions, so their exemplar processing cannot shine through. Smith, Coutinho, and Couchman (2011) explored this using an exclusive-or (XOR) task. In an XOR task (e.g., Category A items: Black Circles, White Squares; Category B items: White Circles, Black Squares), no shape or color cue discriminates the categories. There is no prototype to learn or family-resemblance assumption to make. Everything is individually exceptional, recommending exemplar memory to map four responses to four stimuli.
Macaques found the XOR tasks terribly difficult. They missed a third of all trials. They received more than 15,000 20-s timeout penalties for errors (300,000 s, equivalent to 9 full seasons of Dr. Who). Asymptotic performance was poor—about 75% correct after 5,800 trials—though the stimuli repeated hundreds of times.
XOR tasks have seemed to provide strong evidence for a unitary exemplar system in categorization (Medin, Altom, Edelson, & Freko, 1982). Yet macaques show extremely weak exemplar processing—even when that process seems mandatory. Perhaps there is some exemplar processing, but again not ecologically relevant or useful exemplar processing. Thus the monkeys answer a fundamental question. It is extremely unlikely that exemplar processing is their unitary or primary mode of category learning. Applied in nature, it would be a fitness disaster.
However, we acknowledge that animals may sometimes need specific-exemplar Rolodexes, especially when categories are populated with few, disparate, recurring category exemplars as in the XOR task. For example, specific-exemplar memory for individuals could be useful for managing dominance relations in a primate troop. Exemplar memory may have a place as part of an overall categorization system. Prototypes do as well. So do explicit category rules, as another influential literature makes clear (e.g., Ashby & Maddox, 2010). A good theoretical starting point is to note that category representations are memories—multiple categorization systems could map onto multiple memory processes (e.g., exemplar categorization onto episodic memory).
Humans also perform terribly in weak family-resemblance tasks that enforce exemplar processing. For example, in many experiments by Medin and colleagues, about half of all participants never reached the learning criterion (e.g., Medin & Schwanenflugel, 1981; Medin & Schaffer, 1978). To give due credit, Murphy (2003, 2005) warned human categorization researchers of this problem. These tasks probably take humans outside their family-resemblance comfort zone and force them toward weak exemplar processes instead. We must not mistake performance in artificial, barely solvable tasks for performance in ecological, normal tasks. The latter has deeper relevance.
6. WHY PROTOTYPES; WHY FAMILY-RESEMBLANCE ASSUMPTIONS?
Smith (2014) addressed these questions through a distinctive application of formal models. He used exemplar and prototype models to ask what strategies let organisms perform optimally (also Briscoe & Feldman, 2011). This issue has received scant attention. The reason is the dominant narrative of one processing system to rule all categorization. If there aren’t different processes, the optimality question vanishes. The dominant narrative has sometimes done harm.
Smith’s simulations showed that prototype processing in family-resemblance and prototype-exception tasks produces a meaningful performance advantage of about 4% over exemplar processing. This advantage would allow for improved prey and predator recognition, increased foraging efficiency, and increased fitness. Why is the prototype often the best cognitive reference point?
The prototype is all category signal, no noise. Specific exemplars have misleading, idiosyncratic features (Figure 7). They are signal and noise—weaker signals overall. Geometrically, the central prototype is the closest reference point in psychological space to the most members of the category. It is the category problem’s least-squares solution. Briscoe and Feldman (2011) noted that stored exemplars can be too specific, undermining successful generalization. Ashby & Alfonso-Reese (1995) noted that estimating the statistical distribution of exemplars can be the hardest aspect of learning a category. But if the family-resemblance distribution kept recurring, organisms would gain if they pre-assumed that category structure. This makes it especially intriguing that pigeons, macaques, and humans make this family-resemblance pre-assumption.
Figure 7.

Monkeys! Specific exemplars often have idiosyncratic, distracting features that can make them misleading cognitive reference points for use in categorization. Photo credit: Jef Russell (Top), Frederick Dunn of Suspended Moments (Middle), © Can Stock Photo Inc. / kjorgen (Bottom).
