
Figure 2. New roles for non-coding RNAs.
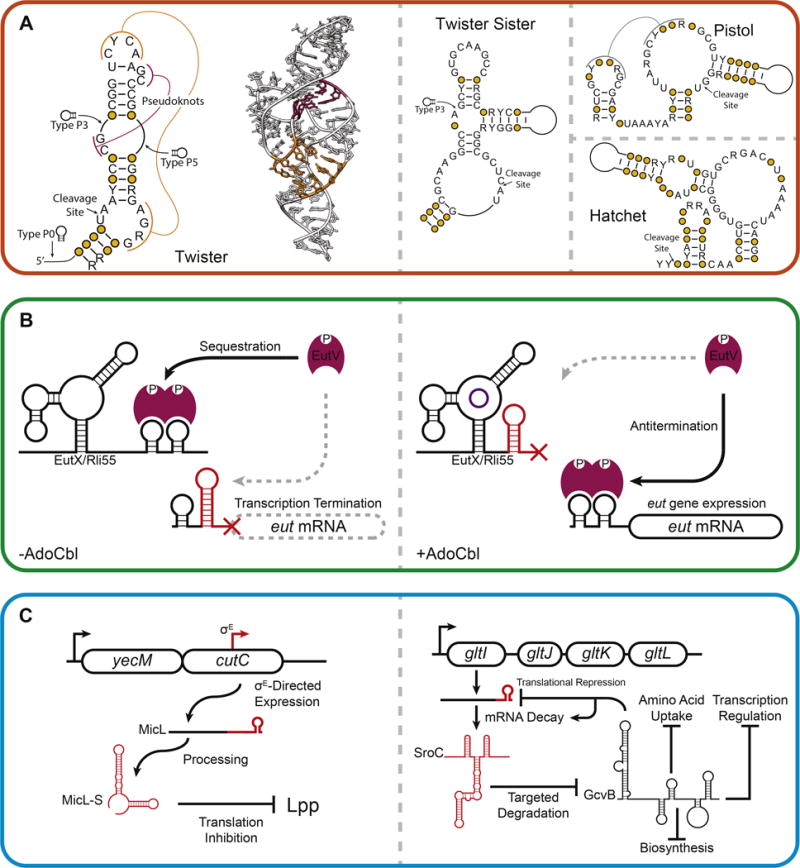
(A) Widespread identification of novel self-cleaving ribozymes. Secondary structures for the twister, twister sister, pistol, and hatchet ribozymes are shown [3,7]. A crystal structure of a twister ribozyme is shown (PDBID: 4QJH) [5]. Pseudoknot interactions are shown in magenta and orange in the twister ribozyme secondary structure and correspond to magenta and orange nucleotides in the twister ribozyme crystal structure. (B) An AdoCbl riboswitch regulates the expression of an sRNA sponge [16,17]. In the absence of AdoCbl, the full length EutX (or Rli55) sRNA is synthesized and sequesters the transcription antiterminator EutV. In the presence of AdoCbl, premature termination of EutX/Rli55 permits EutV to antiterminate transcription of the eut operon, leading to the expression of proteins involved in ethanolamine catabolism. (C) New roles for sRNAs in the global regulation of cellular processes. (Left) In response to outer membrane stress in γ-proteobacteria, the sRNA MicL is expressed from a σE-dependent promoter embedded in the cutC coding sequence and is processed into MicL-S, which inhibits translation of the outer membrane protein Lpp [26]. (Right) Decay of the gltI mRNA yields the sRNA SroC, which promotes degradation of the global regulatory sRNA GcvB, which regulates translation of the gltI mRNA [24].