

Abstract
Polymorphisms in the HOX transcript antisense intergenic RNA (HOTAIR) have been recently associated with susceptibility to different cancers. Here, a meta-analysis was performed to derive a more precise estimation of the involvement of HOTAIR polymorphisms in cancer development. Data from cases (n = 7,772) and controls (n = 9,075) were extracted from eligible studies (n = 10) identified in a comprehensive literature search conducted in PubMed, Embase, and the Web of Science databases through January 20, 2016. Overall, association between polymorphism rs920778 and increased cancer risk was significant in allele contrast (odds ratio (OR) = 1.239, 95% confidence interval (CI) = 1.032 - 1.487) and recessive models (OR = 1.614, 95% CI = 1.082 - 2.406). In subgroup analysis based on ethnicity, a significant association between polymorphism rs920778 and cancer susceptibility was observed in Asians under all models, but was most compelling under recessive (OR = 2.128, 95% CI = 1.417 - 3.197) and homozygous models (OR = 2.764, 95% CI = 2.221 - 3.440). Subgroup analysis by cancer type revealed a significant association between polymorphism rs4759314 and susceptibility to gastric cancer in allele contrast (OR = 1.262, 95% CI = 1.073 - 1.486), dominant (OR = 1.280, 95% CI = 1.060 - 1.547), and heterozygous models (OR = 1.288, 95% CI = 1.057 - 1.570). In conclusion, the results indicated that HOTAIR polymorphism rs920778 was more generally associated with cancer risk, particularly in Asians, while rs4759314 was a risk factor for gastric cancer.
Keywords: cancer, genetic susceptibility, HOTAIR, polymorphism
INTRODUCTION
Long non-coding RNAs (lncRNAs) are defined as transcribed RNA molecules that are longer than 200 nucleotides and not translated into proteins [1]. Although their function was initially unclear, lncRNAs are now known to have critical roles in the regulation of gene expression through transcription, processing of small RNAs, and epigenetic modification as well as other regulatory functions [2, 3]. As key components in gene regulatory complexes, lncRNAs contribute to the activation or inhibition of expression of genes involved in diverse normal cellular processes, such as proliferation and apoptosis. Many of these same processes are corrupted in cancer, and thus deregulated lncRNA expression has been linked to development of the disease [3–6].
Increasing evidence indicates that one of these molecules, HOX transcript antisense intergenic RNA (HOTAIR), has an oncogenic role in the development of human cancer in diverse tissues [5, 7, 8]. HOTAIR is a 2158-nucleotide lncRNA transcribed from the antisense strand of the HOXC gene which is located on chromosome 12 [9]. HOTAIR represses transcription, but surprisingly, in trans at the HOXD gene cluster on chromosome 2 [8, 10]. The lncRNA has been proposed to function as a molecular scaffold for the assembly of polycomb repressive complex 2 (PRC2) and lysine specific demethylase 1/REST corepressor 1/RE1-silencing transcription factor (LSD1/CoREST/REST) complex at 5′ and 3′ domains, respectively. Histone H3K27 methylation and H3K4 demethylation activities are thus effectively localized [11], which ultimately results in efficient chromosome condensation and transcriptional repression of targeted genes [7].
Numerous studies have demonstrated that overexpression of HOTAIR occurs in many cancers. It is furthermore associated with poor prognosis, and in experimental models, it promotes tumor progression, invasion, and metastasis [12–16]. Finally, HOTAIR single nucleotide polymorphisms (SNPs) have been investigated as potential cancer susceptibility loci and linked to increased risk for human cancers, such as breast [17–19], esophageal squamous cell carcinoma [20], gastric [21–24], lung [25], and colorectal cancers [26]. However, the results remain controversial possibly due to the fact that independent studies are underpowered and biased, especially for small cohorts. Here, a meta-analysis of eligible studies conducted before January 20, 2016 was performed in order to obtain more precise and comprehensive insight into the impact of HOTAIR polymorphisms on cancer susceptibility. The results indicated that HOTAIR polymorphisms are associated with increased cancer risk, but mainly in stratified analysis based on ethnicity and cancer type.
RESULTS
Study characteristics
Our database search yielded 10 studies with a total of 7,772 cases and 9,075 controls that were eligible for our meta-analysis [17–26]. The main features of the eligible studies, which included genotyping method, are listed in Table 1. All were case-control studies and were comprised of individuals of Asian (n = 7) and Turkish descent (n = 3). In addition, the studies covered diverse tumor types: gastric cancer (n = 4) [21–24], breast cancer (n = 3) [17–19], colorectal cancer (n = 1) [26], lung cancer (n = 1) [25], and esophageal squamous cell carcinoma (n = 1) [20]. Quality of the included studies was assessed using the Newcastle Ottawa Scale, and all the studies scored a 7 or above (high-quality).