7. POSSIBLE WORLDS: WHAT KINDS OF NATURAL KINDS?
Prototype processing has a downside. Smith’s (2014) formal simulations confirmed that exemplar processing is far superior to prototype processing in XOR tasks and whenever a task defeats prototype processing. Then organisms would need robust exemplar memory to establish cognitive reference points. So it matters what world animals live in. Consider the relative plausibility of two possible worlds.
The world of natural kinds might be full of XORness and other prototype-defeating categories, like the ecological XOR task shown in Figure 8. From Smith et al. (2011), we know that monkeys would pick deadly fruit every day (or, perhaps only for one day!). If the world were persistently exemplar demanding like that, animals would likely have evolved to be exemplar adept, unless exemplar processing is computationally intractable which no exemplar theorist would accept. That they’re not exemplar adept suggests the world is not exemplar demanding like that. It isn’t XOR filled.
Figure 8.
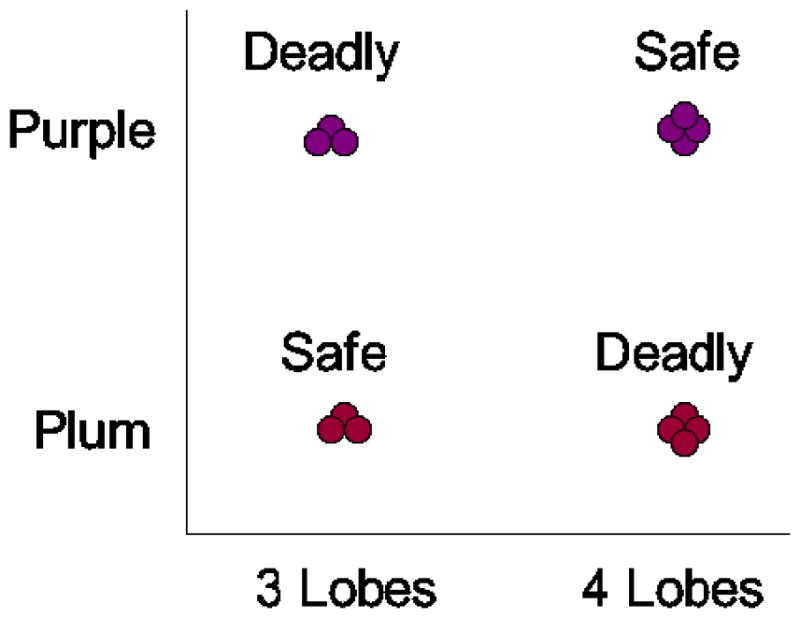
A hypothetical ecological exclusive-or (XOR) category task. To our knowledge, nature almost never presents this kind of foraging or predator-avoidance problem to living organisms, and it is a graceful state of nature that this is so.
But what if the world is mostly prototype averageable? Then prototype averaging would be adaptive. It might be selected for and developed during cognitive evolution, unless it is computationally intractable which the results reviewed here disconfirm. Animals would be attuned to prototypes. They are. They would pre-assume family-resemblance structure. They do. They could be poor with exemplars. They are. So would their human descendants. They are. This world is much more plausible.
This conclusion is very bad for unitary exemplar theory. But its defenders have this response: The logical universe of possible categories is almost all poorly structured categories with weak family resemblance. Family-resemblance categories are a sliver in its overall Venn diagram. How can it be that animals are adept at a sliver of the tasks they might have to face? It’s an impossible coincidence and constraint. It would be like babies being born prepared only to speak Basque.
This is how it can be. The logical universe is irrelevant; only the ecological world is relevant. There, by hypothesis, family-resemblance categories dominate. Animals are adept at what they experience and what they need. Their adeptness is neither a coincidence nor a disability.
