

Abstract
Artificial cells have attracted much attention as substitutes for natural cells. There are many different forms of artificial cells with many different definitions. They can be integral biological cell imitators with cell-like structures and exhibit some of the key characteristics of living cells. Alternatively, they can be engineered materials that only mimic some of the properties of cells, such as surface characteristics, shapes, morphology, or a few specific functions. These artificial cells can have applications in many fields from medicine to environment, and may be useful in constructing the theory of the origin of life. However, even the simplest unicellular organisms are extremely complex and synthesis of living artificial cells from inanimate components seems very daunting. Nevertheless, recent progress in the formulation of artificial cells ranging from simple protocells and synthetic cells to cell-mimic particles, suggests that the construction of living life is now not an unrealistic goal. This review aims to provide a comprehensive summary of the latest developments in the construction and application of artificial cells, as well as highlight the current problems, limitations, challenges and opportunities in this field.
Introduction
Since the discovery in 1665 by Robert Hooke, cells have been studied for several centuries [1]. The cell theory, which was formally proposed in 1839 by Matthias Jakob Schleiden and Theodor Schwann, stated that cell is the basic structural and functional unit of all known living organisms [1]. It is the fundamental ‘building block of life’. Understandably, modern cell biology is not satisfied by just investigating the structure, functions and working principles of cells. The research has expanded into many new areas such as the origin of life, cell engineering, biotechnology, bio-factories, medicine, drug delivery, pharmaceuticals, biosensors, and bioremediation [2]. However, along with the rapid development of cell biology, many issues have arisen mainly because of the inherent complexity of biological cells as well as the frangibility, that is to say, easy loss of activity or death in vitro. To overcome these issues while still mimicking biological cells, artificial cells are built [3] which are expected to be more easily controlled and are more robust than natural cells. The concept of artificial cells was first proposed by Dr. Thomas Ming Swi Chang in 1957 [4]. They can be used as biomimetic systems to study and understand properties of biological cells, to investigate the dynamics of cells with minimal interference from cellular complexity, and to explore new possible applications in place of biological cells.
Artificial cells can be defined in many ways. In this review, they are classified into two main categories based upon their intrinsic characteristics – typical and non-typical. The typical ones are artificial cells in ‘full-sense’. Strictly speaking, the typical artificial cells should have cell-like structures and exhibit at least some of the key characteristics of living biological cells, such as to evolve, to self-reproduce and to metabolize [3,5,6]. The non-typical artificial cells, on the other hand, are engineered materials that mimic one or more features of biological cells and most importantly, have no restrictions in structure. The more precise definition of these materials would be ‘cell mimics’ [7] that mimic some functions, surface characteristics, shapes, and even morphology of biological cells.
This review focuses on recent advances in the design of artificial cells, provides an overview of their current development status, as well as highlights the various applications and limitations of artificial cells.
Typical artificial cells
The construction of typical artificial cells is considered as one of the pillars of synthetic biology [8,9]. Research on these synthetic cells has many purposes, such as (i) providing a way to investigate and understand cellular life; (ii) connecting the non-living with the living world; (iii) adding new functions which are absent in biological cells for the development of new applications; (iv) providing plausible theory for the origin of life.
Typical artificial cells should ideally have similar structures and essential properties of living cells [3,10]. The ultimate goal is to construct artificial cells that can be considered as ‘alive’. While defining ‘alive’ is rather difficult, there is some consensus in biology. Generally speaking, when we describe an organism as alive, it means that it can self-maintain, self-reproduce, evolve, and die. However, even the simplest known organisms are very complex, making the efforts to synthesize ‘living’ artificial cells arduous and very challenging. Biological cells possess three main components for performing the essential functions of life [11]:
A stable, semi-permeable membrane that encloses cell constituents protecting them from being damaged by the external environment while allowing selective material and energy exchanges.
Biomacromolecules (DNA or RNA) that carry the genetic information, control the dynamics of the cell, and endow it with the capability of evolution.
A series of metabolic pathways used for providing energy to cells, to make them self-maintain and self-renew, as well as self-process information.
It is highly desirable that artificial cells possess all three features of biological cells. Although an artificial cell that possesses all basic properties of a living cell has not been created so far, recent advancements indicate that it is now a realistic goal.
Up till now, two main fundamental approaches have been considered for the construction of an artificial cell: A top-down approach and a bottom-up approach (Fig. 1) [12]. The top-down approach starts from a living organism, stripping down the genome to the lowest number of genes that are required to maintain the essential properties of the cellular life, or totally replacing the genome with a synthetic one [13]. In contrast, the bottom-up approach starts from scratch [14]. It constructs a ‘living’ artificial cell by assembling biological and/or non-biological molecules. These two approaches are very different but complementary to each other, fabricating a broad range of artificial cells from a simple protocell to an engineered living life.
FIGURE 1.

Approaches for the design and construction of artificial cells: In the top-down approach, artificial cells are created by stripping or replacing the genomes of living organisms (cells, bacteria or viruses), reducing their complexity, and only retaining minimum substances to maintain the essential life. In the bottom-up approach, artificial cells are constructed by assembling non-living components to form an integral that can replicate essential properties of natural cells.
Top-down approach
The top-down construction of artificial cells focuses on building ‘minimal’ cells by reducing or simplifying the genome of a living cell. A minimal cell means a theoretical cell that only has the minimum number of genes needed to perform the most essential life function, that is, to survive [15]. It is expected that minimal cells will help us understand and regulate essential life processes. In 1995, Venter and colleagues discovered that the parasitic bacterium Mycoplasma genitalium has only 517 genes, which is among the simplest living organisms known [16]. Such a small genome was an inspiration for scientists to develop ‘minimal cells’. When non-essential genes were knocked down to examine the cell’s viability [17], it turned out that approximately 256–350 genes were sufficient to maintain the life of a cell [16,18–24]. Subsequently, in 2004, Gil et al. redefined the minimal gene number [21] for cell viability. Based on both computational methods and experimental strategies, they proposed that the minimal gene set to maintain a functional living bacterial cell could be reduced to 206 genes. However, among these 206 genes, there are still many genes with part of non-essential functions, suggesting that the number can still be further reduced [25]. For example, some of the enzymes that are responsible for the synthesis of low molecular weight compounds (nucleotides and amino acids) can be dispensable. The corresponding genes can therefore be knocked down and their products can be supplied from the surrounding medium [26]. This brings the gene number down to about 150. However, the survival of these minimal cells requires that the corresponding compounds are available in the environment and the cell membranes are permeable to these compounds. This indicates that the definition of minimal cells is also related to the type of environment they are in. The resources in surroundings determine which genes they can lose. In addition, with respect to the outside supply of substrates, these minimal cells could maintain their basic cellular life. However, because they cannot synthesize these substrates by themselves, they may lose some of self-repair abilities. For example, if the genes for lipid synthesis are knocked out and the lipid molecules are supplied by environment instead, the artificial cell membranes will lose self-producibility and cell division will simply be due to a physical dynamics process.
Although effective, knocking out genes one at a time is cumbersome and the process is rather ineffective. There is a more advanced and challenging way to construct top-down artificial cells by replacing the original genes of the biological cells with synthetic ones. Although artificial eukaryotic cells have not been successfully created by this method so far, there were several reports about the construction of some primary organisms, such as viruses and bacteria. In 2002, Cello et al. reported the creation of an artificial infectious poliovirus [27]. They synthesized the full-length poliovirus DNA (cDNA) de novo from basic chemical building blocks. The synthetic cDNA was then transcribed into highly infectious viral RNA with the T7 RNA polymerase. Subsequently, the viral RNA was translated and replicated in cytoplasmic extracts of uninfected cells, generating poliovirus with physiological and pathological properties similar to those of the natural virus. Although viruses are not considered as typical lives, this report demonstrated the possibility of synthesizing more complex living organisms by chemical/biochemical methods without using a genetic template.
In 2010, Venter et al. [13] designed computer-based genome sequences named ‘M. mycoides JCVI-syn1.0’ on two laboratory strains of Mycoplasma mycoides subspecies capri GM12, the genome sequences of which were already decoded. The designed genomes were somewhat different from the original ones with some genes altered, added or deleted. The genes were chemically synthesized, stitched in yeast, and then transplanted into Mycoplasma capricolum recipient cells. The resulting new cells had expected phenotypic properties of M. mycoides and could self-replicate (Fig. 2). These cells were referred to as ‘synthetic cells’. The designed synthetic DNA acted as the genetic material in these artificial cells. Although there are some philosophical and ethical controversies, the work undoubtedly represents a new height of creating artificial cells and controlling over life at the molecular level.
FIGURE 2.
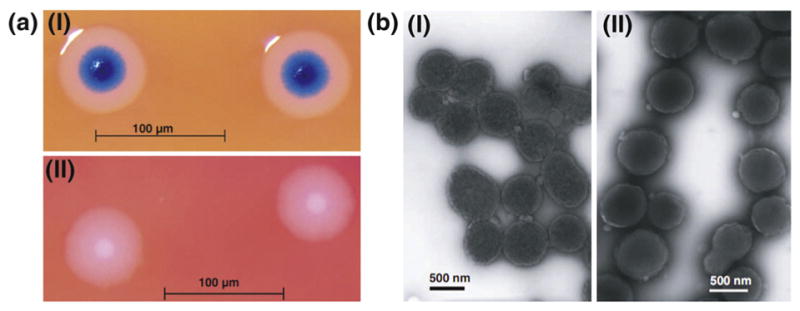
(a) Images of (I) M. mycoides JCVI-syn1.0 (synthesized cells) and (II) wild-type (WT) M. mycoides: The two cell colonies display similar fried egg-like morphology. The blue color of JCVI-syn1.0 colonies is due to X-gal staining. The WT cells do not contain the lacZ gene and hence no blue color. (b) TEM images of (I) M. mycoides JCVI-syn1.0 and (II) WT M. mycoides. The two cell colonies show similar ovoid morphology [13]. Copyright © 2010, The American Association for the Advancement of Science.