Table 1. Characteristics of studies on association between HOTAIR polymorphisms and cancers.
| Author | Year | Ethnicity | Cases | Controls | Source of Controls | Cancers | Single Nucleotide Polymorphisms | Genotyping Method | Quality Score |
|---|---|---|---|---|---|---|---|---|---|
| Gong | 2016 | Chinese | 498 | 213 | HB | Lung cancer | rs4759314, rs7958904, rs1899663 | MALDI-TOF mass spectrometry | 7 |
| Yan | 2015 | Chinese | 502 | 504 | PB | Breast cancer | rs1899663, rs4759314, rs920778 | PCR-RFLP, CRS-RFLP | 8 |
| Xue | 2015 | Chinese | 1734 | 1855 | HB | Colorectal cancer | rs4759314, rs7958904, rs874945 | TaqMan | 7 |
| Pan | 2015 | Chinese | 800 | 1600 | HB | Gastric cancer | rs920778, rs1899663, rs4759314 | PCR-RFLP | 7 |
| Guo | 2015 | Chinese | 515 | 654 | HB | Gastric cardia adenocarcinoma | rs12826786, rs4759314, rs10783618 | PCR-RFLP | 7 |
| Du | 2015 | Chinese | 1275 | 1646 | HB | Gastric cancer | rs4759314, rs7958904, rs874945 | TaqMan | 8 |
| Bayram | 2015 | Turkish | 123 | 122 | HB | Breast cancer | rs12826786 | TaqMan | 7 |
| Bayram | 2015 | Turkish | 104 | 209 | HB | Gastric cancer | rs920778 | TaqMan | 7 |
| Bayram | 2015 | Turkish | 123 | 122 | HB | Breast cancer | rs920778 | TaqMan | 7 |
| Zhang | 2014 | Chinese | 2098 | 2150 | HB | Esophageal squamous cell carcinoma | rs920778, rs1899663, rs4759314 | PCR-RFLP | 8 |
PB, population-based; HB, hospital-based; PCR-RFLP, polymerase chain reaction-restriction fragment length polymorphism; CRS-RFLP, created-restriction-site PCR-RFLP.
The number of HOTAIR SNPs extracted from all eligible studies was 7. Of these, only 2 SNPs, rs4759314 and rs920778, were reported in 5 or more studies and were thus the focus of the meta-analysis. The genotype distributions of HOTAIR rs4759314 and rs920778 SNPs are shown in Table 2.
Table 2. Genotype distributions of HOTAIR polymorphisms rs4759314 and rs920778.
| rs4759314 | AA genotype | AG genotype | GG genotype | P for HWE in controls | |||
|---|---|---|---|---|---|---|---|
| Study | Cases | Controls | Cases | Controls | Cases | Controls | |
| Yan (2015) | 451 | 448 | 50 | 54 | 1 | 2 | 0.78 |
| Xue (2015) | 1528 | 1608 | 200 | 236 | 5 | 11 | 0.47 |
| Pan (2015) | 451 | 914 | 48 | 83 | 1 | 3 | 0.45 |
| Guo (2015) | 461 | 589 | 53 | 64 | 1 | 1 | 0.59 |
| Du (2015) | 1083 | 1464 | 186 | 172 | 6 | 8 | 0.23 |
| Zhang (2014) | 917 | 910 | 81 | 89 | 2 | 1 | 0.44 |
| rs920778 | CC genotype | CT genotype | TT genotype | P for HWE in controls | |||
| Study | Cases | Controls | Cases | Controls | Cases | Controls | |
| Yan (2015) | 12 | 18 | 151 | 190 | 339 | 296 | 0.06 |
| Pan (2015) | 420 | 980 | 321 | 575 | 59 | 45 | < 0.01 |
| Bayram (2015-May) | 20 | 38 | 52 | 105 | 32 | 66 | 0.74 |
| Bayram (2015-Jan) | 31 | 15 | 52 | 66 | 40 | 41 | 0.14 |
| Zhang (2014) | 1091 | 1323 | 826 | 749 | 181 | 78 | 0.03 |
HWE, Hardy-Weinberg equilibrium.