It is clear—looking at real monkey categories—that the world does not give nasty XOR berry tasks. It gives coherent family-resemblance categories—insects, snakes, leopards, eagles, mates, and Masai. Ashby (1992) agreed that many natural kinds have this structure. Fried and Holyoak (1986, p. 235) suggested organisms might safely have this general expectation. Rosch (1973, 1975) famously pointed out the family-resemblance structure of natural kinds and its psychological importance. Ethnobiologists confirm that many biological kinds have a family-resemblance structure with psychological privilege for humans worldwide. Malt (1995) contributed an extraordinary review of this extensive literature.
So family-resemblance categories may have played a role in categorization’s evolution. Their processing advantages would have been available through the many millions of years that animals confronted category problems. There may have been a gentle shaping pressure—phylogeny long—toward family-resemblance pre-assumptions. This could be an example of evolutionary tuning by which animals became cognitively adept within their natural world (also Anderson, 1991; Shepard, 1987, 2001). In the end, monkeys may be making the most important theoretical statement of all: that vertebrates have not been doing unitary exemplar processing for half a billion years.
They are not alone in challenging categorization’s long-dominant theoretical narrative. Many other lines of research also demonstrate multiple, dissociable systems of category learning (reviews in Ashby, 2013; Ashby & Maddox, 2010; Seger & Miller, 2010). This large body of evidence complements that considered here. It would need another article to do it justice. Thus, abundant, broadly converging evidence makes it clear that categorization is served by multiple systems of process and representation. Animals have made a distinctive contribution to this area. Our literature’s great debate is ending, and the monkeys are chattering the news through all the jungles of cognitive science.
Footnotes
The preparation of this article was supported by Grant HD061455 from NICHD, Grant BCS-0956993 from NSF, and Grant HD060563 from NICHD.
Contributor Information
J. David Smith, Department of Psychology, Georgia State University.
Alexandria C. Zakrzewski, Department of Psychology, University at Buffalo, The State University of New York
Jennifer M. Johnson, Department of Psychology, Georgia State University
Jeanette C. Valleau, Department of Psychology: Georgia State University
References
- Anderson JR. The adaptive nature of human categorization. Psychological Review. 1991;98:409–429. [Google Scholar]
- Ashby FG. Multidimensional models of categorization. In: Ashby FG, editor. Multidimensional models of perception and cognition. Hillsdale, NJ: Erlbaum; 1992. pp. 449–483. [Google Scholar]
- Ashby FG. Human Category Learning, Neural Basis. In: Pashler H, editor. The Encyclopedia of the Mind. Thousand Oaks, CA: Sage Publishing; 2013. [Google Scholar]
- Ashby FG, Alfonso-Reese LA. Categorization as probability density estimation. Journal of Mathematical Psychology. 1995;39:216–233. [Google Scholar]
- Ashby FG, Maddox WT. Human category learning 2.0. Annals of the New York Academy of Sciences. 2010;1224:147–161. doi: 10.1111/j.1749-6632.2010.05874.x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Aydin A, Pearce JM. Prototype effects in categorization by pigeons. Journal of Experimental Psychology: Animal Behavior Processes. 1994;20:264–277. [Google Scholar]
- Briscoe E, Feldman J. Conceptual complexity and the bias/variance tradeoff. Cognition. 2011;118:2–16. doi: 10.1016/j.cognition.2010.10.004. [DOI] [PubMed] [Google Scholar]
- Cheney DL, Seyfarth RM. How monkeys see the world. Chicago: University of Chicago Press; 1990. [Google Scholar]
- Cook R, Smith JD. Stages of abstraction and exemplar memorization in pigeons’ category learning. Psychological Science. 2006;17:1059–1067. doi: 10.1111/j.1467-9280.2006.01833.x. [DOI] [PubMed] [Google Scholar]
