

Abstract
Peripheral CD4+CXCR5+PD-1+ T cells are a putative circulating counterpart to germinal center T follicular helper (TFH) cells. They show both phenotypic and functional similarities to TFH cells, which provide necessary help for the differentiation of B cells to antibody-secreting plasmablasts. In this study we evaluated the frequency, phenotypes, and responses of peripheral TFH-like (pTFH) cells to superantigen and recall antigen stimulation in 10 healthy and 34 chronically infected treatment-naïve HIV-1+ individuals. There was no difference in the frequency of pTFH cells between HIV+ and HIV− individuals. Surface expression of ICOS, but not CD40L, was higher on pTFH cells at baseline in HIV+ individuals. Compared to HIV− individuals, pTFH cells from HIV+ individuals had decreased maximal expression of ICOS and CD40L in response to in vitro superantigen stimulation. This decreased response did not correlate with viral control, CD4+ T cell count, duration of infection, or the degree of neutralizing antibody breadth. Despite a decreased maximal response, however, pTFH responses to HIV gag and tetanus toxoid recall antigens were preserved.
Keywords: CD4+ T cells, pTFH cells, HIV-specific CD4+ T cells, broadly neutralizing antibodies, chronic HIV infection
Introduction
T follicular helper (TFH) cells are a subset of CD4+ T cells that help B cells differentiate into antibody-secreting plasmablasts.1 Along with PD-1, CXCR5 is the canonical TFH marker, binding to CXCL13 to properly migrate within the germinal centers (GC). There is also a subset of CD4+CXCR5+PD-1+ T cells in the periphery, which are superior to other CD4+ subsets in their ability to help B cells.2–4 Based on functional properties, peripheral CD4+CXCR5+PD-1+ T cells are a putative circulating TFH population, although their precise relation to GC TFH cells is still under investigation.
Defining TFH-like cells in the periphery has been challenging since the transcription factor that promotes TFH differentiation, Bcl-6, is not detectable in protein form in cells outside the germinal center.5–8 Early studies relied only on CXCR5 expression on CD4+ T cells to identify ‘pTFH’ cells.3,9 However, subsequent studies determined that both CXCR5 and PD-1 are required to identify functional pTFH memory cells.5,6,10 Human GC TFH cells also express ICOS, CD40L, and SAP, and secrete IL-21. These proteins are necessary for T cell – B cell co-localization, adhesion, and signaling.1 Specifically, ICOS is a co-stimulatory molecule that engages ICOS ligand on B cells, resulting in T cell proliferation and production of cytokines that further support B cell differentiation.11 CD40L must also be expressed on the surface of TFH cells, as its interaction with CD40 on B cell surfaces is required for germinal center formation, and directly promotes B cell proliferation and isotype switching.12 Circulating CD4+CXCR5+PD-1+ cells express high levels of CD40L and ICOS after stimulation and thus combinations of these markers are routinely used to identify TFH-like cells in the periphery.5,6,8,10,13–15
In HIV infection, CD4+ T cell dysfunction occurs early and precedes the absolute loss of CD4+ T cells.16–18 T helper cells from HIV-infected individuals express high levels of inhibitory receptors, resulting in a diminished ability to help B cells.10,18–25 The role of TFH cell dysfunction in the dysregulation of B cells is less clear, but GC TFH cell populations have been shown to be expanded in both HIV and SIV infection.26,27 This is consistent with B cell dysregulation observed in HIV-1 infection, which is characterized by hypergammaglobulinemia, altered maturation patterns, and exhausted phenotypes.28
In this study we evaluated the frequency, phenotype, and responsiveness of peripheral TFH cells (‘pTFH cells’, defined as CD4+CXCR5+PD-1+ T cells) in HIV-1− and chronically infected treatment-naïve HIV-1+ individuals. In a series of in vitro stimulation assays we observed that pTFH cells from HIV-infected individuals had decreased maximal responses to superantigen stimulation as measured by their ability to express ICOS and CD40L. These decreased maximal responses in HIV+ subjects did not correlate with clinical aspects of disease or neutralizing antibody responses. We also show for the first time that HIV-specific and tetanus-specific responses are maintained within the pTFH cell population in HIV-infected individuals.
Methods
Human subjects
Peripheral blood mononuclear cells (PBMCs) from 10 HIV− and 34 HIV+ individuals were separated from blood samples using a Ficoll-Paque™ Plus density gradient. PBMCs were cryopreserved and stored in liquid nitrogen in media composed of 90% fetal bovine serum containing 10% DMSO. All HIV+ individuals were treatment-naïve and CD4+ T cell counts and viral loads were obtained at the time of donation (Table S1). The Vanderbilt University School of Medicine’s Institutional Review Board approved this study, and all individuals provided written informed consent.
In vitro stimulation assays
Cyropreserved PBMCs were thawed and washed twice in PBS and either stained immediately or cultured for stimulation assays. PBMCs were cultured at 10 million cells/mL in R10 media (RPMI 1640 containing 10% heat inactivated FCS, 2 mM L-glutamine, 50 ug/mL penicillin, 50 ug/mL streptomycin, and 10mM HEPES buffer (Gibco, Life Technologies)) and co-stimulated with anti-CD28 and anti-CD49d (1uL/mL each, from BD). Stimulation conditions included Staphylococcal Enterotoxin B (SEB) (1ug/mL, Sigma), HIV-1 PTE Gag peptides (1ug/mL, NIH AIDS Reagent Program),29,30 tetanus toxoid (10ug/mL, Astarte Biologics), and AT-2 inactivated HIV-1 MN particles (0.53ug/mL p24, generously provided by Dr. Jeff Lifson).23,31,32. For comparison to SEB and tetanus stimulation, PBMCs were incubated in R10 media alone. As a control for HIV-1 PTE Gag peptide stimulation (suspended in 0.8% DMSO), cells were suspended in R10 media containing 0.8% DMSO. For comparison to HIV-1 MN, PBMCs were incubated with MN control particles containing AT-2 treated microvesicles prepared from matched uninfected cultures, used at a comparable total protein concentration.23,31,32 In all stimulation assays, cells were incubated overnight at 37°C with 5% CO2. After 16 hours cells were removed from the plate, washed twice with PBS, and stained as described below.