HOTAIR polymorphism rs920778 is associated with a general susceptibility cancer
The first step in the analysis was therefore to determine whether either rs4759314 or rs920778 was associated with a general risk for cancer regardless of tissue origin. Analysis including all individuals from all eligible studies revealed that association between the polymorphism rs920778 and increased cancer risk was statistically significant in allele contrast (T vs C, OR = 1.239, 95% CI = 1.032 - 1.487, P = 0.021) and recessive genotype models (TT vs CT+CC, OR = 1.614, 95% CI = 1.082 - 2.406, P = 0.019). However, no significant association between the polymorphism rs4759314 and increased susceptibility to cancer in any genotype model was observed. The analysis thus revealed an association between the T allele or the TT genotype for polymorphism rs920778 and increased risk for cancer.
HOTAIR polymorphism rs4759314 is associated with an increased risk for gastric cancer
To determine whether HOTAIR polymorphisms rs4759314 or rs920778 were associated with risk for a specific cancer type, stratified analyses were performed on the basis of cancer tissue of origin. In this analysis, association of polymorphism rs4759314 with increased gastric cancer susceptibility was statistically significant under allele contrast (G vs A, OR = 1.262, 95% CI = 1.073 - 1.486, P = 0.005), dominant (GG+AG vs AA, OR = 1.280, 95% CI = 1.060 - 1.547, P = 0.010), and heterozygous (AG vs AA, OR = 1.288, 95% CI = 1.057 - 1.570, P = 0.012) models (Figure 1). These results indicated that the G allele of rs4759314 was linked to increased susceptibility specifically for gastric cancer.
Figure 1. Forest plot of cancer risk in different cancer types associated with HOTAIR polymorphism rs4759314.
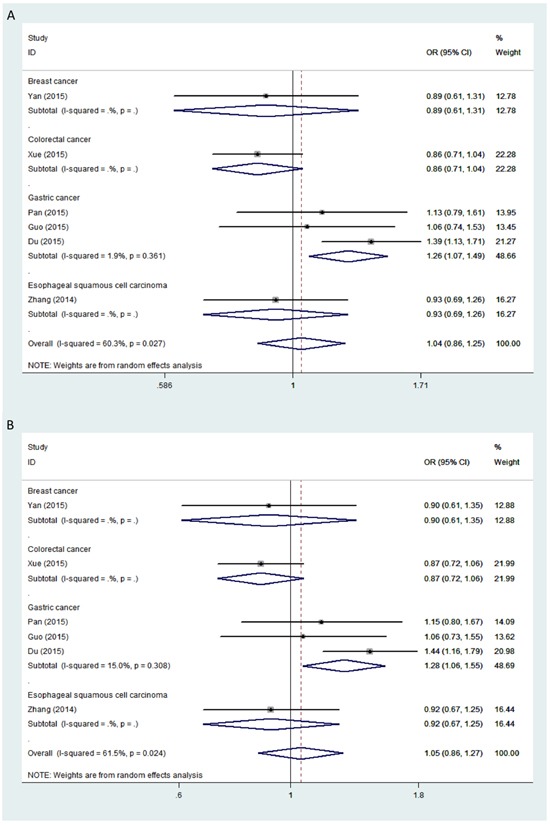
Models represented in A. allele contrast and B. dominant.
Significant between-study heterogeneity was observed in allele contrast (I-square = 60.3%, P = 0.027), dominant (I-square = 61.5%, P = 0.024), and heterozygous models (I-square = 60.4%, P = 0.027). Therefore, the random-effects model was used to pool the data. In the stratified analysis based on cancer tissue of origin, no significant heterogeneity was found in the gastric cancer group.
HOTAIR polymorphism rs920778 is associated with cancer susceptibility in Asians
The studies consisted of individuals of Asian and Turkish descent. Therefore associations between the polymorphisms and a general risk for cancer were examined on the basis of ethnicity. All individuals from studies involving rs4759314 were Asian so analysis stratified by ethnicity was performed only for rs920778. A significant association between rs920778 and cancer susceptibility was observed in Asians under all genetic models with the results as follows: allele contrast (T vs C, OR = 1.456, 95% CI = 1.349 - 1.571, P < 0.001); dominant model (TT+CT vs CC, OR = 1.462, 95% CI = 1.325 - 1.613, P < 0.001); recessive model (TT vs CT+CC, OR = 2.128, 95% CI = 1.417 - 3.197, P < 0.001); homozygous model (TT vs CC, OR = 2.764, 95% CI = 2.221 - 3.440, P < 0.001); and heterozygous model (CT vs CC, OR = 1.323, 95% CI = 1.194 - 1.466, P < 0.001) (Figure 2). The T allele or the TT genotype rs920778 thus emerged as a potential genetic marker for increased cancer susceptibility especially in Asians.
Figure 2. Forest plot of cancer risk in different ethnicities associated with HOTAIR polymorphism rs920778.