- Fried LS, Holyoak KJ. Induction of category distributions: A framework for classification learning. Journal of Experimental Psychology: Learning, Memory, and Cognition. 1984;10:234–257. doi: 10.1037//0278-7393.10.2.234. [DOI] [PubMed] [Google Scholar]
- Huber L, Lenz R. A test of the linear feature model of polymorphous concept discrimination with pigeons. Quarterly Journal of Experimental Psychology. 1993;46B:1–18. [Google Scholar]
- Jitsumori M. A prototype effect and categorization of artificial polymorphous stimuli in pigeons. Journal of Experimental Psychology: Animal Behavior Processes. 1996;22:405–419. doi: 10.1037//0097-7403.22.4.405. [DOI] [PubMed] [Google Scholar]
- Knowlton BJ, Squire LR. The learning of categories: Parallel brain systems for item memory and category knowledge. Science. 1993;262:1747–1749. doi: 10.1126/science.8259522. [DOI] [PubMed] [Google Scholar]
- Malt BC. Category coherence in cross-cultural perspective. Cognitive Psychology. 1995;29:85–148. [Google Scholar]
- Medin DL, Altom MW, Edelson SM, Freko D. Correlated symptoms and simulated medical classification. Journal of Experimental Psychology: Learning, Memory, and Cognition. 1982;8:37–50. doi: 10.1037//0278-7393.8.1.37. [DOI] [PubMed] [Google Scholar]
- Medin DL, Schaffer MM. Context theory of classification learning. Psychological Review. 1978;85:207–238. [Google Scholar]
- Medin DL, Schwanenflugel PJ. Linear separability in human classification learning. Journal of Experimental Psychology: Human Learning & Memory. 1981;7:355–368. [Google Scholar]
- Murphy GL. Ecological validity and the study of concepts. In: Ross BH, editor. The psychology of learning and motivation. Vol. 43. San Diego: Academic Press; 2003. pp. 1–41. [Google Scholar]
- Murphy GL. The study of concepts inside and outside the laboratory: Medin versus Medin. In: Ahn W, Goldstone RL, Love BC, Markman AB, Wolff P, editors. Categorization inside and outside the laboratory: Essays in honor of Douglas L. Medin. Washington, DC: APA; 2005. pp. 179–195. [Google Scholar]
- Posner MI, Keele SW. On the genesis of abstract ideas. Journal of Experimental Psychology. 1968;77:353–363. doi: 10.1037/h0025953. [DOI] [PubMed] [Google Scholar]
- Rosch E. On the internal structure of perceptual and semantic categories. In: Moore TE, editor. Cognitive development and the acquisition of language. New York: Academic Press; 1973. pp. 111–144. [Google Scholar]
- Rosch E. Cognitive representations of semantic categories. Journal of Experimental Psychology: General. 1975;104:192–233. [Google Scholar]
- Rosch E, Mervis CB. Family resemblances: Studies in the internal structure of categories. Cognitive Psychology. 1975;7:573–605. [Google Scholar]
- Seger CA, Miller EK. Category learning in the brain. Annual review of neuroscience. 2010;33:203. doi: 10.1146/annurev.neuro.051508.135546. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Shepard RN. Toward a universal law of generalization for psychological science. Science. 1987;237(4820):1317–1323. doi: 10.1126/science.3629243. [DOI] [PubMed] [Google Scholar]
- Shepard RN. Perceptual-cognitive universals as reflections of the world. Behavioral and Brain Sciences. 2001;24:581–601. [PubMed] [Google Scholar]
- Smith JD. Exemplar theory’s predicted typicality gradient can be tested and disconfirmed. Psychological Science. 2002;13:437–442. doi: 10.1111/1467-9280.00477. [DOI] [PubMed] [Google Scholar]
- Smith JD. Prototypes, exemplars, and the natural history of categorization. Psychonomic Bulletin and Review. 2014;21:312–331. doi: 10.3758/s13423-013-0506-0. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Smith JD, Minda JP. Prototypes in the mist: the early epochs of category learning. Journal of Experimental Psychology: Learning, Memory, and Cognition. 1998;24:1411–1436. [Google Scholar]