Multicolor flow cytometry
Surface markers were evaluated using combinations of fluorochrome-conjugated monoclonal antibodies that were each titrated individually for their optimal stain index. PBMCs were stained at 10 million cells/mL in 200uL PBS. All PBMCs were incubated for 10 minutes with an amine-reactive viability dye (LIVE/DEAD Aqua, Invitrogen), washed twice, and then stained for 15 minutes at room temperature with combinations of monoclonal antibodies. For ex vivo phenotyping, cells were stained with CD3-AF700 (UCHT1, BD), CD4-PECy5 (RPA-T4, BD), CD8-APC-AF750 (3B5, Invitrogen), CD45RO-PETR (UCHL1, Beckman Coulter), CCR7-BV421 (150503, BD), CXCR5-AF488 (RF8B2, BD), PD-1-PE (EH12.2H7, BioLegend), CD14-V500 (M5E2, BD), and CD19-V500 (HIB19, BD). In vitro phenotyping was performed with combinations of CD3-AF700, CD4-PECy5, CD8-APC-AF750, CD45RO-PETR, CXCR5-AF488, CD14-V500, CD19-V500, ICOS-PE (DX29, BD), CD40L-PE (TRAP1, BD) and PD-1-BV421 (EH12.2H7, BioLegend). All PBMCs were washed twice after staining, fixed with 2% paraformaldehyde, and analyzed on a BD LSR Fortessa (BD Biosciences) at the VMC Flow Cytometry Shared Resource.
Flow cytometry data was analyzed using BD Biosciences FACSDiva Software. In all experiments, forward and side scatter were used to identify lymphocytes and from that population non-viable, CD14+, CD19+, CD8+ cells were excluded from further analysis (Fig. S1).
Antibody neutralization assays
Neutralization assays were performed using envelopes from clades A, B, and C in the TZM-bl cell based pseudovirus assay, as previously described.33 The clade B and C env clones were chosen from standard env panels,34,35 and the clade A env clones were isolated from Kenyan sex workers.36 The env clones chosen for this study represent a range of neutralization sensitivities of transmitted HIV-1 viruses. Plasma samples were titrated 2 fold from 1:20 to 1:2560 and were incubated for 90 minutes at 37°C in the presence of single-round-competent virions (pseudovirus). The neutralization values reported here are the IC50. Only 30 individuals were evaluated in the antibody neutralization assays due to sample availability.
Statistical Analysis
Analysis was performed using GraphPad Prism Software (GraphPad, La Jolla, CA, USA). Paired comparisons (within a single subject) were analyzed with the Wilcoxon matched paired t test. Comparisons between healthy controls and HIV+ subjects were analyzed with the Mann-Whitney t tests. Correlation data was evaluated for statistical dependence using Spearman’s rank correlation coefficient rho (ρ). All tests were 2-tailed and were considered statistically significant at p<0.05.
Results
Frequency of CD4+CXCR5+PD-1+ T cells in the blood of HIV+ and HIV− individuals
T follicular helper (TFH) cells are more frequent in the lymph nodes during HIV and SIV infection compared to lymph nodes from uninfected controls20,26,27,37; however, there are conflicting reports comparing the frequency of circulating TFH-like cells among HIV-infected and HIV-uninfected individuals.6,9,10 We first assessed the expression of the canonical TFH markers CXCR5 and PD-1 on circulating CD4+ T cells (Fig. 1A and Fig. S1A). The frequency of CD4+CXCR5+PD-1+ T cells was measured in a cohort of 34 treatment-naïve HIV-infected individuals with CD4+ T cell counts > 300 cells/mm3 and a wide range of viral loads (Table S1). There was no difference in the frequency of CD4+ cells with dual CXCR5 and PD-1 expression (pTFH cells) between HIV− and HIV+ individuals, which constituted an average of 2% of CD4+ T cells (Fig. 1B).
Figure 1. Ex vivo identification of peripheral CD4+ T cells expressing CXCR5 and PD-1 in HIV+ and HIV-individuals.
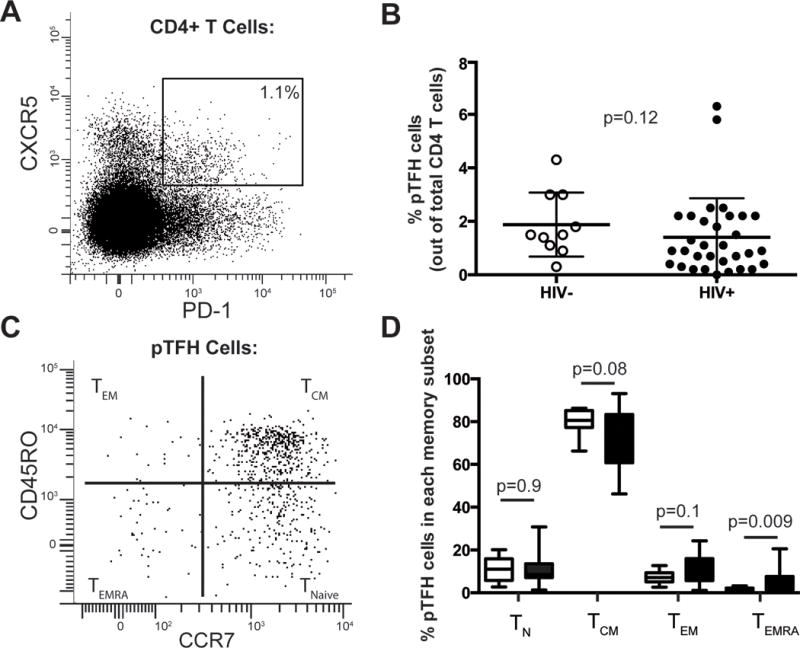
(A) Representative plot of CXCR5 and PD-1 expression on live CD19−CD14−CD8−CD3+CD4+ cells. Gate indicates frequency of pTFH cells out of total CD4+ T cells. (B) Frequency of pTFH cells (CXCR5+PD-1+) out of total CD4+ T cells in HIV− (open circles) and HIV+ individuals (closed circles). (C) Representative plot of CD45RO and CCR7 expression on pTFH cells. A quadrant gate was used to distinguish the four memory subsets. (D) Distribution of pTFH cells among each memory compartment in HIV− (open bars) and HIV+ (closed bars) individuals. p values are from Mann-Whitney t tests.