Synthetic biology has enabled the construction of large DNA sequences. These chemically assembled genomes provide the necessary genetic information to create new organisms with self-replicating ability. This not only helps to understand minimal cellular life, but also provides a variety of new functions with potential applications such as in the production of pharmaceuticals and energy. Despite these advances, there is still a long way to go to engineer, alter, and create a more complex artificial life. As described above, so far, only minimal low organisms (viruses, bacteria) were constructed, whereas minimal eukaryotic cells, due to the complexity of their genomes, have not been successfully created. The accuracy of genome engineering is also a problem. Although modern genomic technology is substantially improved in speed and accuracy, obtaining an error-free genome is still a challenge. Also, the high cost for genome analysis, modification, and synthesis is an obstacle for broad application of these artificial cells. Therefore, improving the accuracy and lowering the cost of genome engineering are urgently needed for the future development of artificial cells by the top-down method.
Bottom-up approach
Compared with the top-down strategy, the bottom-up approach is much more challenging. Instead of starting from existing living organism, it aims to create a cell by assembling a stack of non-biotic components [28]. Actually, the original life is supposed to have initiated from the non-living blocks in a step by step procedure. Constructing artificial cells by the bottom-up approach may set up bridges between non-living and living worlds, as well as help to understand the origin of life. Furthermore, the bottom-up method can overcome some of the shortcomings of the top-down approach. Modifying existing organisms by stripping their genes one by one or replacing them wholly is cumbersome, and may bring some unpredictable and undesirable results. For example, when a modern cell is engineered to produce some chemicals, drugs or biofuels, altering its metabolism may affect its fitness resulting in high mortality and low production rates, as well as generate some unwanted by-products. Also, the products may have inherent toxicity to the host cells. These shortcomings are unfavorable to their biological applications. On the favorable side, the systems designed from the bottom-up approach containing only minimal components required to perform the desired functions are simpler and more controllable and can therefore overcome the limits of the top-down method.
Three basic elements are needed for the construction of a ‘living’ artificial cell using a bottom-up approach. These include information-carrying molecules, cell membranes, and metabolism systems [11,29]. The information molecules, RNA or DNA, define the nature and function of the cell. Metabolism generates energy for cell operation, growth, division, and evolution. Finally, the third essential element, the membrane, provides a habitat for cellular molecules and also serves as a mediator to communicate with the outside world and as a transporter for material exchange. The detailed descriptions of these three parts are outlined below.
It is well known that in most of the cells, genetic information is carried in the form of double-stranded DNA. The hereditary information can be copied by template-directed polymerization. The genetic information can be transcribed from DNA into RNA, and then translated from RNA into proteins.
During the artificial cell design process, genes are defined as the basic biological units. It is now technically possible to synthesize genomes with dozens of genes in the laboratory [30]. After logical assembly and encapsulation in a vesicle, these custom-made DNA program units can be used as softwares to develop the functions of the artificial cells. To make the genetic circuits work, promoters (e.g., endogenous T7 promoter from bacteriophage) and different transcription factors are also needed, as well as ribosomes, some enzymes, and low molecular weight compounds [31]. The translation process involves many more components, about one hundred, and the reaction conditions depend on very precise adjustments of some parameters, such as salt concentration, temperature, and pH [30,32,33]. Although challenging, the cell-free expression within micro-vesicles has been successfully demonstrated in vitro [34,35].
Metabolism is the set of life-sustaining chemical transformations within the cells. This process provides energy for cells and allows them to grow and reproduce, synthesize and digest, transfer information and substances, maintain structures, and respond to their environment. These metabolic processes are catalyzed by a set of enzymes. To drive metabolic reactions, a source of chemical energy is also required. The energy can be supplied in the form of ATP or other such molecules [36]. In most cases, these energy molecules are supplied from the environment, which requires that the artificial cell membranes are permeable [3].
All known cells have plasma membranes to separate the intra-cellular substances from the outside environment. Cell membranes are primarily composed of a mixture of lipids and functional proteins. These membranes play a variety of roles including, (1) protecting the intracellular substances from the external environment, (2) letting the nutrients in and the wastes out, while blocking unnecessary molecules, (3) providing communication channels with the surroundings and (4) modulating many cell processes such as adhesion and migration. Although biological cell membranes are very complex and hard to mimic, the artificial cell membranes are expected to be constructed to possess some of the critical features of natural ones.
Phospholipid membrane
As noted by their identity as the main components of plasma membranes of almost all known living organisms, phospholipids are the primary molecules for constructing artificial cell membranes. Phospholipids are double-chained amphiphiles, possessing a hydrophilic phosphate ester head and two hydrophobic acyl tails. In water, they can spontaneously self-assemble into spherical vesicles (liposomes) with bilayer membranes. There are various methods for preparing liposomes of different sizes ranging from nanometers to micrometers to suit the desired size of a particular cell [37,38]. The cellular components, either from nature or by synthesis, can be included in the vesicles to build artificial cells. So far, many attempts have been made to use liposomes as containers for carrying out specific cell processes.
Production of bio-macromolecules, such as DNA, RNA, and proteins, is the primitive form of cell metabolism. Thus, realizing the synthesis of biomolecules inside liposomes can bring these lipid-based artificial cells closer to biological cells. An early report [39] showed that by encapsulating template-independent RNA polymerase (polynucleotide phosphorylase) inside lipid vesicles, long-chain RNA polymers (poly(A)) could be synthesized within these vesicles. Intermediate chain length lipids were chosen for entrapping the enzymes while allowing ionic substrates exchange with outside environment. Shortly thereafter, the first example of polymerase chain reaction (PCR) carried out inside liposomes was reported [40]. The liposomes were composed of 1-palmitoyl-oleoyl-sn-glycero-3-phosphocholine (POPC), or of a mixture of POPC and phosphatidylserine (PS) and were demonstrated to be stable at the high temperature (95°C) used for PCR. The integrity of the aqueous contents was well maintained during the long time course of PCR. The ability to perform PCR within liposomes is attractive as PCR is not only important in clinical applications and basic research but may also be of relevance for the origin of life [41]. It has been suggested that life might have originated in a hydrothermal environment [42,43], in which the spatial temperature gradient provided conditions for primitive genetic molecules (DNA, RNA) to self-replicate and proliferate in a PCR-like fashion [41,44,45].
Protein biosynthesis can also be carried out in liposomes. Oberholzer et al. [46] reported that by incorporating the ribosomal subunits, messenger RNA (poly(U)), tRNAPhe, the phenylalanyl-tRNA synthetase, some elongation factors, and the substrate phenylalanine or 14C-labeled phenylalanine within the POPC liposomes, the peptide poly(Phe) was successfully synthesized. Furthermore, the efficiency of the polypeptide synthesis was quite high.
In recent years, giant vesicles (GVs) have attracted much attention as biochemical reactors for macromolecule synthesis [47–49]. The advantages of GVs used for artificial cell membranes are manifold compared with conventional small or large-sized unilamellar vesicles (SUVs or LUVs). GVs formed by electroformation have typical diameters of 30–100 μm, which are similar to those of biological membranes while the conventional SUVs or LUVs have approximately 100–1000 times smaller diameters. The giant sizes of GVs endow them with many novel properties. For example, they can be punctured by a micropipette without being destroyed. As a result, substances can be microinjected accurately [50,51] and the species and concentrations of the substances can also be precisely controlled. On the contrary, in the case of conventional liposomes, the substances are encapsulated randomly during the vesicle synthesis process, which is uncontrollable and may lead to concentration inhomogeneity. Permeability is another important aspect. It has been demonstrated that the membranes of GVs formed by electroformation allow penetration of external nucleases. These merits make GVs suitable for imitating the cytological processes of biological cells [52,53].
Besides synthesis of bio-macromolecules within lipid vesicles, a minimal cell should be capable of performing more complex functions. Gene expression is one of the advanced properties of a living cell that allows the information to be transferred from a gene to the functional products. These products are often proteins, and in some cases, they are functional RNAs. Gene expression is the most fundamental level that gives rise to the organism’s phenotype. Obviously, it is an enormous engineering challenge to construct complex gene expression networks inside a synthetic liposome. There have been some promising experimental approaches to express a single protein product or simple two-stage genetic network within liposomes.
Yu et al. [35] reported the expression of a mutant GFP, GFPmutl, in the giant lipid vesicles (eggPC/cholesterol/DSPE-PEG5000). For cell-free transcription and translation, the protein expression reaction mixture containing low molecular weight cofactors and substrates (ATP, GTP, UTP, CTP, 20 amino acids), T7 RNA polymerase, cell extract, and GFPmutl-encoding gene (plasmid) were entrapped in the liposomes and incubated. An intense fluorescence inside the liposomes could be detected, showing that not only the protein expression successfully occurred, but also the protein was as functional as that expressed in biological cells. Subsequently, expression of rsGFP (red-shifted green fluorescent protein) within individual cell-sized giant vesicles was observed by Nomura et al. [54]. They also demonstrated that the vesicles were able to prevent the synthesized rsGFP from being degraded by the externally added proteinase K.
After accomplishing the expression of a single protein inside liposomes, the next step was to increase the genetic complexity by creating a functional genetic network. Ishikawa et al. [55] constructed a two-stage genetic cascade in liposomes. A T7 RNA polymerase gene was used for the first stage and a GFP gene for the second stage. The product of the first stage (T7 RNA polymerase) was a promoter for driving the GFP protein synthesis of the second stage; thus, these two genes formed a transcriptional activation cascade network.
Noireaux and Libchaber [56] fused the gene of enhanced GFP (eGFP) with the gene of a pore-forming protein from Staphylococcus aureus, α-hemolysin, and tested their expression within egg lecithin liposomes. The results showed that co-expression of α-hemolysin resulted in pore formation in the lipid membranes resulting in an increase in the permeability of nutrients. This, to some extent, solved the energy and material limitations of the biochemical reactor and increased the eGFP expression time for up to four days without vesicle burst.