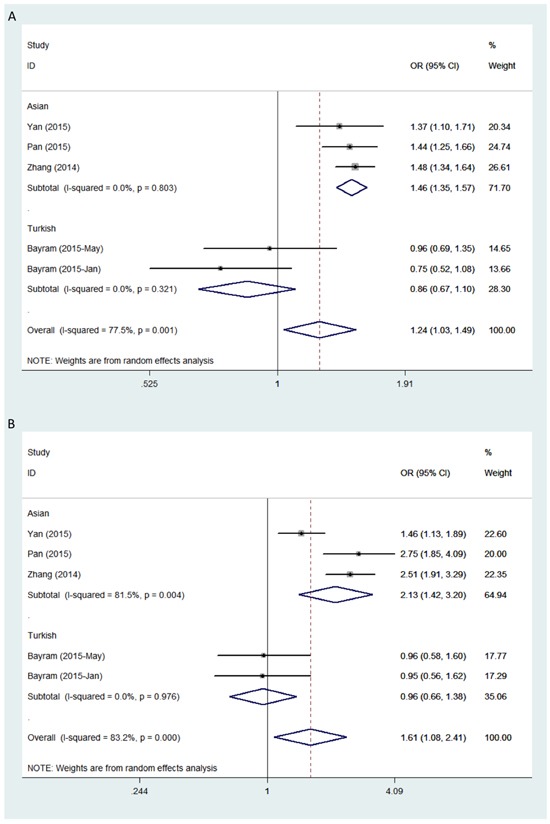
Models represented A. allele contrast and B. recessive.
Significant between-study heterogeneity was observed in all genetic models; therefore, analysis using the random-effects model was conducted on data stratified by ethnicity. Under these conditions, statistically significant heterogeneity was no longer observed within Asian or Turkish groups.
Sensitivity analysis
The stability of the results of the meta-analysis was assessed in sensitivity analysis, where the effect of each study on the pooled OR was examined by repeating the meta-analysis after sequentially omitting each study. Through the deletion of only a single study which was based on a Turkish population [18], an association of the polymorphism rs920778 with cancer risk emerged under dominant, homozygous, and heterozygous models. The P-values were not highly significant, however (data not shown). Therefore, the results of this meta-analysis including all eligible studies were generally and statistically stable.
Publication bias
Publication bias for each polymorphism in each model was evaluated using both the Begg's and Egger's tests. No obvious asymmetry was observed in any of the Begg's funnel plots indicating that publication bias was generally not a factor influencing the results (Figure 3). All P-values from the Egger's test and the Begg's test are listed in Table 3. The values are consistent with the absence of significant publication bias in the analysis of cancer risk and polymorphisms in most genotype models except in the case of polymorphism rs920778. For this polymorphism, publication bias was apparent in the allele contrast model. However, analysis with the trim and fill method demonstrated that the results of our study did not significantly change even after adjusting for the publication bias.
Figure 3. Publication bias tested by Begg's funnel plot.
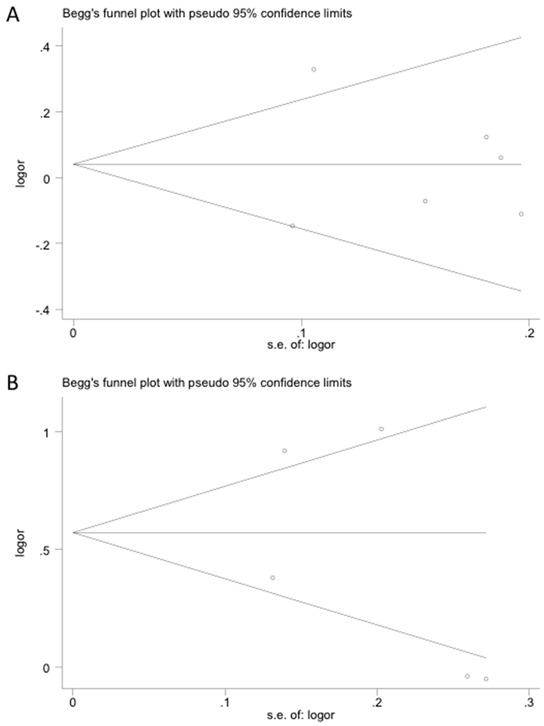
Polymorphisms represented in A. rs4759314 and B. rs920778.