The distribution of pTFH cells within memory T cell subsets was then evaluated. We distinguished CD4+ memory subsets based on CCR7 and CD45RO expression: T naive (CCR7+CD45RO−), T central memory (TCM) (CCR7+CD45RO+), T effector memory (CCR7−CD45RO+), and T effector memory cells expressing CD45RA (CCR7−CD45RO−) (Fig. 1C and Fig. S1A).38,39 There were no significant differences between the distribution of CD4+ T cell memory populations between HIV+ and HIV− individuals in our cohort (Fig. S1B), and most pTFH cells had a TCM phenotype (Fig. 1D). In the absence of stimulation, few pTFH cells expressed CD40L (<5%) or ICOS (<10%) (Fig. S2A–B). After stimulation, however, significantly more pTFH cells were ICOS+ (mean 47%) and CD40L+ (44%) compared to other non-pTFH CD4+ populations (CXCR5+PD-1− and CXCR5− cells) (p<0.0001, Fig. S2C–D).
pTFH cells from HIV+ individuals have decreased maximal responses to in vitro stimulation compared to healthy donors
We next compared the ability of pTFH cells from HIV− and HIV+ individuals to express ICOS and CD40L in response to SEB stimulation (Fig. 2). The frequency of ICOS expression without stimulation was higher on pTFH cells from HIV+ individuals compared to HIV− individuals (median 6.5% and 2.2% respectively, p=0.0005; Fig. 2A–B). After stimulation with SEB, the frequency of ICOS-expressing pTFH cells increased dramatically but was lower in HIV+ compared to HIV− subjects (median 51.1% and 63.8% respectively, p=0.03; Fig. 2A–B). Accordingly, the change in the frequency of ICOS expression on pTFH cells in response to potent stimulation was decreased in HIV+ individuals compared to HIV− individuals (p=0.003; Fig. 2C).
Figure 2. pTFH cells from HIV+ individuals have decreased responsiveness to superantigen stimulation as measured by CD40L and ICOS expression.
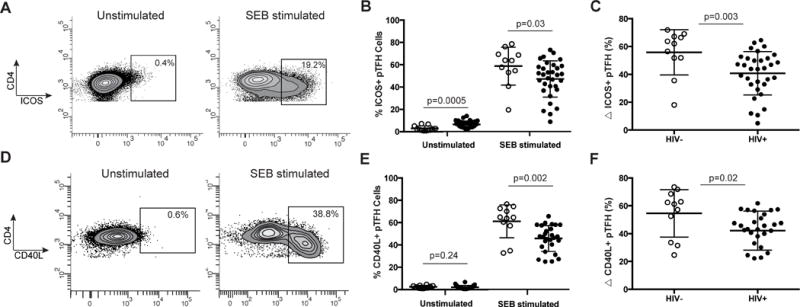
(A) Representative plots of ICOS expression on CD4+ T cells with and without SEB stimulation. (B) Frequency of ICOS on pTFH cells with and without SEB stimulation in healthy controls (open circles) and HIV+ individuals (closed circles). (C) Change in frequency of ICOS expression on pTFH cells in response to SEB stimulation was measured by subtracting the unstimulated from the stimulated conditions. (D) Representative plots of CD40L expression on CD4+ T cells. (E) Frequency of CD40L on pTFH cells with and without SEB stimulation in healthy controls (open circles) and HIV+ individuals (closed circles). (F) Change in frequency of CD40L expression on pTFH cells in response to SEB stimulation. All p values are derived from Mann-Whitney t tests.
The frequency of CD40L expression on pTFH cells was similar in HIV− and HIV+ subjects without stimulation (median 2.5% and 1.4% respectively, p=0.24; Fig. 2D–E). After SEB stimulation, however, the frequency of CD40L expression on pTFH cells isolated from healthy individuals was higher (64.6%) compared to HIV+ individuals (47.4%; p=0.002; Fig. 2D–E). Similarly to ICOS, the change in the frequency of CD40L+ expressing pTFH cells in HIV− individuals was higher than that of HIV+ individuals after SEB stimulation (p=0.02; Fig. 2F).
Decreased maximal pTFH responses did not correlate with clinical aspects of disease
We evaluated whether expression of TFH surface markers or responsiveness to in vitro stimulation was associated with either clinical aspects of the disease or the T cell phenotypes of our cohort. We found no correlation between the degree of response to SEB stimulation and viral load, CD4+ T cell count, or duration of infection (Fig. 3). Although PD-1 is a marker that identifies TFH cells, it is also a marker of immune exhaustion. We therefore evaluated whether the degree of PD-1 expression on CXCR5+ T cells predicted the ability of these cells to respond to in vitro stimulation. No relationship existed between the MFI of PD-1 on CXCR5+ cells in the HIV+ individuals and the ability of these cells to express ICOS or CD40L responses after SEB stimulation.
Figure 3. Decreased maximal pTFH responses did not correlate with clinical aspects of HIV infection or neutralizing antibody breadth.
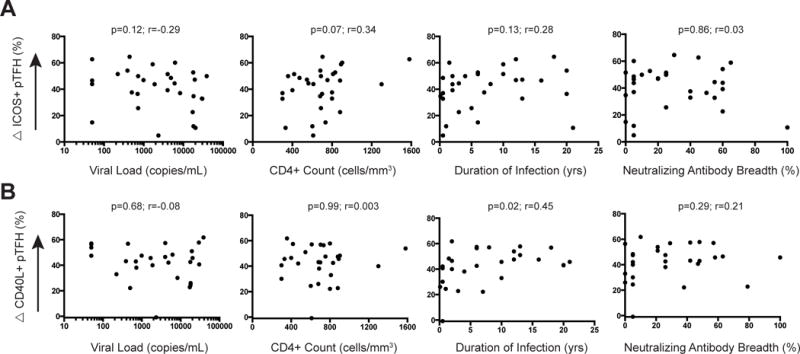
Change in frequency of ICOS (A) or CD40L (B) expression on pTFH cells in response to SEB stimulation did not correlate with viral load, CD4+ T cell count, duration of infection, or neutralizing antibody breadth in HIV+ individuals. There was a weak correlation between change in CD40L+ pTFH cells and duration of infection. p and r values were derived from Spearman rank tests.