- Smith JD, Minda JP. Journey to the center of the category: The dissociation in amnesia between categorization and recognition. Journal of Experimental Psychology: Learning, Memory, and Cognition. 2001;27:984–1002. doi: 10.1037//0278-7393.27.4.984. [DOI] [PubMed] [Google Scholar]
- Smith JD, Minda JP. Distinguishing prototype-based and exemplar-based processes in category learning. Journal of Experimental Psychology: Learning, Memory, and Cognition. 2002;28:800–811. [PubMed] [Google Scholar]
- Smith JD, Ashby FG, Berg ME, Murphy MS, Spiering B, Cook RG, Grace RC. Pigeons’ categorization may be exclusively nonanalytic. Psychonomic Bulletin and Review. 2011;18:422–428. doi: 10.3758/s13423-010-0047-8. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Smith JD, Beran MJ, Crossley MJ, Boomer J, Ashby FG. Implicit and explicit category learning by macaques (Macaca mulatta) and humans (Homo sapiens) Journal of Experimental Psychology: Animal Behavior Processes. 2010;36:54–65. doi: 10.1037/a0015892. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Smith JD, Chapman WP, Redford JS. Stages of category learning in monkeys (Macaca mulatta) and humans (Homo sapiens) Journal of Experimental Psychology: Animal Behavior Processes. 2010;36:39–53. doi: 10.1037/a0016573. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Smith JD, Coutinho MVC, Couchman JJ. The learning of exclusive-or categories by monkeys (Macaca mulatta) and humans (Homo sapiens) Journal of Experimental Psychology: Animal Behavior Processes. 2011;37:20–29. doi: 10.1037/a0019497. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Smith JD, Murray MJ, Minda JP. Straight talk about linear separability. Journal of Experimental Psychology: Learning, Memory, and Cognition. 1997;23:659–680. [Google Scholar]
- Smith JD, Redford JS, Haas SM. Prototype abstraction by monkeys (Macaca mulatta) Journal of Experimental Psychology: General. 2008;137:390–401. doi: 10.1037/0096-3445.137.2.390. [DOI] [PubMed] [Google Scholar]
- Von Fersen L, Lea SEG. Category discrimination by pigeons using five polymorphous features. Journal of the Experimental Analysis of Behavior. 1990;54(2):69–84. doi: 10.1901/jeab.1990.54-69. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Wasserman EA, Kiedinger RE, Bhatt RS. Conceptual behavior in pigeons: categories, subcategories, and pseudocategories. Journal of Experimental Psychology: Animal Behavior Processes. 1988;14:235–246. [Google Scholar]
- White KG, Alsop B, Williams L. Prototype identification and categorization of incomplete figures by pigeons. Behavioural Processes. 1993;30:253–258. doi: 10.1016/0376-6357(93)90137-G. [DOI] [PubMed] [Google Scholar]
Recommended Readings
- Ashby FG, Maddox WT. Human category learning 2.0. Annals of the New York Academy of Sciences. 2010;1224:147–161. doi: 10.1111/j.1749-6632.2010.05874.x. A description of the cognitive-neuroscience research that complements the research discussed in this article, from premier researchers in that area. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Malt BC. Category coherence in cross-cultural perspective. Cognitive Psychology. 1995;29:85–148. A beautiful review of the enormous literature on natural kinds and categories, and systems of folk biology and fold taxonomy worldwide. [Google Scholar]
- Murphy GL. The big book of concepts. Cambridge, MA: MIT Press; 2003. The book on psychological theories of concepts and categories. [Google Scholar]
- Smith JD. Prototypes, exemplars, and the natural history of categorization. Psychonomic Bulletin and Review. 2014;21:312–331. doi: 10.3758/s13423-013-0506-0. An extensive formal-simulation study of the issues discussed in this article. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Smith JD, Redford JS, Haas SM. Prototype abstraction by monkeys (Macaca mulatta) Journal of Experimental Psychology: General. 2008;137:390–401. doi: 10.1037/0096-3445.137.2.390. An in-depth demonstration of monkeys’ prototype-based category learning, with careful formal comparisons between exemplar and prototype processing. [DOI] [PubMed] [Google Scholar]