Kuruma et al. [57] successfully produced two membrane proteins, sn-glycerol-3-phosphate acyltransferase (GPAT) and lyso-phosphatidic acid acyltransferase (LPAAT), by the PURE (protein synthesis using recombinant elements) system encapsulated in liposomes. The two enzymes could catalyze a cascade of reactions that produced phosphatidic acid in situ inside the liposomes. This was the first evidence that integral membrane proteins could be synthesized inside liposomes, and their activities could be successfully detected. However, the forced discontinuity between two reactions and the low product yield, as well as the poor membrane permeability of water-soluble substrates, made it difficult to directly observe morphological changes in liposomes. In order to overcome these problems, the authors proposed some resolutions which included building transporters inside lipid membranes, reforming the enzyme by a genetically engineered method. This study might provide a new way for the construction of self-reproducing artificial cells in which the lipid membrane can be self-synthesized.
To be considered as alive, an artificial cell should not only have essential cellular processes such as bio-macromolecule synthesis and gene expression mentioned above but also display more complex behaviors that mimic biological cells and new functions that are absent in nature. Communication, both with the environment and with each other, is a very important high-level behavior of cells. These functions allow cells to adapt to the environmental change and work co-operatively to perform important bodily behaviors that are necessary to survive.
Gardner et al. [58] developed a lipid-bound protocell that could synthesize complex carbohydrates by the autocatalytic sugar-synthesizing formose reaction, which was able to initiate a quorum-sensing mechanism in the marine bacterium Vibrio harveyi and cause a bioluminescent response. The construction and working principles of the protocell were illustrated in Fig. 3a. Concentrated formaldehyde feedstock and metal (calcium) ions were encapsulated within the robust lipid vesicles. When the pH increases, the formose catalyst, calcium hydroxide, was formed in situ within the vesicles, which catalyzed the production of carbohydrate complexes. The primary products, carbohydrate–borate complexes, were analogs of signaling molecules in the quorum-sensing system of V. harveyi. When the products diffused out across the vesicle membranes, they were detected by the marine bacterium V. harveyi and provoked a quorum-sensing response, resulting in a protein phosphorylation mechanism and the subsequent expression of bioluminescent proteins. This report indicated the capability of artificial cells to display complex behaviors such as communication with biological cells.
FIGURE 3.

(a) Illustration of the working principle of sugar-producing protocells. The formose reaction is carried out inside a lipid vesicle, which is driven by the increased pH outside (top left). The product, carbohydrate–borate, is formed and diffuses from the vesicle into the medium (top right). The diffused products are then detected by the bacterium V. harveyi and bind to LuxP/LuxQ signal transduction protein, resulting in a protein phosphorylation response and the subsequent expression of the luxCDABE gene (bottom right) with detectable bioluminescence (bottom left) [58]. Copyright © 2009, Nature Publishing Group. (b) Artificial cells translate chemical signals for E. coli. The molecule theophylline cannot be detected by E. coli (top). Through the artificial cell system, theophylline can be translated into isopropyl β-D-1-thiogalactopyranoside (IPTG), which can be sensed by E. coli and induce its response (bottom) [59]. Copyright © 2014, Nature Publishing Group.
Lentini et al. [59] described an artificial cell system that could expand the senses of E. coli (Fig. 3b). This system can translate a chemical compound, theophylline, which cannot be sensed by E. coli in nature, to a molecule, isopropyl β-D-1-thiogalactopyrano-side (IPTG) that E. coli can naturally respond to. The artificial cell was engineered with a lipid vesicle containing theophylline-sensing genetic devices and the molecule IPTG. In the genetic devices, a theophylline riboswitch was fused with the gene of pore-forming protein α-hemolysin. When the external theophylline molecules were diffused into the artificial cells, they could bind to riboswitch and induce the transcription–translation processes of genes, resulting in the expression of α-hemolysin. The protein α-hemolysin could form unspecific pores in the lipid membranes and caused the release of IPTG. Thus, when theophylline molecules were added to the mixture of E. coli and artificial cells, they could not be sensed by E. coli. However, they could trigger the release of IPTG molecules from the artificial cells, which could be detected by E. coli cells and induce both engineered and natural pathways in these bacterial cells.
Fatty acid membrane
Fatty acids are single-chained amphiphilic molecules with a long, saturated or unsaturated hydrophobic aliphatic tail and a hydrophilic carboxylic acid head. The tails for the naturally occurring fatty acids contain about 4–28 carbon atoms. Similar to phospholipids, fatty acids can also form bilayers, vesicles, and micelles in water, but with much simpler structures. Currently, more and more studies use fatty acids as primitive membrane models to investigate protocells on early Earth [60,61].
Studies showed that although contemporary organisms have phospholipid bilayers as the primary components of cell membranes, there are many obstacles for them to be prebiological cell membranes. The first problem is the availability of the phospholipid-like compounds on the primitive Earth. Phospholipids might be synthesized from the mixture of fatty acids, glycerol, and phosphate under prebiotic conditions [62], however, the probability of having all these components co-presented simultaneously is highly questionable. The attention was therefore turned to simpler amphiphiles. Several studies showed that fatty acids could be formed under simulated primitive Earth conditions [63–65]. By gradual accumulation, small carbon-containing molecules such as methane could form hydrocarbons in primeval times and then deposit on the rocks. Analysis showed that unsaturated hydrocarbons did exist in the ancient rocks [66]. It was postulated that these hydrocarbon mixtures could then be brought to the surface through earth movements and subsequently carboxylated by atmospheric carbon oxides. Experiments simulating prebiotic hydrocarbon synthesis generally have confirmed this hypothesis, and have demonstrated the synthesis of fatty acids as well as homologous fatty alcohols and aldehydes [67]. Normal fatty acids with C6 to C18 carbons can also be synthesized from CO and H2 by Fischer–Tropsch-type reactions, using meteoritic nickel-iron catalyst under conditions that may have occurred in the primordial solar nebula or during degassing of the primitive Earth. In non-Earth resources, fatty acids (with 2–12 carbons) and other amphiphiles have been found as abundant components of organic mixture in Murchison meteorite [68–70].
The ability of fatty acids to self-assemble into vesicles was first reported by Gebicki and Hicks [71]. They reported that oleic acid, an unsaturated fatty acid, could form spheres at slightly alkaline conditions. These spheres either had complex interior structures with multilayer membranes or consisted of collections of small spheres within a common envelope. They believed that these vesicles could be used as original membrane models. Hargreaves and Deamer et al. [67,72,73] suggested that saturated fatty acids, which were present in the primeval sea in significant amounts, can form membrane structures. They also systematically studied the influence of pH, temperature and hydrocarbon chain-length on the membrane formations. They postulated that to form the vesicles, the hydrocarbon chains should be eight carbons or greater in length and the temperature should be above the phase transition temperatures of the fatty acid to keep them in a fluid state. Also, the pH should be close to the apparent pKa of the membrane-incorporated fatty acid to make carboxyl groups half-charged. Under these conditions, membranous vesicles are produced, presumably due to the hydrogen bond networks formed between ionized RCOO− and neutral acid HOOCR. At higher pH, only micelles form, whereas at lower pH oil droplets condense [74].
The cell membrane is the only physical barrier that connects the inner cellular substance with the outer environment. The selective permeability to outside molecules may determine the nature of the primitive protocells. For example, there are many potential possibilities for the compositions of the prebiotic genetic polymers, while the final form chosen by nature may not contribute to the stability, but to the substrate permeability.
Szostak’s group [75] tested the permeability of fatty acid or lipid membranes to a series of carbohydrates, including five pentoses, selected aldohexoses, ketohexoses, and carbon alditols. The results showed that ribose diffused across the lipid membrane 3- to 10-fold faster than other pentoses and hexoses. More interestingly, they found that permeability coefficient of ribose was 5-fold larger than that of its diastereomers. The dynamics simulations, proton NMR, and crystal structure analysis showed that the permeability is influenced by molecular conformation and flexibility rather than hydrophobicity. The selective permeability of model prebiotic membranes for ribose suggests an early kinetic advantage of ribose over other five-carbon sugars. This may explain why ribose is the sugar chosen to be the component of the nucleic acid backbone. Szostak et al. also found that the fatty acid allows the crossing of charged molecules such as nucleotides, while typical lipid bilayers are relatively impermeable to polar or ionic substrates, including nucleotides [76]. The passage of complex macromolecular nutrients is very significant to prebiotic protocells as they have not evolved sophisticated protein channels and pumps to help the selective transportation of these nutrients. The group also showed that adding Mg2+ can significantly increase the permeability of nucleotides through fatty acid membranes.
As mentioned above, different from contemporary cells, the early forms of primitive cells presumably lacked the complex transport system to facilitate the passage of ions, nutrients, and metabolites. Thus, the passive diffusion played an important role in maintaining the primitive cellular life and the permeability was critical in determining the fate of a protocell. The interesting permeability characteristics of fatty acid membranes for biomolecules make them a valuable model for protocell investigations.
One of the major challenges for the synthesized artificial cells is to make them accomplish some complex biological functions such as growth, division, and self-replication, using both genetic polymers and membranes [6]. Fatty acid vesicles, which have long been studied as a model system for protocell membranes, were found to be able to grow and divide in many ways.
The first study that clearly demonstrated the growth ability of fatty acid vesicles was reported by Luisi’s group [77]. By incorporating the protein, ferritin, and using cryo-transmission electron microscopy (cryo-TEM), they observed the growth and division of oleic acid vesicles. After treatment with oleic acid, spontaneous vesicle growth occurred either gradually by the incorporation of oleic acid molecules or micelles, or stepwise by fusion with other vesicles. When the size reached a critical value, the vesicles split because of the thermodynamic instability, thus dividing into smaller vesicles (Fig. 4).
FIGURE 4.

Modes of vesicle growth and division. Vesicle growth can occur by incorporation of fatty acid monomers, micelles or other vesicles. The size of the vesicle will increase and the vesicle may divide into smaller vesicles spontaneously due to the thermodynamic instability [6]. Copyright © 2001, Nature Publishing Group.