No correlation between pTFH responses and antibody neutralization breadth
pTFH cells in the blood are capable of stimulating B cells to differentiate to immunoglobulin-secreting plasmablasts.3,4,6,10 Despite B cell dysregulation in HIV-1 infection, 20–30% of HIV+ subjects produce antibodies able to neutralize heterologous HIV-1 strains across clades A, B and C.33,40 To assess whether the frequency of pTFH cells or responsiveness to antigenic stimulation correlated with the production of broadly neutralizing antibodies, we analyzed the serum from all HIV+ subjects for the ability to neutralize heterologous HIV-1 envelopes (Fig. 4). The subjects in our cohort ranged widely (0–100%) in their ability to neutralize 20 different HIV-1 isolates from clades A, B, and C (Fig. 4). We found no correlation between the degree of neutralizing antibody breadth and the frequency of pTFH cells (Fig. S4). Additionally we found no correlation between the degree of neutralizing antibody breadth and the frequency or ability of pTFH cells to express ICOS or CD40L after in vitro stimulation with SEB (Fig. 3).
Figure 4. Plasma neutralization breadth of HIV+ individuals.
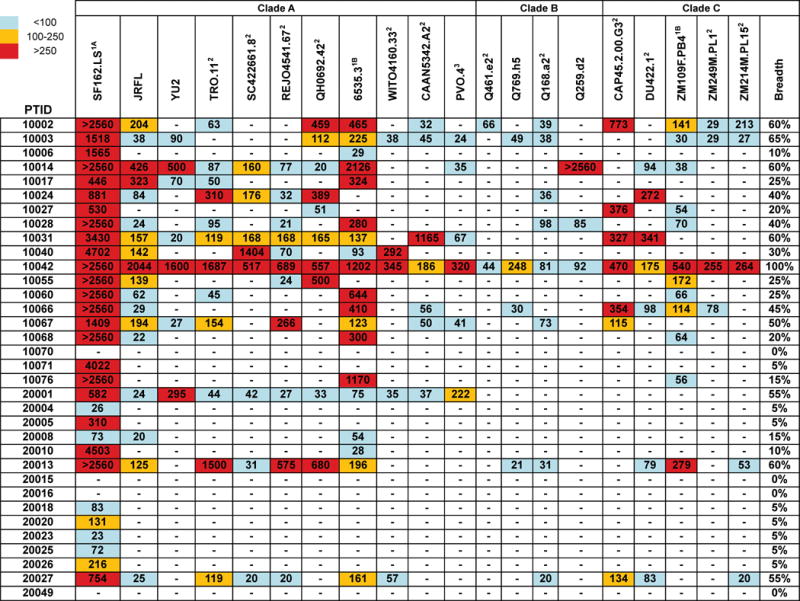
The cross-neutralizing activities of plasmas from HIV+ individuals were evaluated against the indicated clade A, B and C viruses. The values are the plasma titers at which 50% neutralization (IC50) was recorded. IC50 values are color-coded: (blue) IC50<1:100; (yellow) 1:100≤IC50≥1:250; (red) IC50>1:250. A (−) symbol indicates less than 50% neutralization was recorded. The term ‘breadth’ represents the percent of isolates neutralized by a plasma sample, out of the total number of isolates tested, irrespective of the potency of neutralization. With the exception of SF162.LS, which is a tier 1 virus, all viruses were tier 2.
pTFH cells from HIV+ individuals maintain recall antigen-specific responses
We next investigated recall antigen-specific responses of pTFH and non-pTFH (defined as CD3+CD4+CXCR5−) T cells. Since the majority of naïve T cells reside in the non-pTFH population, we only assessed memory (CD45RO+) CD4+ T cells in this assay. We performed stimulations with tetanus toxoid, HIV-1 Gag peptides, and inactivated HIV-1 MN to measure antigen-specific responses in HIV+ individuals (Fig. 5, Fig. S3, Fig. S5, Fig. S6). While the overall responses to tetanus toxoid were low, a higher fraction of pTFH cells from HIV+ individuals increased ICOS and CD40L expression compared to non-pTFH cells (p=0.02; Fig. 5A–B). There were no differences in ICOS or CD40L expression on pTFH and non-pTFH cell after tetanus toxoid stimulation in HIV− individuals (Fig. S5A–B). Furthermore, the responses of pTFH cells to tetanus were not different between HIV− and HIV+ individuals though the responses appear to be more robust in HIV+ individuals (Fig. S5C–D).
Figure 5. Memory pTFH cells preferentially express ICOS and CD40L after recall antigen stimulation compared to memory non-pTFH cells.
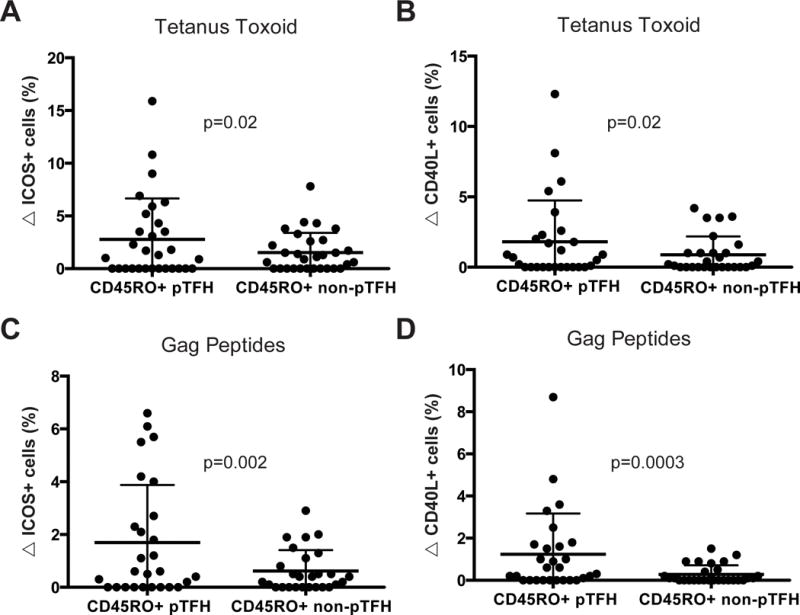
Cells analyzed were either memory pTFH cells (defined as CD45RO+CXCR5+PD-1+ CD4+ T cells) or memory non-pTFH cells (defined as CD45RO+CXCR5-CD4+ T cells). (A–B) Change in the frequency of ICOS+ (A) or CD40L+ (B) memory pTFH or non-pTFH cells in HIV+ individuals in response to 16 hours of stimulation with tetanus toxoid. (C–D) Change in the frequency of ICOS+ (C) or CD40L+ (D) memory pTFH or non-pTFH cells in HIV+ individuals in response to 16 hours of stimulation with Gag peptides. All p values are derived from Wilcoxon matched-pairs t-tests.