Table 3. ORs and 95% CI for cancers and the HOTAIR polymorphisms under different genetic models.
| Genetic models | n | OR [95% CI] | P (OR) | Model (method) | I-square (%) | P (H) | P (Begg) | P (Egger) |
|---|---|---|---|---|---|---|---|---|
| rs4759314 | ||||||||
| Allele contrast (G vs A) | ||||||||
| All | 6 | 1.037 [0.863 - 1.246] | 0.700 | R (D-L) | 60.3 | 0.027 | 1.000 | 0.903 |
| Gastric cancer | 3 | 1.262 [1.073 - 1.486] | 0.005 | R (D-L) | 1.9 | 0.361 | - | - |
| Dominant model (GG+AG vs AA) | ||||||||
| All | 6 | 1.049 [0.864 - 1.275] | 0.629 | R (D-L) | 61.5 | 0.024 | 1.000 | 0.868 |
| Gastric cancer | 3 | 1.280 [1.060 - 1.547] | 0.010 | R (D-L) | 15.0 | 0.308 | - | - |
| Recessive model (GG vs AG+AA) | ||||||||
| All | 6 | 0.739 [0.395 - 1.381] | 0.343 | F (M-H) | 0.0 | 0.880 | 0.707 | 0.495 |
| Gastric cancer | 3 | 0.933 [0.378 - 2.300] | 0.880 | F (M-H) | 0.0 | 0.934 | - | - |
| Homozygous model (GG vs AA) | ||||||||
| All | 6 | 0.747 [0.400 - 1.396] | 0.361 | F (M-H) | 0.0 | 0.863 | 0.707 | 0.532 |
| Gastric cancer | 3 | 0.968 [0.393 - 2.387] | 0.944 | F (M-H) | 0.0 | 0.931 | - | - |
| Heterozygous model (AG vs AA) | ||||||||
| All | 6 | 1.059 [0.872 - 1.286] | 0.566 | R (D-L) | 60.4 | 0.027 | 1.000 | 0.833 |
| Gastric cancer | 3 | 1.288 [1.057 - 1.570] | 0.012 | R (D-L) | 19.1 | 0.291 | - | - |
| rs920778 | ||||||||
| Allele contrast (T vs C) | ||||||||
| All | 5 | 1.239 [1.032 - 1.487] | 0.021 | R (D-L) | 77.5 | 0.001 | 0.027 | 0.030 |
| Asian | 3 | 1.456 [1.349 - 1.571] | < 0.001 | R (D-L) | 0.0 | 0.803 | - | - |
| Dominant model (TT+CT vs CC) | ||||||||
| All | 5 | 1.199 [0.916 - 1.570] | 0.186 | R (D-L) | 73.2 | 0.005 | 0.221 | 0.170 |
| Asian | 3 | 1.462 [1.325 - 1.613] | < 0.001 | R (D-L) | 0.0 | 0.953 | - | - |
| Recessive model (TT vs CT+CC) | ||||||||
| All | 5 | 1.614 [1.082 - 2.406] | 0.019 | R (D-L) | 83.2 | <0.001 | 0.462 | 0.442 |
| Asian | 3 | 2.128 [1.417 - 3.197] | < 0.001 | R (D-L) | 81.5 | 0.004 | - | - |
| Homozygous model (TT vs CC) | ||||||||
| All | 5 | 1.549 [0.843 - 2.846] | 0.159 | R (D-L) | 85.8 | <0.001 | 0.086 | 0.078 |
| Asian | 3 | 2.764 [2.221 - 3.440] | < 0.001 | R (D-L) | 0.0 | 0.403 | - | - |
| Heterozygous model (CT vs CC) | ||||||||
| All | 5 | 1.115 [0.862 - 1.441] | 0.407 | R (D-L) | 67.8 | 0.014 | 0.221 | 0.143 |
| Asian | 3 | 1.323 [1.194 - 1.466] | < 0.001 | R (D-L) | 0.0 | 0.938 | - | - |
OR, Odds ratio; CI, confidence intervals; P (H), P for heterogeneity; n, number of included studies; F, fixed-effect model; R, random-effects model; M-H, Mantel-Haenszel method; D-L, DerSimonian-Laird method.
DISCUSSION
The overexpression of HOTAIR in various cancer types has led to its examination as a candidate molecule for the diagnosis and treatment of the disease. Here, HOTAIR was investigated for potential links to cancer susceptibility. We performed a meta-analysis using data from available cases (n = 7,772) and controls (n = 9,075) in the literature to date to provide further evidence that HOTAIR polymorphisms rs4759314 and rs920778 are associated with cancer risk. Only polymorphism rs920778 was found to be generally associated with increased risk for cancer, but statistically significant associations for both polymorphisms were revealed when data were stratified based on ethnicity and cancer type.