Szostak et al. [78] also reported a different pathway for fatty acid membrane growth and division. They described that after feeding with fatty acid micelles, the spherical multilamellar fatty acid vesicles could grow and transform into long thread-like vesicles (Fig. 5). The process was driven by the transient imbalance between surface area and volume growth. Then, the shear stress triggered the thread-like vesicles to divide into multiple daughter vesicles. The main advantage of this pathway is that the internal contents can be retained in the process of division.
FIGURE 5.
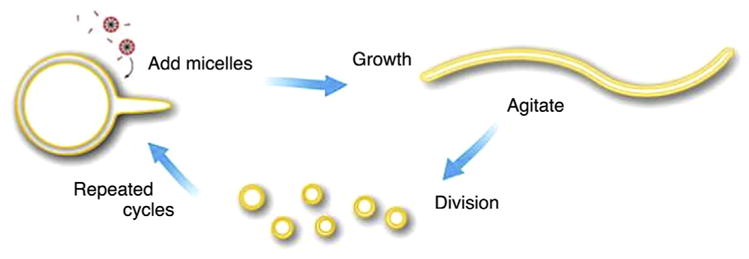
Schematic diagram of cyclic multilamellar vesicle growth and division. After being fed with micelles, the fatty acid vesicles grow in size. Because of the transient imbalance between surface area and volume growth, the initial spherical morphology eventually turns into long thread-like shapes. Then the thread-like vesicles divide into multiple daughter vesicles due to the shear stress [78]. Copyright © 2009 American Chemical Society.
As listed above, fatty acid vesicles are permeable to ribose and nucleotides, and they also have the ability to grow and divide. These features make them attractive candidates as the components of protocell membranes. Oberholzer et al. [79] constructed a model protocell consisting of 100 nm oleate/oleic acid small vesicle. They encapsulated the RNA template, viral RNA polymerase (Qβ repli-case), and the ribonucleotides ATP, CTP, GTP, and UTP inside the vesicles. Oleic anhydride was added externally, which can bind to the vesicle membranes and catalyze the growth and division of the vesicles. The Qβ replicase could catalyze the replication of RNA templates.
Szostak’s group [80] constructed a Mg2+ responsive protocell (Fig. 6a). The components of the protocell membrane were a mixture of myristoleic acid and its glycerol monoester. This membrane was Mg2+-tolerant and had Mg2+ permeability. Mg2+-dependent ribozyme molecules were encapsulated inside the pro-tocell. By adding external Mg2+, the ribozymes were activated and self-cleaved. The Mg2+ could also increase membrane permeability of small, negatively charged solutes, including uridine monophosphate, which facilitated RNA replication inside the vesicles. Membranes could also grow spontaneously because of the osmotically driven competition between vesicles and the addition of new micelles. This work was further extended by chelating Mg2+, which exhibited two main beneficial effects [83]. First, this protects the protocell membranes composed of pure fatty acids, such as oleic acid, from the disruption caused by Mg2+. And second, this protects the inner RNA oligonucleotides from Mg2+-catalyzed degradation, while the chelated Mg2+ can still catalyze template-directed RNA synthesis. Several chelators were tested including citrate, isocitrate, oxalate, nitrilotriacetic acid (NTA), and EDTA, with citrate being the most efficient. It is possible that in the prebiotic condition, some short acidic peptides were used as chelators instead of citrate, and the peptide-metal ion complexes might constitute the central components of cellular RNA poly-merases.
FIGURE 6.

(a) Illustration of Mg2+ responsive protocell. Mg2+ dependent ribozyme molecules are encapsulated inside fatty acid vesicles. By adding external Mg2+, the ribozymes can be activated and catalyzed RNA replication [80]. Copyright © 2005, American Chemical Society. (b) Clay-catalyzed protocell model. Nomarski optics images (I and III) and fluorescence images (II and IV) of clay-catalyzed fatty acid vesicles. (I and II) Negatively charged alumino-silicate ceramic microspheres (Zeeospheres) catalyze the formation of myristoleate vesicles. Arrowheads indicate Zeeospheres. (III and IV) Natural montmorillonite clay particles catalyze the formation of myristoleate vesicles [81]. Copyright ©2003, the American Association for the Advancement of Science. (c) The amplification of encapsulated DNA combines with self-reproduction of giant vesicles (GVs). (I) Illustration of the chemical link between amplification of DNA and self-reproduction of GVs. The amplification of DNA can be carried out within GVs. The amplified DNA can adhere to the inner leaflet of the GV, accelerating the self-reproduction of GV due to the imbalance between the inner and outer leaflets. (II) Real-time observation of morphological changes of DNA-amplified GVs after addition of membrane precursor (V*) [82]. Copyright ©2011, Nature Publishing Group.
Szostak et al. also proposed a striking clay-catalyzed protocell model, the formation, growth, and division of which were catalyzed by clay montmorillonite (Fig. 6b) [81]. Previous reports indicated that montmorillonite could catalyze the polymerization of RNA from activated ribonucleotides [84–86]. It was reported that montmorillonite could also accelerate the spontaneous conversion of fatty acid micelles into vesicles. The membranes formed in this way could, in turn, expand by absorbing additional fatty acids to encapsulate the mineral particles along with any additional substrates subsequently formed at the mineral surface, including oligonucleotides. The oligonucleotides could also be synthesized on the surface of encapsulated clay particles within the vesicles.
Although the protocell models involving self-reproduction of membrane and self-replication of information polymer have been constructed, the two processes were often carried out independently. Kurihara et al. [82] demonstrated, for the first time, that these processes could happen in parallel. They encapsulated DNA within a cationic GV and observed that the growth and division of the GVs were synchronized with the self-replication of DNA (Fig. 6c). The amplified DNA could interact with the vesicular membrane by electrostatic interactions and cause the instability and morphological change of the host GV resulting in the division. Also, the amplified DNA could be distributed into the daughter GVs. The result suggests the feasibility of DNA-membrane interaction [87], which presents a competitive protocell model for studying natural interactions in prebiotic processes.
Protein-polymer nano-conjugate membrane (proteinosome)
Protein molecules are attractive building blocks for the construction of membrane-bound compartments because of their intrinsic biocompatibility, biodegradability, and biofunctionality. Recently, Mann’s group [88] reported an interfacial assembly of protein-polymer nano-conjugates (proteinosomes) into stimulus-responsive biomimetic protocells. The conjugates were prepared by crosslinking of bovine serum albumin (BSA) and temperature-responsive polymer poly(N-isopropylacrylamide) (PNIPAAm) (Fig. 7). When dispersed in the Pickering emulsion, the conjugates were spontaneously assembled into micro-compartments in oil and could be transferred to a bulk water phase without losing the structural integrity. Such elastic compartments are semi-permeable, temperature-sensitive, and enzymatically active, making them good candidates for artificial cell construction. Furthermore, this proteinosome membrane is also very robust. It can withstand partial dehydration and rehydration, and remains structurally intact at a temperature of 70°C for 90 min. These features suggest that this membrane could be useful for studying the microscale confinement of thermophilic enzymes or for developing artificial cells capable of undertaking temperature cycling procedures, such as PCR-mediated amplification of entrapped genetic polymers.
FIGURE 7.
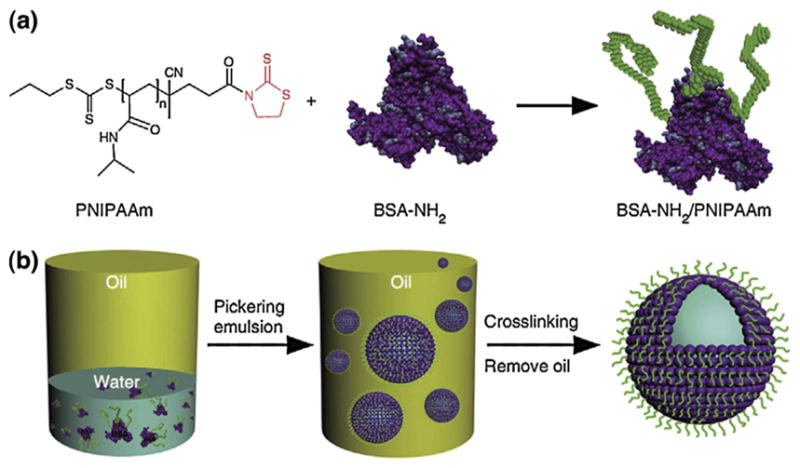
The procedures for preparation of proteinosomes. (a) The temperature sensitive polymer PNIPAAm is grafted onto the protein BSA via a mercaptothiazoline-amine reaction to form protein-polymer conjugates. (b) The conjugates self-assemble into vesicles in the Pickering emulsion. After removing the oil phase, the membrane of the proteinosome is cross-linked using PEG-bis(N-succinimidyl succinate) into a continuous membrane [88]. Copyright © 2013, Nature Publishing Group.
A wide range of functional components such as small molecule fluorescent dyes, drug molecules, proteins, enzymes and inorganic nanoparticles of various sizes can be encapsulated inside the proteinosomes [69]. An eGFP-expressing plasmid was constructed and added to the proteinosome synthesis solution with elements for cell-free gene expression. The plasmids were then encapsulated in situ inside the formed proteinosomes. The eGFP expression within the vesicles could be observed by fluorescence microscopy after two hours. It was also demonstrated that the membrane permeability can be thermally controlled. Myoglobin, a kind of peroxidases, was encapsulated inside the proteinosomes. The result showed that its activity was temperature-dependent. When the membrane of proteinosome was in a hydrated and bloated state at temperatures below the low critical solution temperature of PNIPAAm, it had increased permeability to the substrates thus increasing the activity of the enzyme. However, when the temperature was higher than the low critical solution temperature of PNIPAAm, the polymer membrane returned to its normal state and became hydrophobic. Also, its permeability to the substrate molecules diminished resulting in a sharp decrease of the reaction rate. Considering these multifunctional properties, the proteinsome-based artificial cells are expected to have potential applications in cytomimetic engineering.