Responses to Gag peptides as measured by the change in ICOS and CD40L on CD4+ T cells was greater on pTFH compared to non-pTFH cells in HIV+ individuals (p=0.002 and p=0.0003; Fig. 5C–D). There were no significant changes in ICOS or CD40L expression in HIV− individuals stimulated with Gag peptides (p=0.34, data not shown). Since the majority of CD4+ T cells are CXCR5− a higher absolute number of CXCR5− cells increased ICOS expression in response to antigen stimulation; however, a higher fraction of CXCR5+PD-1+ cells were antigen-specific. These data demonstrate that recall antigen-specific memory pTFH cells are preserved in HIV+ individuals.
We also evaluated responses to AT-2 inactivated HIV-1 MN particles, which contain conformationally intact envelope proteins on their surface. Compared to control particles, MN did not induce changes in ICOS on pTFH in HIV− subjects (p=0.13). In HIV+ individuals, MN induced changes in ICOS on pTFH cells compared to control particles (p<0.0001) and responses were greater in memory pTFH cells compared to non-pTFH cells (p=0.02; Fig. S6A). Changes in CD40L in response to MN, however, were rare (only 12 of 28 subjects in the assay) and weak and did not allow us to accurately compare non-pTFH and pTFH responses (Fig. S6B). No antigen-specific responses to any of the recall antigens correlated with neutralizing antibody breadth in HIV+ individuals (Fig. S4 and data not shown).
Since the core identity markers of pTFH cells, PD-1 and CXCR5, can also go up with activation, we assessed the frequency of pTFH cells before and after stimulation. The median increase in the frequency of pTFH cells in response to SEB stimulation was 1.95%. Stimulation of PBMC with recall antigens only slightly increased the frequency of pTFH cells (median increase of 0.2% with Gag stimulation; median increase of 0.5% with tetanus stimulation). To further investigate the possibility of CXCR5− cells contributing to our pTFH population after overnight stimulation, we FACS sorted CD4+ T cells into four populations based on CXCR5 and PD-1 expression in 4 healthy individuals. Cells that were sorted CXCR5− (regardless of PD-1 expression) became CXCR5+ after SEB stimulation at a frequency of <2.5% and the MFI of CXCR5 after SEB stimulation never changed (Fig. S7 and data not shown). Thus while we cannot exclude the possibility that a small number of cells we termed ‘pTFH’ in these assays were recently activated T cells, the large majority of this ‘pTFH’ population expressed CXCR5 and PD-1 prior to stimulation.
Discussion
Multiple studies have demonstrated pTFH cells are similar to GC TFH cells in their ability to provide B cell help.2–6,41 We performed an ex vivo phenotypic analysis of pTFH cells in healthy and chronically infected HIV+ individuals, and we assessed the in vitro responses of this cell population to superantigen and recall antigen stimulation. CXCR5+ and PD-1 are currently accepted canonical markers required to identify pTFH cells.5,6,8,10,13–15 While additional cell markers such as CXCR3,6 CCR6,10 and TIGIT+42 have been suggested to further discriminate pTFH cells, we focused on CD4+ T cells expressing the main pTFH identifying canonical markers. We evaluated the ability of these cells to respond to various stimuli, and identified tetanus-specific and HIV-specific pTFH populations in HIV-infected individuals. We focused on ICOS and CD40L expression of pTFH cells, since these surface receptors must engage ICOS ligand and CD40 on the surface of B cells to drive B cell differentiation to antibody-secreting plasmablasts.
We observed pTFH cells circulate at similar frequencies in healthy and HIV+ treatment-naïve individuals. The frequencies of circulating pTFH cells we measured are in agreement with those measured in a cohort with CD4+ T cell numbers similar to ours.9 In separate studies, however, decreased pTFH cell frequencies were reported in a cohort with a mean CD4+ T cell count of 320 cells/mm3,10 and increased frequencies were found in a cohort of individuals that were only infected for an average of three years (CD4+ T cell values were not reported).6 Thus, variability within these cohorts likely explains the variability in measured frequencies of pTFH cells. We also show that the distribution of pTFH cells within T cell memory compartments in HIV infection is not perturbed by HIV infection, and confirm that most pTFH cells reside in the T central memory compartment in both HIV− and HIV+ individuals.2,3,9
GC TFH cells and pTFH cells isolated from HIV and SIV infected individuals are impaired in their ability to provide help necessary for B cells to differentiate to plasmablasts.9,10,19,20,26,27 However, the underlying changes in expression of key surface receptors on pTFH cells required for proper function have not been described. Here we provide an analysis of the surface expression of ICOS and CD40L in response to stimuli in chronic HIV-1 infection.
We found that after in vitro superantigen stimulation, pTFH cells from HIV-infected individuals expressed less ICOS and CD40L than pTFH cells from HIV-uninfected individuals. This reduced maximal expression of ICOS and CD40L might explain the decreased function of these cells observed in HIV and SIV infection.9,10,19,20,26,27 We attempted to determine correlates of this decreased response. We hypothesized that the MFI of PD-1 expression on pTFH cells would correlate with decreased responses in HIV-infected individuals. While PD-1 is required for proper TFH cell function,43,44 it is also associated with functional exhaustion of T cells during chronic HIV-1 infection.18,45 Blocking PD-1 and PD-L1 has been shown to enhance the ability of T cells to activate B cells in culture,23 and improves the ability of pTFH cells to stimulate B cells to produce IgG in HIV+ but not HIV− individuals.19 We found no correlation between PD-1 expression and decreased pTFH cell responsiveness in HIV-infected individuals. We also did not find a correlation between the responsiveness of pTFH cells with viral load, CD4 + T cell count, years post-infection, or neutralizing antibody breadth.
