
Figure 5. Rate of EF-Tu dissociation influences the proofreading factor Pf.

(a) Pf is shown as a function of  for the induced fit mechanism of proofreading (Fig. 4c). For this calculation, accommodation of cognate tRNA is modelled as being 50-fold faster than near-cognate:
for the induced fit mechanism of proofreading (Fig. 4c). For this calculation, accommodation of cognate tRNA is modelled as being 50-fold faster than near-cognate: 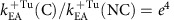 . Only
. Only 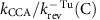 =10 is shown since Pf is only sensitive to this ratio when it is <1. (b) For the EA-destabilized mode of proofreading (Fig. 4d), Pf depends on the relative rates of EF-Tu dissociation
=10 is shown since Pf is only sensitive to this ratio when it is <1. (b) For the EA-destabilized mode of proofreading (Fig. 4d), Pf depends on the relative rates of EF-Tu dissociation 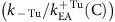 and 3′-CCA accommodation
and 3′-CCA accommodation  . Each curve is labelled by the value of the relative rate of 3′-CCA accommodation (
. Each curve is labelled by the value of the relative rate of 3′-CCA accommodation (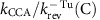 ). A representative value of ΔΔFNC=8 kBT was used to demonstrate the overall behaviour of Pf. In contrast to the induced fit description, proofreading through EA destabilization depends on the value of kCCA. This is because, when tRNA is in the EA basin, the reverse step for near-cognate tRNA, with a rate of
). A representative value of ΔΔFNC=8 kBT was used to demonstrate the overall behaviour of Pf. In contrast to the induced fit description, proofreading through EA destabilization depends on the value of kCCA. This is because, when tRNA is in the EA basin, the reverse step for near-cognate tRNA, with a rate of 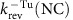 , will be competitive with forward accommodation (rate of kCCA). In contrast, the reverse barrier is large for all molecules in the induced-fit framework. Common to both descriptions is that fidelity increases monotonically with the rate of EF-Tu dissociation k−Tu. Solid (dashed) lines distinguish between regions where efficiency is greater (less) than 0.9. As the rate of EF-Tu dissociation increases, the efficiency decreases. This suggests that EF-Tu dissociation should be sufficiently slow, such that it may accelerate elbow accommodation (increase efficiency), yet fast enough to allow for rejection (increase selectivity).
, will be competitive with forward accommodation (rate of kCCA). In contrast, the reverse barrier is large for all molecules in the induced-fit framework. Common to both descriptions is that fidelity increases monotonically with the rate of EF-Tu dissociation k−Tu. Solid (dashed) lines distinguish between regions where efficiency is greater (less) than 0.9. As the rate of EF-Tu dissociation increases, the efficiency decreases. This suggests that EF-Tu dissociation should be sufficiently slow, such that it may accelerate elbow accommodation (increase efficiency), yet fast enough to allow for rejection (increase selectivity).