Semipermeable membrane
It is known that in nature, enzymes are always compartmentalized within the organelles that have membranes to protect them while permeable to substrates and products. The concept of an ‘artificial cell’ using semipermeable microcapsules to mimic cells or organelles was first proposed by Chang, who developed a simple method for preparing semipermeable nylon microcapsules to encapsulate enzymes in situ [89]. The stable nylon microcapsules with a mean diameter of 15 μm were made by a simple interfacial polycondensation method. The stabilities and activities of enzymes and proteins encapsulated inside these capsules were tested. Chang reported that carbonic anhydrase from a red cell hemolysate retained its activity after several weeks and did not leak out. Similarly, hemoglobin could retain more than half of its oxygen-carrying capacity in a favorable preparation and storage environment. In vivo toxicological tests were also carried out and no significant toxicity was observed. Remarkably, injection of encapsulated urease increased the concentration of ammonia in the blood. The author then proposed that this may have a potential application in enzyme replacement therapy.
Later, Chang showed that besides hemoglobin and enzymes, the membranes could also encapsulate cells, microorganisms, adsorbents, magnetic materials and other biologically active materials (Fig. 8) [90–99] These artificial cells could protect the inner biologically active materials from directly contacting with external substrates while allowing smaller molecules, such as enzyme substrates, hormones, small peptides, and proteins, equilibrate across the membrane. Also, the permeability could be controlled over a wide range by using different types of membrane materials. The sizes of the artificial cells can also range from macro-dimensions to micro- and nano-dimensions and even down to molecular dimensions. So far, a number of potential medical applications using these artificial cells have been proposed, mainly for replacing or supplementing deficient cell functions [90–99].
FIGURE 8.

Various biologically active materials can be encapsulated in semipermeable polymer-based artificial cells, individually or in combination [90]. The semipermeable membrane can protect these materials from being degraded or affected by external environment and prevent them from leaking out to contaminate the body fluids. The membrane has a selective permeability to small substrate molecules and can also release products to the environment. Copyright © 2005, Nature Publishing Group.
The first and most common successful attempt was to use these semipermeable microcapsules to encapsulate activated charcoal for treating poisoning, relieving symptoms in uremic patients and removing toxic molecules in patients with liver failure [96,100,101]. Although naked charcoal has been used in the clinic for detoxification [102], it has some inherent problems such as a sustained release of fragments in vivo often causing an embolism. This problem can be resolved by encapsulating the charcoal in microcapsules. Since the early 1980s, this approach has been used routinely worldwide for the treatment of acute poisoning. There are also other exciting, successful attempts for enzyme therapy and cell therapy that has been reviewed extensively elsewhere [90–94].
Inorganic membrane
Recently, attempts have been made to use inorganic colloidosomes as micro-compartments to construct artificial cells [103]. These colloidosomes are mainly synthesized from spontaneous self-assembly of colloidal particles in water/oil (Pickering emulsions) biphasic systems [104–112]. They have enclosed, continuous membranes composed of a monolayer of closely packed particles. In most cases, these membranes have pores due to the interstices between particles, resulting in selective permeability. The diameters of these colloidosomes are always in several tens of micrometers, which are similar to those of cells. Furthermore, the inorganic membranes are easy to modify, providing the colloidosomes with some specific functions. Thus, these inorganic colloidosomes are very attractive for building artificial cells.
The first inorganic protocell model was exploited by Mann’s group [105]. The membrane of this inorganic protocell was described as a colloidosome capsule composed of 20–30 nm-sized hydrophobic/hydrophilic silica nanoparticle arrays, which were synthesized in the Pickering emulsion biphasic system. Water droplets containing various metabolism related biomolecules, ranging from proteins to metal ions, can be encapsulated within these silica colloidosomes during an interfacial assembly process. Specifically, it was demonstrated that within the colloidosomes, cell-free gene expression can be carried out as efficiently as in bulk aqueous solution. Also, owing to their porous membranes, these colloidsomes were used to construct microscale bio-reactors. To make these inorganic protocell models more intelligent, in a later report [113], Mann et al. grafted a pH-responsive copolymer onto the silica surface generating an electrostatically gated membrane. This protocell has a pH-dependent permeability to substrates and thus can control the enzymatic reactions. More interestingly, this inorganic protocell could also spontaneously grow and divide [114].
Subramaniam et al. [107] reported a simple method to construct a robust, semi-permeable protocell model by using the clay montmorillonite, which is a soft phyllosilicate type mineral that is abundant on the Earth. Montmorillonite is believed to be possibly involved in the process of the origin of life. For example, it is able to catalyze fatty acids to assemble into vesicles [81]. It can also help activated nucleotides to form polynucleotides [115], and protect them from ultraviolet radiation damage [116]. Subramaniam et al. described that thin layer shelled clay vesicles can be formed simply from clay-armored bubbles through a mechanical force. The formed vesicles are stable in many liquids and have submicron sized pores, which exhibit size-selective permeability. They stressed that this may facilitate the small precursors (fatty acids, nucleotides) to diffuse freely into the vesicles, while preventing the macromolecules from leaking out. These clay vesicles might provide ideal harbors for the synthesis of original biomolecules.
Non-membrane
It is known that a variety of bio-macromolecule complexes (e.g., polylysine and polyadenilic acid) can self-assemble into hydrophilic droplets in aqueous solution. This process, known as coacervation, is driven by electrostatic interactions [117]. Based on this principle, Mann’s group [118] proposed a membrane-free protocell model formed by mixing mononucleotides and cationic peptides in water (Fig. 9). These microdroplets are stable and can tolerate changes in temperature, pH, and high concentration of salt and also have a preference to take up heterocyclic molecules, such as organic dyes and porphyrins. They are also capable of sequestering some catalytic inorganic nanoparticles, proteins, and enzymes. These peptide-nucleotide droplets can be used to develop new type of primitive artificial cells, which may be considered as a plausible protocell model before the emergence of lipid-based organizations on the early Earth.
FIGURE 9.
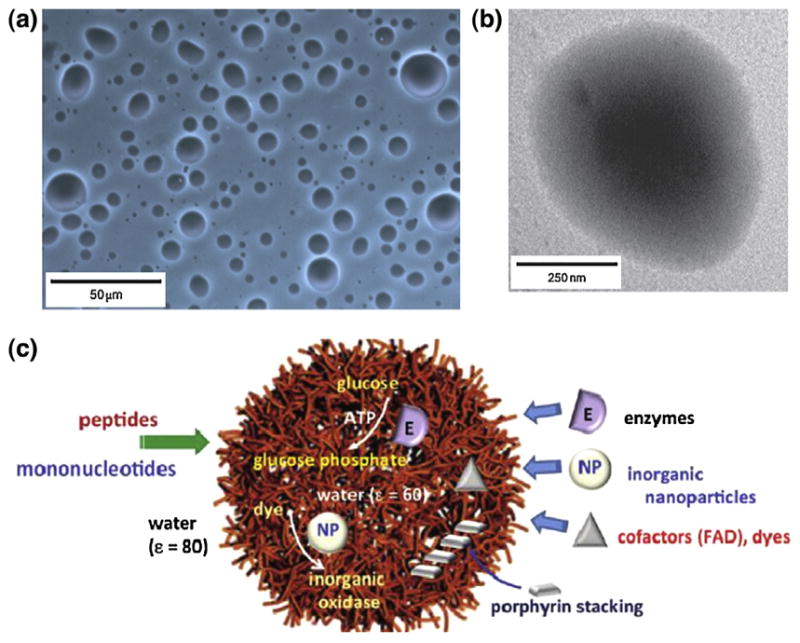
(a) Optical micrographs and (b) TEM image of polylysine–ATP droplets. (c) Scheme summarizing the properties of peptide/mononucleotide microdroplets relevant to their use as membrane-free protocell models (ε = dielectric constant). The microdroplets selectively sequester porphyrins, inorganic nanoparticles, and enzymes to generate supramolecular stacked arrays of light-harvesting molecules, nanoparticle-mediated oxidase activity, and enhanced rates of glucose phosphorylation, respectively [118]. Copyright © 2011, Nature Publishing Group.
With the merits of minimality and controllability, various types of bottom-up artificial cells have been constructed by assembling non-living components. They share some of the fundamental characteristics of natural cells and have diversified purposes ranging from a gene expression device to a bioreactor for the cell origination studies. However, the development of this field is still in its infancy. For further studies, more complex characteristics of natural cells should be engineered into these man-made cells. For example, totally self-reproducible and replicable artificial cells have not been created yet. To achieve these, the elements needed for cell replication should be able to either self-produce within the cells or be actively taken up from the environment; a driving force is also needed to regulate the replication process. Establishing intelligent communication with the outer world and with each other is also an important issue requiring an increase in the complexity of cell membranes to make them smarter. Construction of artificial cells that can perform complex cell functions ‘on demand’ is the goal. In the future, due to the flurry of studies in this field, many new benefits in biotechnology are expected.
Non-typical artificial cells (cell mimics)
As mentioned above, the non-typical artificial cells are engineered materials that simulate one or more features of biological cells and, most importantly, no restrictions on structures. They are more precisely called ‘cell mimics’ that imitate some surface characteristics, shapes, functions, or even morphology of biological cells [7].
Surface mimics
A variety of receptors, antigens, and ligands present on the surface membrane of cells perform diverse functions such as cell-cell communication. Mimicking surface characteristics, in a sense, provides an efficient way to emulate some or all of the functions of template cells. Two main methods for cell surface mimicking have been described: one is to incorporate some of the functional membrane groups into the designed particles, and the other is to coat the particles with intact cell membranes directly with or without further modifications.
Virosomes are membranous vesicles that carry viral fusion proteins in their membranes. The first reported virosome was prepared by Almeida et al. [119] by isolating the influenza virus subunits and incorporating them into lipid membranes. The resulting particles were defined as ‘virosomes’. Although whole influenza virus can be used as the immunogen, it is unsafe even after inactivation and may be pyrogenic. The highly purified influenza virus subunits, which are safer than the whole inactivated virus, are not very effective [120]. The virosomes-based artificial vaccines are efficient in inducing an immune response and do not produce any pyrogen. These artificial vaccines, therefore, represent a promising method to improve adjuvanticity. Following this work, various types of virosome-like particles containing different viral spike proteins have been generated, and the functions have been expanded broadly, such as antigen delivery, induction of cellular immune responses, DNA delivery, and gene transfer [121–123]. Although virosomes have shown promising results in various fields, safety remains a primary concern requiring further investigation before they may be used in the clinic.