Recall-antigen specific pTFH cells have been identified in a few prior studies: tetanus-specific pTFH cells were identified using tetramers in healthy individuals6 and in HIV-infected individuals, small populations of Gag-specific pTFH cells were identified by CD40L expression and IL-21 secretion.10 More recently, HIV-specific pTFH cells (identified by IL-21 secretion after in vitro stimulation, and subsequently found to express transcripts for CXCR5, CD40L, and ICOS) were shown to recognize both HIV-1 Gag and Env peptides by ELISpot.41 In agreement with these studies, we found the majority of recall memory responses resides within the CD4+CXCR5- population, which makes up the majority of CD4+ T cells. However, the proportion of pTFH cells that increase ICOS or CD40L expression is significantly higher than that of non-pTFH cells in response to HIV Gag peptides and tetanus toxoid in HIV+ individuals. This was also true for ICOS responses to HIV-1 MN in HIV+ individuals, but not CD40L which we were unable to assess accurately since responses were so rare. HIV− individuals did not respond to Gag or MN stimulation as expected, but did have weak responses to tetanus toxoid stimulation. There was no difference in the frequency of these weak tetanus responses in pTFH compared to non-pTFH cells in HIV− individuals. It is difficult to make accurate conclusions from these findings, however, since we have no tetanus vaccination records from our study populations. However, the HIV+ individuals are followed very closely and more likely to be up-to-date on all vaccinations. Our novel findings show that HIV-specific CD4+ T cells are maintained within the pTFH population during chronic infection.
HIV and SIV infection have been shown to drive the expansion of GC TFH cells,19,27 but the relationship between the frequencies of GC TFH and peripheral TFH cells is unclear. Recent gene expression and functional studies disagree on precisely which combination of surface receptors identifies TFH-like cells in the periphery; however, there appears to be consensus that CXCR5 and PD-1 are required for identifying pTFH cells.5,6,10 Our observation of preferential expression of CD40L and ICOS expression on CD4+CXCR5+PD-1+ T cells in response to recall antigens further supports the identification of pTFH cells as a circulating counterpart of GC TFH cells. It is important to note that superantigen and recall antigen stimulation increased the frequency of CXCR5+PD-1+ cells in vitro. This increased frequency could reflect proliferation of pTFH cells or a new population of recently activated T cells. Cell sorting experiments, however, demonstrated that few sorted CXCR5- cells became CXCR5+PD-1+ after short-term superantigen stimulation, thus the majority of this population was likely CD4+CXCR5+PD-1+ cells prior to stimulation.
Future studies should recapitulate pTFH phenotypic changes in response to antigen with IL-21 measurement to establish the trends are the same. We tested many IL-21 antibody clones in intracellular cytokine assays under many stimulation conditions (PMA/ionomycin, anti-CD3/anti-CD28, and SEB) at 4, 6, 24, 48, and 72 hours without obtaining reliable results. Unfortunately, current antibodies to IL-21 protein are poor, making intracellular cytokine assays unable to address this question accurately.46 The development of new commercially available IL-21 detection kits (of protein or transcript) will be valuable for future single-cell IL-21 research.46
The generation of HIV-neutralizing antibody responses through vaccination has remained an elusive goal; understanding how naturally infected individuals are able to generate broadly neutralizing antibodies will be critical to our understanding of vaccine-induced immune responses. Here we analyzed the neutralization breadth of plasma from 30 HIV+ individuals in our cohort. In agreement with prior studies we did not find a direct correlation between neutralization breadth and the frequency of pTFH cells during chronic HIV infection.6,8 We also found no correlation between neutralization breadth and the frequency of ICOS+ pTFH cells. This is in contrast to findings by Locci, et al., however, subjects in that study were grouped into ‘top’ and ‘low’ neutralizers and statistical correlations were not performed.6 Despite a lack of correlation during chronic infection, Cohen, et al., found correlations between the frequency of pTFH cells and broadly neutralizing antibody responses in early infection (0.1–1 year after infection).8 One interpretation of these data is that during early HIV infection pTFH responses may not yet be impaired and thus can facilitate the generation of broadly neutralizing antibodies, and these antibody responses can be maintained over the course of chronic infection. Future studies should investigate the responsiveness of pTFH cells to HIV antigens longitudinally from early through chronic HIV infection.
In this study we describe our efforts to determine the responsiveness of pTFH cells in chronic HIV-1 infection. Specifically, we investigated two surface receptors required by TFH cells to provide help to B cells, ICOS and CD40L. We demonstrate that pTFH cells expressed the highest levels of CD40L and ICOS in response to superantigen and recall antigen stimulation compared to CD4+CXCR5+PD-1− T cells and CD4+CXCR5− T cells. We found the pTFH cells of HIV-infected individuals had impaired ability to increase expression of CD40L and ICOS in response to superantigen stimulation. Despite impaired maximal responses, however, this cell subset maintained the ability to respond to recall antigens. These results suggest that the evaluation of immune responses of pathogen-specific circulating pTFH cells will be important for future studies of natural infection and immune responses to vaccines.
Supplementary Material
Acknowledgments
We would like to acknowledge the VMC Flow Cytometry Shared Resource for their support.
Funding Sources: K.J.N. was supported by NIH-T32A1089554. K.J.N., S.K. and N.S. were supported by NIH-P01A1078064. Flow Cytometry experiments were performed in the VMC Flow Cytometry Shared Resource. The VMC Flow Cytometry Shared Resource is supported by the Vanderbilt Ingram Cancer Center (P30CA68485) and the Vanderbilt Digestive Disease Research Center (DK058404). Pilot flow cytometry experiments were supported in part by CTSA award No. UL1TR000445 from the National Center for Advancing Translational Sciences and the contents of this paper are solely the responsibility of the authors and do not necessarily represent official views of the National Center for Advancing Translational Sciences or the National Institutes of Health. This research was also supported in part by the Vanderbilt-Meharry Center for AIDS Research (CFAR), NIH-P30AI54999. The funders had no role in study design, data collection and interpretation, or the decision to submit the work for publication.
Footnotes
Contributions: overall plan (KJN, SAK), experimental design (KJN, DKF, SAK), flow cytometry experiments (KJN, RMS), flow cytometry data analysis (KJN, DKF), HIV-1 neutralization assays to determine patient antibody breadth (DNS), integrated data analysis and writing (KJN, SAK).
Conflicts of Interest: All authors declare no competing financial interests.