The results of our analysis indicated that the T allele or TT genotype of polymorphism is a potential genetic marker for cancer susceptibility, especially in Asians, and that the G allele in polymorphism rs4759314 might be related to susceptibility for gastric cancer. A possible mechanism underlying the association of these alleles to increased susceptibility is through the regulation of the expression of HOTAIR itself. Polymorphism rs4759314 resides in an intronic promoter region, which was found to influence the activity of this promoter and expression of HOXC11 gene [23]. For polymorphism rs4759314, increased HOTAIR expression originated from the G (relative to the A) allele in luciferase reporter assays. A similar finding was made for polymorphism rs920778, which resides in a novel HOTAIR intronic enhancer. In this case, increased HOTAIR expression was associated with T allele carriers [20, 22]. For both polymorphisms, the results of the functional assay correlated with the results of our meta-analysis; higher risk alleles drove increased expression of HOTAIR.
A sufficient number of cases and controls were pooled from different studies and provided a more accurate estimation of the associations between the HOTAIR polymorphisms and cancer risk as compared to individual studies. However, some limitations of this meta-analysis exist. First, although the analysis was performed with strict criteria for study inclusion and precise data extraction, significant between-study heterogeneity existed in some comparisons. However, after analysis stratified by cancer type or ethnicity, heterogeneity between the subgroups was significantly reduced. Second, our analysis was limited to individuals of Asian and Turkish descent so that it remains unclear as to whether these results can be generalized to other populations.
In conclusion, our results indicate that HOTAIR polymorphism rs920778 is more generally associated with cancer risk, particularly in Asians, whereas polymorphism rs4759314 may be a risk factor for gastric cancer. However, these polymorphisms require further evaluation in future well-designed studies as potential genetic susceptibility loci in different cancers as well as ethnic populations.
MATERIALS AND METHODS
Search strategy
PubMed, Embase, and Web of Science databases were searched for studies reporting association of HOTAIR polymorphisms with cancer risk up to the date of January 20, 2016. The literature search was performed using free-text words combined with Medical Subject Headings (MeSH), such as “Neoplasms”, “HOTAIR long untranslated RNA, human” and “Polymorphism, Single Nucleotide”. Gene-specific terms (HOX transcript antisense intergenic RNA or HOTAIR) were combined with polymorphism-specific terms (polymorphism or polymorphisms or variation or variations or variant or variants or mutation or mutations or genotype or genotypes) and disease-specific terms (cancer or cancers or tumor or tumors or neoplasm or neoplasms) to retrieve eligible studies. References cited in retrieved articles were also reviewed to identify additional potentially relevant studies.
Inclusion and exclusion criteria
Inclusion criteria for studies were the following: (1) case-control or cohort study design; (2) evaluating associations between HOTAIR polymorphisms and all types of cancer; (3) providing sufficient data for SNP allele and genotype frequencies; (4) published in English; and (5) performed on humans. Exclusion criteria were the following: (1) reviews and comments; (2) performed on animals, and (3) duplication of a previous publication.
Data extraction
Two investigators (QQ and JW) independently extracted the following data from each study: first author's surname, publication year, ethnicity, cancer types, numbers of cases and controls, and genotype distributions of cases and controls. Different ethnicity descents were categorized as Asian or Turkish. Study design was stratified into population-based and hospital-based studies. The results were compared and disagreement was resolved by discussion with a third reviewer (BH) until consensus was reached.
Quality assessment
The Newcastle-Ottawa Scale and Agency for Healthcare Research and Quality (http://www.ohri.ca/programs/clinical_epidemiology/oxford.asp; maximum score = 9 points) was used to evaluate the methodological quality, which scored studies based on the selection of patients, the comparability of the groups, and the quality of the sampling process. A study awarded a score of 0 - 3, 4 - 6, or 7 - 9 was considered as a low-, moderate-, or high-quality study, respectively.
Statistical analysis
To obtain a more comprehensive assessment of associations between HOTAIR polymorphisms and cancer susceptibility, five different comparison models were used: allele contrast, dominant, recessive, homozygous and heterozygous. Odds ratio (OR) with 95% confidence intervals (CI) was used to estimate the strength of associations, and the significance of ORs was determined with the Z test. Heterogeneity among the included studies was assessed by Chi square-based Q statistic. A random-effects (DerSimonian-Laird method) or fixed-effect (Mantel-Haenszel method) model was used to calculate pooled effect estimates in the presence (P < 0.05) or absence (P > 0.05) of heterogeneity. The method of subgroup analysis according to the ethnicity or cancer type of the participants was also applied to gain more precise results. Sensitivity analysis was conducted by sequentially excluding each study. The Begg's test and the Egger's test were performed to analyze for the presence of publication bias. If any possible bias was observed, the trim and fill method was used to identify and adjust for those studies. Data analyses were carried out using Stata software, version 11.0 (Stata Corporation; College Station, TX, USA). P-values < 0.05 were considered statistically significant.