Mammalian platelets are fragments of cytoplasm derived from megakaryocytes [124]. They play important roles in hemostasis [125] and immune response [126]. Their distinctive features are attributed to specific receptors expressed on the surface membrane [127]. Among these receptors, the glycoprotein (GP) Ibα, which binds to the A1 domain of multimeric von Willebrand factor (VWF-A1), makes platelets gather at the site of damaged vascular surfaces. Doshi et al. [128] modified poly(lactic-co-glycolic acid) (PLGA) nanoparticles with GPIbα/VWF-A1 to mimic natural platelets. Results showed that these nanoparticles are able to target and aggregate on the damaged vasculature.
Red blood cells (RBCs) have many attractive, beneficial features such as prolonged circulation (~120 days) and the ability to evade immune clearance. This may be due to their outstanding shapes, mechanical flexibility, as well as surface characters. The protein CD47, which is expressed on the surface of the RBC membrane, is considered an immunosuppressive protein that plays important roles in immune escape [129]. Studies showed that RBCs with CD47-knockout were taken up by macrophages very rapidly in normal mice [129]. Taking advantage of this, Tsai et al. [130] attached a recombinant domain of CD47 to IgG-opsonized poly-styrene particles. In vitro phagocytosis assays showed that CD47 can protect polystyrene particles from phagocytic clearance in a dose-dependent manner.
Zhang’s group [131–135] developed a top-down method to mimic biological cells by covering nanoparticles with cell membranes (Fig. 10). Although great progress has been made in cell surface mimics by decorating particles with cell surface groups, the chemical incorporation often causes protein denaturation. Also, these bottom-up chemical conjugation techniques are always plagued by complicated synthetic processes and the inefficacy to duplicate all of the complex properties of biological cells. The top-down method bypasses the cumbersome synthetic processes, providing a simple and efficient way to maximally retain the functional protein activities as well as replicate cell properties. Based on this principle, Zhang’s group developed a variety of cell-mimic particles such as (a) erythrocyte membrane-camouflaged nanoparticles for long-circulating drug delivery [131] (Fig. 10), (b) nanosponges to remove toxins in vivo [132], (c) cancer cell membrane-coated nanoparticles for promoting anticancer immune response and cancer-targeted drug delivery [133], (d) bacterial membrane-coated nanoparticles for developing antibacterial vaccine [134], and (e) platelet membrane-cloaking nanoparticles for improving drug localization to diseased vasculatures and against platelet-adhering pathogens [135]. Reports from other investigators also described novel approaches such as stem cell nanoghosts for targeted delivery of drugs to cancer cells [136] and leukocyte-like vectors for enhanced circulation time and improved accumulation in the tumor [137].
FIGURE 10.

Schematic of the preparation process of the RBC-membrane-coated PLGA nanoparticles (NPs). After hypotonic treatment and extrusion, the RBC membranes can be separated from the cells. These membranes are incubated with NPs. RBC membrane camouflaged NPs can be obtained following extrusion [131]. Copyright © 2011, by the National Academy of Sciences.
Shape mimics
In addition to surface characteristics, shape and mechanical properties are also crucial for cellular functions [138–140]. Particles with unique physical properties that are similar to cells have been fabricated and shown to be capable of replicating some of the functions of cells [141].
Because of the biconcave discoidal shape, RBCs have high flexibility and can squeeze through narrow capillaries with diameters smaller than the size of the cells allowing them to circulate efficiently in vivo [140]. Doshi et al. [142] synthesized polymer particles with RBC-like shape, size, and mechanical flexibility (Fig. 11a). The spherical polymeric particles used hollow polystyrene spheres or PLGA as a template. Upon solvent- or heat-induced fluidization, these particles can collapse into an RBC-like shape. To obtain flexible and biocompatible membranes, alternate layers of protein/polymer were assembled on the particle surface. After etching the particle cores, the RBC-shaped soft materials can be obtained. Similar to RBCs, these particles can pass through narrow glass capillaries with diameters smaller than that of the particles. By incorporating diagnostic and/or therapeutic agents inside, these RBC-mimics have the potential for disease diagnosis and therapy. Haghgooie et al. [143] reported the synthesis of soft PEG colloids with sizes and shapes similar to RBCs (Fig. 11b). The resulting particles have RBC-like deformability and are capable of passing through thin capillary channels. Merkel et al. [144] fabricated RBC mimetic hydrogel particles by using the particle replication through a nonwetting template (PRINT) technique (Fig. 11c). They observed that these soft particles could bypass several organs, resulting in prolonged circulation time well over those of conventional microparticles, underscoring their value for drug delivery and medical imaging.
FIGURE 11.

(a) (I) Preparation of RBC-shaped particles from hollow polystyrene (PS) templates: Proteins and polyelectrolytes are deposited onto the template surface using layer-by-layer (LBL) technique, followed by cross-linking of the layers to increase stability. PS cores are dissolved by an organic solvent to yield RBC-shaped soft materials. Proteins, drugs or contrast agents can be loaded inside the particles. (II) Preparation of biocompatible RBC-mimicking particles from PLGA templates: RBC-shaped PLGA templates are synthesized by electrohydrodynamic jetting technology. Protein layers are then coated on the template by the LBL method. After the dissolution of the template core, RBC-shaped biocompatible materials are obtained. (III) SEM image of RBC-mimicking particles prepared from PLGA template by LbL deposition of PAH/BSA and subsequent dissolution of the polymer core [142]. Copyright © 2009, by the National Academy of Sciences. (b) (I) RBC-shaped PEG particles prepared by an all polydimethylsiloxane (PDMS) microfluidic channel with a height of 4.5 mm using the stop flow lithography (SFL) technique. (II) Microscope images of RBC-shaped PEG colloids [143]. Copyright © 2009, WILEY-VCH Verlag GmbH & Co. KGaA, Weinheim. (c) RBC mimetic hydrogel particles prepared by the particle replication through the nonwetting template (PRINT) technique. (Left) Illustration for the PRINT process. The pre-hydrogel mixture (red), composed of 2-hydroxyethyl acrylate (HEA), poly (ethylene glycol) diacrylate, a photoinitiator, and polymerizable fluorescent dyes, is implanted in the disk-shaped wells of an elastomeric fluoropolymer mold (green). The mold is peeled away at a pressured nip (black), wicking away the excess liquid. Then the filled mold is exposed to UV light to irradiate the cross-linking of the hydrogel. After removing the mold by freezing onto a thin film of 1% poly (vinyl alcohol) in water (blue), the RBC-shaped hydrogel particles are obtained. (Right) Fluorescent images of (Top) 10% cross-linked, (Bottom) 2% cross-linked RBC shaped hydrogel. Scale bars are 20 μm [144]. Copyright © 2011, by the National Academy of Sciences.
In nature, viruses occur in an extensive variety of shapes including sphere, ellipsoid, rod, filament, bullet, brick and tadpole. These features play significant roles in physiological properties of viruses [145–147]. Studies showed that compared with spheroids, which are cleared immediately or readily taken up into cells, the filamentous micelles have longer circulation time [148]. Geng et al. [149] synthesized filamentous virus-mimic micelles (filomicelles) by self-assembly of amphiphilic block copolymers. These long, flexible polymer filomicelles had remarkably long blood circulation time of about one week, which is substantially longer than that of most reported spherical particles. These polymeric filomicelles could also be loaded with anticancer drug molecules and delayed tumor growth in a micelle length-dependent manner.
Morphology mimics
For many mammalian cells and organisms, their unique morphologies are critical for their functions. For example, the ultrafine morphology enables immunocytes to capture the cellular debris, abnormal cells, and foreign substances efficiently. In the past few decades, a wide range of materials with complex structures that are similar to natural organisms have been reported. However, most of them are mere shape simulation, omitting the morphology. Recently, there are several reports about constructing materials that replicate ultrafine morphological features of biological cells [150,151].
Kaehr et al. [150] developed a biomineralization method that could capture the morphology of mammalian cells precisely (Fig. 12a). The method was described as deposition of silica onto the fixed cells. Following mineralization and calcination, silica replicas that retained both the external and internal features of template cells were obtained. Using this method, Huang et al. [151] replicated the unique surface topography of macrophages by silica and magnetic nanoparticles (Fig. 12b). Cancer cells-targeting antibody was conjugated to the particles with a redox-responsive linker between them. The resulting particles exhibited improved efficiency in capturing target cancer cells from whole blood as compared with commercial magnetic beads. Also, the captured cells could be released on-demand in a redox-responsive manner.
FIGURE 12.

(a) Capturing the morphology of mammalian cells by silica: (I) Illustrations of cell silicification process, (II) Images of AsPC-1 pancreatic carcinoma cells during the silicification process, (i, ii) Images of hydrated cells, (iii, iv) Images of dehydrated composites and silica replicas, (Insets) Energy dispersive spectroscopy description of cells [150], Copyright © 2012, by the National Academy of Sciences. (b) Construction of smart mesoporous silica particles (MSPs) with cell-capturing capacity by using macrophage as a template: (I) Illustrations of the shape and morphology of macrophage by using silica and magnetic particles, (II) (i) SEM image of a pristine macrophage, (ii) TEM image of citrate-coated superparamagnetic iron oxide nanoparticles (SPIONs), (iii) TEM image of a macrophage with endocytosed SPIONs, (Inset) Higher magnification view of internalized SPIONs, Scale bar: 100 nm. (iv) SEM image of the MSP that replicated the morphology of a macrophage, (III) Illustration of modification steps of smart particles: There is a cleavable disulfide bond linker between antibody (anti-EpCAM) and the particle [151]. Copyright © 2014 WILEY-VCH Verlag GmbH & Co. KGaA, Weinheim.
Function mimics
Besides the composition and morphology mimicking, smart systems can also be constructed to mimic cell functions directly.