References
- 1.Crotty S. Follicular helper CD4 T cells (TFH) Annu Rev Immunol. 2011;29:621–663. doi: 10.1146/annurev-immunol-031210-101400. [DOI] [PubMed] [Google Scholar]
- 2.Chevalier N, Jarrossay D, Ho E, et al. CXCR5 expressing human central memory CD4 T cells and their relevance for humoral immune responses. J Immunol. 2011;186(10):5556–5568. doi: 10.4049/jimmunol.1002828. [DOI] [PubMed] [Google Scholar]
- 3.Morita R, Schmitt N, Bentebibel SE, et al. Human blood CXCR5(+)CD4(+) T cells are counterparts of T follicular cells and contain specific subsets that differentially support antibody secretion. Immunity. 2011;34(1):108–121. doi: 10.1016/j.immuni.2010.12.012. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 4.Bentebibel SE, Schmitt N, Banchereau J, Ueno H. Human tonsil B-cell lymphoma 6 (BCL6)-expressing CD4+ T-cell subset specialized for B-cell help outside germinal centers. Proc Natl Acad Sci U S A. 2011;108(33):E488–497. doi: 10.1073/pnas.1100898108. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 5.He J, Tsai LM, Leong YA, et al. Circulating precursor CCR7(lo)PD-1(hi) CXCR5(+) CD4(+) T cells indicate Tfh cell activity and promote antibody responses upon antigen reexposure. Immunity. 2013;39(4):770–781. doi: 10.1016/j.immuni.2013.09.007. [DOI] [PubMed] [Google Scholar]
- 6.Locci M, Havenar-Daughton C, Landais E, et al. Human Circulating PD-1CXCR3CXCR5 Memory Tfh Cells Are Highly Functional and Correlate with Broadly Neutralizing HIV Antibody Responses. Immunity. 2013 doi: 10.1016/j.immuni.2013.08.031. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 7.Nurieva RI, Chung Y, Martinez GJ, et al. Bcl6 mediates the development of T follicular helper cells. Science. 2009;325(5943):1001–1005. doi: 10.1126/science.1176676. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 8.Cohen K, Altfeld M, Alter G, Stamatatos L. Early preservation of CXCR5+ PD-1+ helper T cells and B cell activation predict the breadth of neutralizing antibody responses in chronic HIV-1 infection. J Virol. 2014;88(22):13310–13321. doi: 10.1128/JVI.02186-14. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 9.Pallikkuth S, Parmigiani A, Silva SY, et al. Impaired peripheral blood T-follicular helper cell function in HIV-infected nonresponders to the 2009 H1N1/09 vaccine. Blood. 2012;120(5):985–993. doi: 10.1182/blood-2011-12-396648. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 10.Boswell KL, Paris R, Boritz E, et al. Loss of circulating CD4 T cells with B cell helper function during chronic HIV infection. PLoS Pathog. 2014;10(1):e1003853. doi: 10.1371/journal.ppat.1003853. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 11.Hutloff A, Dittrich AM, Beier KC, et al. ICOS is an inducible T-cell co-stimulator structurally and functionally related to CD28. Nature. 1999;397(6716):263–266. doi: 10.1038/16717. [DOI] [PubMed] [Google Scholar]
- 12.van Kooten C, Banchereau J. CD40-CD40 ligand. Journal of leukocyte biology. 2000;67(1):2–17. doi: 10.1002/jlb.67.1.2. [DOI] [PubMed] [Google Scholar]
- 13.Havenar-Daughton C, Lindqvist M, Heit A, et al. CXCL13 is a plasma biomarker of germinal center activity. Proc Natl Acad Sci U S A. 2016 doi: 10.1073/pnas.1520112113. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 14.Simpson N, Gatenby PA, Wilson A, et al. Expansion of circulating T cells resembling follicular helper T cells is a fixed phenotype that identifies a subset of severe systemic lupus erythematosus. Arthritis Rheum. 2010;62(1):234–244. doi: 10.1002/art.25032. [DOI] [PubMed] [Google Scholar]
- 15.Pallikkuth S, Sharkey M, Babic DZ, et al. Peripheral T Follicular Helper Cells Are the Major HIV Reservoir within Central Memory CD4 T Cells in Peripheral Blood from Chronically HIV-Infected Individuals on Combination Antiretroviral Therapy. J Virol. 2015;90(6):2718–2728. doi: 10.1128/JVI.02883-15. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 16.Kaufmann DE, Kavanagh DG, Pereyra F, et al. Upregulation of CTLA-4 by HIV-specific CD4+ T cells correlates with disease progression and defines a reversible immune dysfunction. Nat Immunol. 2007;8(11):1246–1254. doi: 10.1038/ni1515. [DOI] [PubMed] [Google Scholar]
- 17.Connors M, Kovacs JA, Krevat S, et al. HIV infection induces changes in CD4+ T-cell phenotype and depletions within the CD4+ T-cell repertoire that are not immediately restored by antiviral or immune-based therapies. Nature medicine. 1997;3(5):533–540. doi: 10.1038/nm0597-533. [DOI] [PubMed] [Google Scholar]
- 18.Day CL, Kaufmann DE, Kiepiela P, et al. PD-1 expression on HIV-specific T cells is associated with T-cell exhaustion and disease progression. Nature. 2006;443(7109):350–354. doi: 10.1038/nature05115. [DOI] [PubMed] [Google Scholar]
- 19.Cubas RA, Mudd JC, Savoye AL, et al. Inadequate T follicular cell help impairs B cell immunity during HIV infection. Nature medicine. 2013 doi: 10.1038/nm.3109. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 20.Hong JJ, Amancha PK, Rogers K, Ansari AA, Villinger F. Spatial alterations between CD4(+) T follicular helper, B, and CD8(+) T cells during simian immunodeficiency virus infection: T/B cell homeostasis, activation, and potential mechanism for viral escape. J Immunol. 2012;188(7):3247–3256. doi: 10.4049/jimmunol.1103138. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 21.Klatt NR, Vinton CL, Lynch RM, et al. SIV infection of rhesus macaques results in dysfunctional T- and B-cell responses to neo and recall Leishmania major vaccination. Blood. 2011;118(22):5803–5812. doi: 10.1182/blood-2011-07-365874. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 22.Moir S, Ogwaro KM, Malaspina A, et al. Perturbations in B cell responsiveness to CD4+ T cell help in HIV-infected individuals. Proc Natl Acad Sci U S A. 2003;100(10):6057–6062. doi: 10.1073/pnas.0730819100. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 23.Nicholas KJ, Zern EK, Barnett L, et al. B cell responses to HIV antigen are a potent correlate of viremia in HIV-1 infection and improve with PD-1 blockade. PLoS One. 2013;8(12):e84185. doi: 10.1371/journal.pone.0084185. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 24.Crawford A, Angelosanto JM, Kao C, et al. Molecular and Transcriptional Basis of CD4(+) T Cell Dysfunction during Chronic Infection. Immunity. 2014;40(2):289–302. doi: 10.1016/j.immuni.2014.01.005. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 25.Wherry EJ. T cell exhaustion. Nat Immunol. 2011;12(6):492–499. doi: 10.1038/ni.2035. [DOI] [PubMed] [Google Scholar]
