
Figure 1.
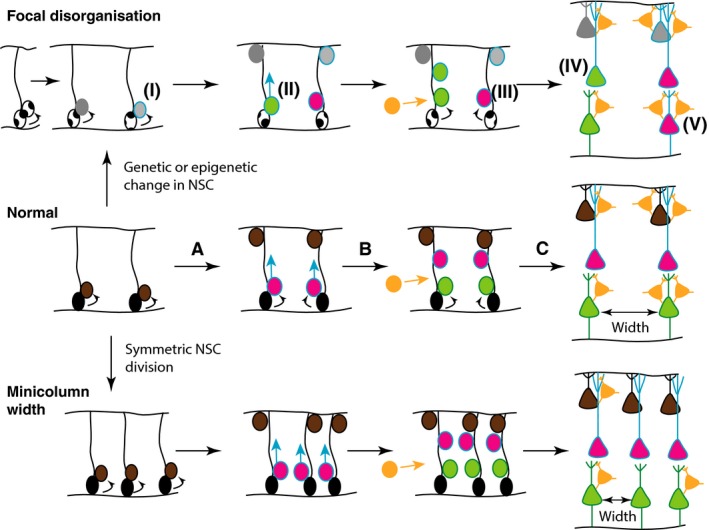
Hypothesis of potential mechanisms underlying focal disorganisation and altered minicolumn distribution in ASD cortex. Normal: During normal cortical development (middle panel), radial glial NSCs (black) located at the ventricular zone, with radial processes extending to the pial surface of the neural epithelium, can undergo symmetrical (to produce more radial glial NSCs) or asymmetric divisions to maintain the NSC pool and generate committed neural progenitors (brown) (A). The latter migrate towards the pial surface on the radial scaffold, where they differentiate into projection neurons. This process continues with different classes of neurons (pink, green), being produced in successive waves over developmental time (B), so that different neuronal types are grouped together in different cortical layers (C). GABAergic interneurons (orange) migrate into the neuroepithelium (B), and integrate into the circuitry (C). At the end of this process, projection neurons are organised in radial minicolumns, a reminder of their developmental origin. Focal disorganisation: We hypothesise that focal disruptions may be the result of early genetic or epigenetic changes in NSCs (i), such that the resulting neuronal progenitors have defects in differentiation (ii) or migration (iii) such that they adopt the wrong fate (iv), or end up in an in appropriate position in the brain (v). The end result is a cortex with apparent disorganisation of neuronal cell types and perhaps even distorted cell densities. Minicolumn width: We hypothesise that alterations in symmetric NSC divisions such that the early density of radial scaffolds is increased, will result in increased density of minicolumns. Alternatively, the migration and integration of GABAergic interneurons and other independent mechanisms might affect the spacing of minicolumns.