It is known that when a virus infects a host cell, after replicating itself and destroying the cell, it escapes from the dead cell to infect other cells. Lee et al. [152] developed a nanosized polymer vehicle that resembles the infective functions of viruses, without compromising the tumor targeting property. This virus-mimetic nanogel (VM-nanogel) consists of a pH sensitive hydrophobic polymer core, an inner polymer shell, and an outer protein shell (Fig. 13). The polymer core is rigid at high pH, but swells at lower pH. Thus, when they are taken up by cells, the pH reduction from a physiological (pH 7.4) to endosomal (pH 6.4) level induces reversible swelling of the nanogel, with a 6-fold increase in diameter. This sharp volume change in response to pH can physically disrupt endosomal membranes, allowing the nanogels to transfer from the endosomes to the cytosol, where they can rapidly shrink back to their original size. The disruption of endosome would cause cell damage and once the cells are killed and disintegrated, the nanogels could move to the neighboring cells and repeat the same cycle analogous to the infection cycle of viruses. To enhance the antitumor ability of VM, tumor-targeting ligands were conjugated to the outer protein shell and the antitumor drugs were loaded inside the polymer core. The infection ability of VM endows upon these particles a great potential for treating tumors and other diseases.
FIGURE 13.
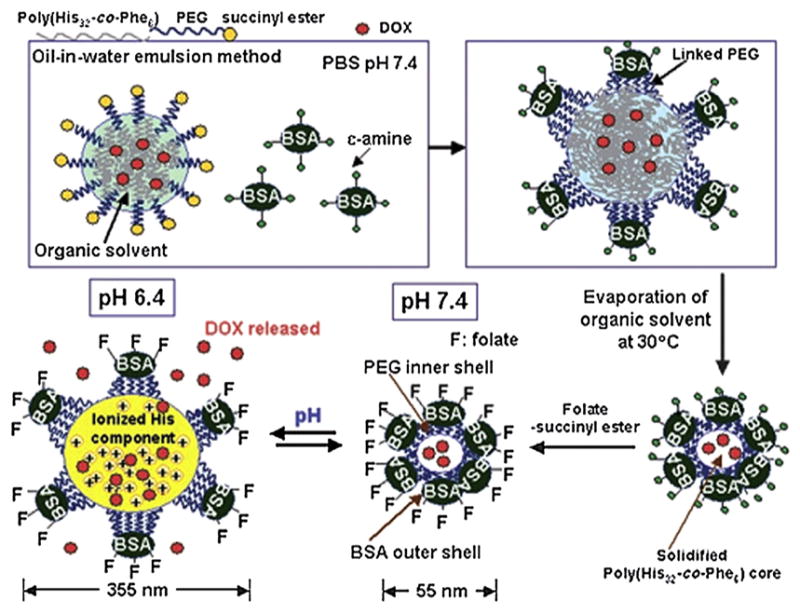
Preparation and structure of the VM-nanogel: Hydrophobic polymer, poly (l-histidine-co-phenylalanine) (poly (His32-co-Phe6)), is used to form the core of the VM-nanogel. Anticancer drug, doxorubicin (DOX) is loaded inside. PEG forms the inner shell by linking onto the polymer core. The free end of the other side of PEG is linked to BSA, resulting in a capsid-like hydrophilic protein outer shell. The resulting nanogel has a diameter of about 55 nm under pH 7.4. When the pH is decreased to 6.4, the diameter of the nanogel is spontaneously increased to 355 nm [152]. Copyright © 2008 WILEY-VCH Verlag GmbH & Co. KGaA, Weinheim.
Cell mimic particles, incorporating some beneficial traits of nature cells, have demonstrated promising advantages over traditional particles, especially in the field of biomedicine. However, the development of cell mimic particles is still in its infancy and not ready for their utility in the clinic. For biomedicine-related cell mimic particles, the physiological toxicity, pharmacokinetics, immunogenicity, and efficacy have not been well established. Also, in view of the complexity of the in vivo biological environment, the performance of these particles needs to be improved further. Nevertheless, these cell mimic particles represent an advanced bioengineering approach, which incorporates the characteristics of natural cells into non-living materials with very promising and broad biological applications potential.
Conclusion and outlook
There have been extensive efforts to construct artificial cells, which have biological cell-like structures and exhibit some of the key characteristics of living biological cells. Artificial cells can be built by a top-down approach, in which the non-essential genes are knocked out from organisms or totally replaced by synthetic ones, or by a bottom-up method, which starts from scratch by assembly of the nonliving materials. The non-typical artificial cells, which are also called ‘cell mimics’, are materials that mimic some properties of biological cells such as surface characteristics, shapes, morphology, or some functions. Despite the impressive progress to date, there is still a wide gap between artificial cells and biological cells. Many issues remain to be solved including: (i) How to make artificial cells communicate efficiently with the environment and with each other? (ii) How to construct artificial cell networks? i.e., how to make artificial cells with different functions work together? (iii) How to improve the replication, division and evolving abilities of artificial cells? And finally, (iv) how to make artificial cells uptake nutrients and move as living organisms? Answering these questions will challenge our technologies as well as our basic understanding of biological cells. Although constructing ‘totally living’ artificial cells is still farfetched, the progress toward this ultimate goal will likely present many benefits and new applications. Generally speaking, the potential benefits that artificial cells may bring include: (i) providing plausible theory for the origin of life, (ii) providing a less-interfering way to investigate and understand the cellular life, (iii) connecting the non-living to the living world, (iv) replacing engineered organisms to produce pharmaceuticals and fuels, (v) biomedical applications such as replacement or supplement of deficient cells, drug delivery or medical imaging, and (vi) adding new functions that are absent in biological cells. Undoubtedly, further development of artificial cells will bring attractive opportunities to many fields such as biotechnology, medicine, and industry.
References
- 1.Masters BR. eLS. John Wiley & Sons, Ltd; 2001. [Google Scholar]
- 2.Garrett WS. J Cell Biol. 2015;210(1):7. doi: 10.1083/jcb.201506019. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 3.Pohorille A, et al. Trends Biotechnol. 2002;20(3):123. doi: 10.1016/s0167-7799(02)01909-1. [DOI] [PubMed] [Google Scholar]
- 4.Ding Y, et al. Life. 2014;4(4):1092. doi: 10.3390/life4041092. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 5.Saraniti M. Nat Nanotechnol. 2008;3(11):647. doi: 10.1038/nnano.2008.327. [DOI] [PubMed] [Google Scholar]
- 6.Szostak JW, et al. Nature. 2001;409(6818):387. doi: 10.1038/35053176. [DOI] [PubMed] [Google Scholar]
- 7.Yoo JW, et al. Nat Rev Drug Discov. 2011;10(7):521. doi: 10.1038/nrd3499. [DOI] [PubMed] [Google Scholar]
- 8.Chiarabelli C, et al. Curr Opin Biotechnol. 2009;20(4):492. doi: 10.1016/j.copbio.2009.08.004. [DOI] [PubMed] [Google Scholar]
- 9.Malinova V, et al. FEBS Lett. 2012;586(15):2146. doi: 10.1016/j.febslet.2012.05.033. [DOI] [PubMed] [Google Scholar]
- 10.Dzieciol AJ, et al. Chem Soc Rev. 2012;41(1):79. doi: 10.1039/c1cs15211d. [DOI] [PubMed] [Google Scholar]
- 11.Sole RV. Int J Biochem Cell Biol. 2009;41(2):274. doi: 10.1016/j.biocel.2008.10.004. [DOI] [PubMed] [Google Scholar]
- 12.Luisi PL, et al. Curr Opin Colloid Interface Sci. 1999;4(1):33. [Google Scholar]
- 13.Gibson DG, et al. Science. 2010;329(5987):52. doi: 10.1126/science.1190719. [DOI] [PubMed] [Google Scholar]
- 14.Mann S. Accounts Chem Res. 2012;45(12):2131. doi: 10.1021/ar200281t. [DOI] [PubMed] [Google Scholar]
- 15.Browning ST, et al. Biotechnol Bioeng. 2001;76(3):187. doi: 10.1002/bit.10007. [DOI] [PubMed] [Google Scholar]
- 16.Fraser CM, et al. Science. 1995;270(5235):397. doi: 10.1126/science.270.5235.397. [DOI] [PubMed] [Google Scholar]
- 17.Hutchison CA, et al. Science. 1999;286(5447):2165. doi: 10.1126/science.286.5447.2165. [DOI] [PubMed] [Google Scholar]
- 18.Koonin EV, et al. Curr Opin Genet Dev. 1996;6(6):757. doi: 10.1016/s0959-437x(96)80032-3. [DOI] [PubMed] [Google Scholar]
- 19.Kolisnychenko V, et al. Genome Res. 2002;12(4):640. doi: 10.1101/gr.217202. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 20.Gil R, et al. Proc Natl Acad Sci U S A. 2002;99(7):4454. doi: 10.1073/pnas.062067299. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 21.Gil R, et al. Microbiol Mol Biol Rev. 2004;68(3):518. doi: 10.1128/MMBR.68.3.518-537.2004. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 22.Glass JI, et al. Proc Natl Acad Sci U S A. 2006;103(2):425. doi: 10.1073/pnas.0510013103. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 23.Luisi PL. Chem Biodivers. 2007;4(4):603. doi: 10.1002/cbdv.200790053. [DOI] [PubMed] [Google Scholar]
- 24.Forster AC, et al. Genome Res. 2007;17(1):1. doi: 10.1101/gr.5776007. [DOI] [PubMed] [Google Scholar]