Acknowledgments
This work was supported by Natural Science Foundation of China Grant (grants 81502164, 81402060 and 81572487), the Shandong Provincial Natural Science Foundation Grant (grants BS2015YY004 and BS2014YY033), the Special foundation for Taishan Scholars (grants ts20110814 and tshw201502056), the Fundamental Research Funds of Shandong University, the Department of Science & Technology of Shandong Province (grant 2015GGE27101 and 2015ZDXX0801A01), the University of Bergen, Helse Bergen, Norway and the Norwegian Centre for International Cooperation in Education (SIU)(UTF-2014/10047).
Footnotes
CONFLICTS OF INTEREST
The authors declare no conflicts of interest.
REFERENCES
- 1.Ponting CP, Oliver PL, Reik W. Evolution and functions of long noncoding RNAs. Cell. 2009;136:629–641. doi: 10.1016/j.cell.2009.02.006. [DOI] [PubMed] [Google Scholar]
- 2.Mercer TR, Dinger ME, Mattick JS. Long non-coding RNAs: insights into functions. Nat Rev Genet. 2009;10:155–159. doi: 10.1038/nrg2521. [DOI] [PubMed] [Google Scholar]
- 3.Qiu MT, Hu JW, Yin R, Xu L. Long noncoding RNA: an emerging paradigm of cancer research. Tumour Biol. 2013;34:613–620. doi: 10.1007/s13277-013-0658-6. [DOI] [PubMed] [Google Scholar]
- 4.Yang G, Lu X, Yuan L. LncRNA: a link between RNA and cancer. Biochim Biophys Acta. 2014;1839:1097–1109. doi: 10.1016/j.bbagrm.2014.08.012. [DOI] [PubMed] [Google Scholar]
- 5.Gibb EA, Brown CJ, Lam WL. The functional role of long non-coding RNA in human carcinomas. Mol Cancer. 2011;10:38. doi: 10.1186/1476-4598-10-38. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 6.Gutschner T, Diederichs S. The hallmarks of cancer: a long non-coding RNA point of view. RNA Biol. 2012;9:703–719. doi: 10.4161/rna.20481. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 7.Bhan A, Mandal SS. LncRNA HOTAIR: A master regulator of chromatin dynamics and cancer. Biochim Biophys Acta. 2015;1856:151–164. doi: 10.1016/j.bbcan.2015.07.001. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 8.Wu Y, Zhang L, Wang Y, Li H, Ren X, Wei F, Yu W, Wang X, Zhang L, Yu J, Hao X. Long noncoding RNA HOTAIR involvement in cancer. Tumour Biol. 2014;35:9531–9538. doi: 10.1007/s13277-014-2523-7. [DOI] [PubMed] [Google Scholar]
- 9.Woo CJ, Kingston RE. HOTAIR lifts noncoding RNAs to new levels. Cell. 2007;129:1257–1259. doi: 10.1016/j.cell.2007.06.014. [DOI] [PubMed] [Google Scholar]
- 10.Zhang S, Chen S, Yang G, Gu F, Li M, Zhong B, Hu J, Hoffman A, Chen M. Long noncoding RNA HOTAIR as an independent prognostic marker in cancer: a meta-analysis. PLoS One. 2014;9:e105538. doi: 10.1371/journal.pone.0105538. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 11.Tsai MC, Manor O, Wan Y, Mosammaparast N, Wang JK, Lan F, Shi Y, Segal E, Chang HY. Long noncoding RNA as modular scaffold of histone modification complexes. Science. 2010;329:689–693. doi: 10.1126/science.1192002. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 12.Endo H, Shiroki T, Nakagawa T, Yokoyama M, Tamai K, Yamanami H, Fujiya T, Sato I, Yamaguchi K, Tanaka N, Iijima K, Shimosegawa T, Sugamura K, Satoh K. Enhanced expression of long non-coding RNA HOTAIR is associated with the development of gastric cancer. PLoS One. 2013;8:e77070. doi: 10.1371/journal.pone.0077070. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 13.Kim K, Jutooru I, Chadalapaka G, Johnson G, Frank J, Burghardt R, Kim S, Safe S. HOTAIR is a negative prognostic factor and exhibits pro-oncogenic activity in pancreatic cancer. Oncogene. 2013;32:1616–1625. doi: 10.1038/onc.2012.193. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 14.Liu XH, Liu ZL, Sun M, Liu J, Wang ZX, De W. The long non-coding RNA HOTAIR indicates a poor prognosis and promotes metastasis in non-small cell lung cancer. BMC Cancer. 