- 26.Petrovas C, et al. CD4 T follicular helper cell dynamics during SIV infection. The Journal of Clinical Investigation. 2012;122(9):3281–3294. doi: 10.1172/JCI63039. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 27.Lindqvist M, et al. Expansion of HIV-specific T follicular helper cells in chronic HIV infection. The Journal of Clinical Investigation. 2012;122(9):3271–3280. doi: 10.1172/JCI64314. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 28.Moir S, Fauci AS. B cells in HIV infection and disease. Nat Rev Immunol. 2009;9(4):235–245. doi: 10.1038/nri2524. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 29.Kalams SA, Parker SD, Elizaga M, et al. Safety and comparative immunogenicity of an HIV-1 DNA vaccine in combination with plasmid interleukin 12 and impact of intramuscular electroporation for delivery. The Journal of infectious diseases. 2013;208(5):818–829. doi: 10.1093/infdis/jit236. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 30.Li F, Malhotra U, Gilbert PB, et al. Peptide selection for human immunodeficiency virus type 1 CTL-based vaccine evaluation. Vaccine. 2006;24(47–48):6893–6904. doi: 10.1016/j.vaccine.2006.06.009. [DOI] [PubMed] [Google Scholar]
- 31.Rossio JL, Esser MT, Suryanarayana K, et al. Inactivation of human immunodeficiency virus type 1 infectivity with preservation of conformational and functional integrity of virion surface proteins. J Virol. 1998;72(10):7992–8001. doi: 10.1128/jvi.72.10.7992-8001.1998. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 32.Rutebemberwa A, Bess JW, Jr, Brown B, et al. Evaluation of aldrithiol-2-inactivated preparations of HIV type 1 subtypes A, B, and D as reagents to monitor T cell responses. AIDS research and human retroviruses. 2007;23(4):532–542. doi: 10.1089/aid.2006.0136. [DOI] [PubMed] [Google Scholar]
- 33.Sather DN, Armann J, Ching LK, et al. Factors associated with the development of cross-reactive neutralizing antibodies during human immunodeficiency virus type 1 infection. J Virol. 2009;83(2):757–769. doi: 10.1128/JVI.02036-08. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 34.Li Y, Migueles SA, Welcher B, et al. Broad HIV-1 neutralization mediated by CD4-binding site antibodies. Nature medicine. 2007;13(9):1032–1034. doi: 10.1038/nm1624. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 35.Li M, Gao F, Mascola JR, et al. Human immunodeficiency virus type 1 env clones from acute and early subtype B infections for standardized assessments of vaccine-elicited neutralizing antibodies. J Virol. 2005;79(16):10108–10125. doi: 10.1128/JVI.79.16.10108-10125.2005. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 36.Blish CA, Jalalian-Lechak Z, Rainwater S, Nguyen MA, Dogan OC, Overbaugh J. Cross-subtype neutralization sensitivity despite monoclonal antibody resistance among early subtype A, C, and D envelope variants of human immunodeficiency virus type 1. J Virol. 2009;83(15):7783–7788. doi: 10.1128/JVI.00673-09. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 37.Perreau M, Savoye AL, De Crignis E, et al. Follicular helper T cells serve as the major CD4 T cell compartment for HIV-1 infection, replication, and production. J Exp Med. 2013;210(1):143–156. doi: 10.1084/jem.20121932. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 38.Sallusto F, Lenig D, Forster R, Lipp M, Lanzavecchia A. Two subsets of memory T lymphocytes with distinct homing potentials and effector functions. Nature. 1999;401(6754):708–712. doi: 10.1038/44385. [DOI] [PubMed] [Google Scholar]
- 39.Oswald-Richter K, Grill SM, Leelawong M, et al. Identification of a CCR5-expressing T cell subset that is resistant to R5-tropic HIV infection. PLoS Pathog. 2007;3(4):e58. doi: 10.1371/journal.ppat.0030058. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 40.Mikell I, Sather DN, Kalams SA, Altfeld M, Alter G, Stamatatos L. Characteristics of the earliest cross-neutralizing antibody response to HIV-1. PLoS Pathog. 2011;7(1):e1001251. doi: 10.1371/journal.ppat.1001251. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 41.Schultz BT, Teigler JE, Pissani F, et al. Circulating HIV-Specific Interleukin-21(+)CD4(+) T Cells Represent Peripheral Tfh Cells with Antigen-Dependent Helper Functions. Immunity. 2016;44(1):167–178. doi: 10.1016/j.immuni.2015.12.011. [DOI] [PubMed] [Google Scholar]
- 42.Godefroy E, Zhong H, Pham P, Friedman D, Yazdanbakhsh K. TIGIT-positive circulating follicular helper T cells display robust B-cell help functions: potential role in sickle cell alloimmunization. Haematologica. 2015;100(11):1415–1425. doi: 10.3324/haematol.2015.132738. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 43.Good-Jacobson KL, Szumilas CG, Chen L, Sharpe AH, Tomayko MM, Shlomchik MJ. PD-1 regulates germinal center B cell survival and the formation and affinity of long-lived plasma cells. Nat Immunol. 2010;11(6):535–542. doi: 10.1038/ni.1877. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 44.Haynes NM, Allen CD, Lesley R, Ansel KM, Killeen N, Cyster JG. Role of CXCR5 and CCR7 in follicular Th cell positioning and appearance of a programmed cell death gene-1high germinal center-associated subpopulation. J Immunol. 2007;179(8):5099–5108. doi: 10.4049/jimmunol.179.8.5099. [DOI] [PubMed] [Google Scholar]
- 45.Trautmann L, Janbazian L, Chomont N, et al. Upregulation of PD-1 expression on HIV-specific CD8+ T cells leads to reversible immune dysfunction. Nature medicine. 2006;12(10):1198–1202. doi: 10.1038/nm1482. [DOI] [PubMed] [Google Scholar]
- 46.Porichis F, Hart MG, Griesbeck M, et al. High-throughput detection of miRNAs and gene-specific mRNA at the single-cell level by flow cytometry. Nat Commun. 2014;5:5641. doi: 10.1038/ncomms6641. [DOI] [PMC free article] [PubMed] [Google Scholar]
Associated Data
This section collects any data citations, data availability statements, or supplementary materials included in this article.