2013;13:464. doi: 10.1186/1471-2407-13-464. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 15.Su X, Malouf GG, Chen Y, Zhang J, Yao H, Valero V, Weinstein JN, Spano JP, Meric-Bernstam F, Khayat D, Esteva FJ. Comprehensive analysis of long non-coding RNAs in human breast cancer clinical subtypes. Oncotarget. 2014;5:9864–9876. doi: 10.18632/oncotarget.2454. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 16.Zhang JX, Han L, Bao ZS, Wang YY, Chen LY, Yan W, Yu SZ, Pu PY, Liu N, You YP, Jiang T, Kang CS. HOTAIR, a cell cycle-associated long noncoding RNA and a strong predictor of survival, is preferentially expressed in classical and mesenchymal glioma. Neuro Oncol. 2013;15:1595–1603. doi: 10.1093/neuonc/not131. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 17.Yan R, Cao J, Song C, Chen Y, Wu Z, Wang K, Dai L. Polymorphisms in lncRNA HOTAIR and susceptibility to breast cancer in a Chinese population. Cancer Epidemiol. 2015;39:978–985. doi: 10.1016/j.canep.2015.10.025. [DOI] [PubMed] [Google Scholar]
- 18.Bayram S, Sumbul AT, Batmaci CY, Genc A. Effect of HOTAIR rs920778 polymorphism on breast cancer susceptibility and clinicopathologic features in a Turkish population. Tumour Biol. 2015;36:3863–3870. doi: 10.1007/s13277-014-3028-0. [DOI] [PubMed] [Google Scholar]
- 19.Bayram S, Sumbul AT, Dadas E. A functional HOTAIR rs12826786 C>T polymorphism is associated with breast cancer susceptibility and poor clinicopathological characteristics in a Turkish population: a hospital-based case-control study. Tumour Biol. 2016;37:5577–5584. doi: 10.1007/s13277-015-4430-y. [DOI] [PubMed] [Google Scholar]
- 20.Zhang X, Zhou L, Fu G, Sun F, Shi J, Wei J, Lu C, Zhou C, Yuan Q, Yang M. The identification of an ESCC susceptibility SNP rs920778 that regulates the expression of lncRNA HOTAIR via a novel intronic enhancer. Carcinogenesis. 2014;35:2062–2067. doi: 10.1093/carcin/bgu103. [DOI] [PubMed] [Google Scholar]
- 21.Bayram S, Ulger Y, Sumbul AT, Kaya BY, Rencuzogullari A, Genc A, Sevgiler Y, Bozkurt O, Rencuzogullari E. A functional HOTAIR rs920778 polymorphism does not contributes to gastric cancer in a Turkish population: a case-control study. Fam Cancer. 2015;14:561–567. doi: 10.1007/s10689-015-9813-0. [DOI] [PubMed] [Google Scholar]
- 22.Pan W, Liu L, Wei J, Ge Y, Zhang J, Chen H, Zhou L, Yuan Q, Zhou C, Yang M. A functional lncRNA HOTAIR genetic variant contributes to gastric cancer susceptibility. Mol Carcinog. 2015 doi: 10.1002/mc.22261. [DOI] [PubMed] [Google Scholar]
- 23.Du M, Wang W, Jin H, Wang Q, Ge Y, Lu J, Ma G, Chu H, Tong N, Zhu H, Wang M, Qiang F, Zhang Z. The association analysis of lncRNA HOTAIR genetic variants and gastric cancer risk in a Chinese population. Oncotarget. 2015;6:31255–31262. doi: 10.18632/oncotarget.5158. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 24.Guo W, Dong Z, Bai Y, Guo Y, Shen S, Kuang G, Xu J. Associations between polymorphisms of HOTAIR and risk of gastric cardia adenocarcinoma in a population of north China. Tumour Biol. 2015;36:2845–2854. doi: 10.1007/s13277-014-2912-y. [DOI] [PubMed] [Google Scholar]
- 25.Gong WJ, Yin JY, Li XP, Fang C, Xiao D, Zhang W, Zhou HH, Li X, Liu ZQ. Association of well-characterized lung cancer lncRNA polymorphisms with lung cancer susceptibility and platinum-based chemotherapy response. Tumour Biol. 2016 doi: 10.1007/s13277-015-4497-5. [DOI] [PubMed] [Google Scholar]
- 26.Xue Y, Gu D, Ma G, Zhu L, Hua Q, Chu H, Tong N, Chen J, Zhang Z, Wang M. Genetic variants in lncRNA HOTAIR are associated with risk of colorectal cancer. Mutagenesis. 2015;30:303–310. doi: 10.1093/mutage/geu076. [DOI] [PubMed] [Google Scholar]