- 25.Luisi PL, et al. Naturwissenschaften. 2006;93(1):1. doi: 10.1007/s00114-005-0056-z. [DOI] [PubMed] [Google Scholar]
- 26.Luisi PL, et al. Helv Chim Acta. 2002;85(6):1759. [Google Scholar]
- 27.Cello J, et al. Science. 2002;297(5583):1016. doi: 10.1126/science.1072266. [DOI] [PubMed] [Google Scholar]
- 28.Rasmussen S, et al. Artif Life. 2003;9(3):269. doi: 10.1162/106454603322392479. [DOI] [PubMed] [Google Scholar]
- 29.Noireaux V, et al. Proc Natl Acad Sci U S A. 2011;108(9):3473. doi: 10.1073/pnas.1017075108. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 30.Nandagopal N, et al. Science. 2011;333(6047):1244. doi: 10.1126/science.1207084. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 31.Kushwaha M, et al. Nat Commun. 2015;6:7832. doi: 10.1038/ncomms8832. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 32.Komili S, et al. Nat Rev Genet. 2008;9(1):38. doi: 10.1038/nrg2223. [DOI] [PubMed] [Google Scholar]
- 33.Brophy JAN, et al. Nat Methods. 2014;11(5):508. doi: 10.1038/nmeth.2926. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 34.Shimizu Y, et al. Nat Biotechnol. 2001;19(8):751. doi: 10.1038/90802. [DOI] [PubMed] [Google Scholar]
- 35.Yu W, et al. J Biosci Bioeng. 2001;92(6):590. doi: 10.1263/jbb.92.590. [DOI] [PubMed] [Google Scholar]
- 36.Monnard PA, et al. Origins Life Evol Biosph. 2001;31(1–2):147. doi: 10.1023/a:1006769503968. [DOI] [PubMed] [Google Scholar]
- 37.Akbarzadeh A, et al. Nanoscale Res Lett. 2013;8 doi: 10.1186/1556-276X-8-485. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 38.Walde P, et al. Chembiochem. 2010;11(7):848. doi: 10.1002/cbic.201000010. [DOI] [PubMed] [Google Scholar]
- 39.Chakrabarti AC, et al. J Mol Evol. 1994;39(6):555. doi: 10.1007/BF00160400. [DOI] [PubMed] [Google Scholar]
- 40.Oberholzer T, et al. Chem Biol. 1995;2(10):677. doi: 10.1016/1074-5521(95)90031-4. [DOI] [PubMed] [Google Scholar]
- 41.Ellington AD. Methods in Enzymology. Vol. 224. Academic Press; 1993. p. 646. [DOI] [PubMed] [Google Scholar]
- 42.Nisbet EG. Nature. 1986;322(6076):206. [Google Scholar]
- 43.Pace NR. Cell. 1991;65(4):531. doi: 10.1016/0092-8674(91)90082-a. [DOI] [PubMed] [Google Scholar]
- 44.Krishnan M, et al. Science. 2002;298(5594):793. doi: 10.1126/science.298.5594.793. [DOI] [PubMed] [Google Scholar]
- 45.Braun D, et al. Phys Rev Lett. 2003;91(15) doi: 10.1103/PhysRevLett.91.158103. [DOI] [PubMed] [Google Scholar]
- 46.Oberholzer T, et al. Biochem Biophys Res Commun. 1999;261(2):238. doi: 10.1006/bbrc.1999.0404. [DOI] [PubMed] [Google Scholar]
- 47.Tsumoto K, et al. Langmuir. 2001;17(23):7225. [Google Scholar]
- 48.Fischer A, et al. Chembiochem. 2002;3(5):409. doi: 10.1002/1439-7633(20020503)3:5<409::AID-CBIC409>3.0.CO;2-P. [DOI] [PubMed] [Google Scholar]
- 49.Shohda K, et al. Soft Matter. 2006;2(5):402. doi: 10.1039/b516834a. [DOI] [PubMed] [Google Scholar]
- 50.Bucher P, et al. Langmuir. 1998;14(10):2712. [Google Scholar]
- 51.Wick R, et al. Chem Biol. 1996;3(2):105. doi: 10.1016/s1074-5521(96)90286-0. [DOI] [PubMed] [Google Scholar]
- 52.Menger FM, et al. Accounts Chem Res. 1998;31(12):789. [Google Scholar]
- 53.Fischer A, et al. BBA-Biomembranes. 2001;1512(2):375. [Google Scholar]
- 54.Nomura S, et al. Chembiochem. 2003;4(11):1172. doi: 10.1002/cbic.200300630. [DOI] [PubMed] [Google Scholar]
- 55.Ishikawa K, et al. FEBS Lett. 2004;576(3):387. doi: 10.1016/j.febslet.2004.09.046. [DOI] [PubMed] [Google Scholar]
- 56.Noireaux V, et al. Proc Natl Acad Sci U S A. 2004;101(51):17669. doi: 10.1073/pnas.0408236101. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 57.Kuruma Y, et al. BBA-Biomembranes. 2009;1788(2):567. doi: 10.1016/j.bbamem.2008.10.017. [DOI] [PubMed] [Google Scholar]
- 58.Gardner PM, et al. Nat Chem. 2009;1(5):377. doi: 10.1038/nchem.296. [DOI] [PubMed] [Google Scholar]
- 59.Lentini R, et al. Nat Commun. 2014;5 doi: 10.1038/ncomms5012. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 60.Schrum JP, et al. Cold Spring Harbor Perspect Biol. 2010;2(9) doi: 10.1101/cshperspect.a002212. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 61.Meierhenrich UJ, et al. Angew Chem Int Ed Engl. 2010;49(22):3738. doi: 10.1002/anie.200905465. [DOI] [PubMed] [Google Scholar]
- 62.Hargreaves WR, et al. Nature. 1977;266(5597):78. doi: 10.1038/266078a0. [DOI] [PubMed] [Google Scholar]
- 63.Deamer DW, et al. Biosystems. 1980;12(3–4):167. doi: 10.1016/0303-2647(80)90014-3. [DOI] [PubMed] [Google Scholar]
- 64.McCollom TM, et al. Origins Life Evol Biosph. 1999;29(2):153. doi: 10.1023/a:1006592502746. [DOI] [PubMed] [Google Scholar]
- 65.Rushdi AI, et al. Origins Life Evol Biosph. 2001;31(1–2):103. doi: 10.1023/a:1006702503954. [DOI] [PubMed] [Google Scholar]
- 66.Ponnampe C, et al. Nature. 1966;209(5027):979. [Google Scholar]
- 67.Astrobiology. 2005;5(2):226. [Google Scholar]
- 68.Lawless JG, et al. Nature. 1979;282(5737):396. [Google Scholar]
- 69.Komiya M, et al. Geochim Cosmochim Acta. 1993;57(4):907. doi: 10.1016/0016-7037(93)90175-v. [DOI] [PubMed] [Google Scholar]
- 70.Mautner MN, et al. Planet Space Sci. 1995;43(1–2):139. doi: 10.1016/0032-0633(94)00205-6. [DOI] [PubMed] [Google Scholar]
- 71.Gebicki JM, et al. Nature. 1973;243(5404):232. doi: 10.1038/243232a0. [DOI] [PubMed] [Google Scholar]
- 72.Hargreaves WR, et al. Biochemistry-Us. 1978;17(18):3759. doi: 10.1021/bi00611a014. [DOI] [PubMed] [Google Scholar]
- 73.Grote JR, et al. Biosystems. 1978;10(4):287. doi: 10.1016/0303-2647(78)90011-4. [DOI] [PubMed] [Google Scholar]
- 74.Cistola DP, et al. Biochemistry-Us. 1988;27(6):1881. doi: 10.1021/bi00406a013. [DOI] [PubMed] [Google Scholar]
- 75.Sacerdote MG, et al. Proc Natl Acad Sci U S A. 2005;102(17):6004. doi: 10.1073/pnas.0408440102. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 76.Mansy SS, et al. Nature. 2008;454(7200):122. doi: 10.1038/nature07018. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 77.Berclaz N, et al. J Phys Chem B. 2001;105(5):1056. [Google Scholar]
- 78.Zhu TF, et al. J Am Chem Soc. 2009;131(15):5705. doi: 10.1021/ja900919c. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 79.Oberholzer T, et al. Biochem Biophys Res Commun. 1995;207(1):250. doi: 10.1006/bbrc.1995.1180. [DOI] [PubMed] [Google Scholar]
- 80.Chen IA, et al. J Am Chem Soc. 2005;127(38):13213. doi: 10.1021/ja051784p. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 81.Hanczyc MM, et al. Science. 2003;302(5645):618. doi: 10.1126/science.1089904. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 82.Kurihara K, et al. Nat Chem. 2011;3(10):775. doi: 10.1038/nchem.1127. [DOI] [PubMed] [Google Scholar]
- 83.Adamala K, et al. Science. 2013;342(6162):1098. doi: 10.1126/science.1241888. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 84.Ferris JP, et al. Science. 1992;257(5075):1387. doi: 10.1126/science.1529338. [DOI] [PubMed] [Google Scholar]
- 85.Ferris JP, et al. J Am Chem Soc. 1993;115(26):12270. doi: 10.1021/ja00079a006. [DOI] [PubMed] [Google Scholar]
- 86.Ertem G, et al. J Am Chem Soc. 1997;119(31):7197. doi: 10.1021/ja970422h. [DOI] [PubMed] [Google Scholar]
- 87.Ogden GB, et al. Cell. 1988;54(1):127. doi: 10.1016/0092-8674(88)90186-9. [DOI] [PubMed] [Google Scholar]
- 88.Huang X, et al. Nat Commun. 2013;4 [Google Scholar]
- 89.Chang TMS. Science. 1964;146(364):524. doi: 10.1126/science.146.3643.524. [DOI] [PubMed] [Google Scholar]
- 90.Chang TMS. Nat Rev Drug Discov. 2005;4(3):221. doi: 10.1038/nrd1659. [DOI] [PubMed] [Google Scholar]
- 91.Chang TMS. Blood Purif. 2000;18(2):91. doi: 10.1159/000014430. [DOI] [PubMed] [Google Scholar]
- 92.Liu ZC, et al. Adv Exp Med Biol. 2010;670:68. doi: 10.1007/978-1-4419-5786-3_7. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 93.Chang TMS. Ann N Y Acad Sci. 1997;831:249. doi: 10.1111/j.1749-6632.1997.tb52200.x. [DOI] [PubMed] [Google Scholar]
- 94.Chang TMS. Artif Organs. 2004;28(3):265. doi: 10.1111/j.1525-1594.2004.47343.x. [DOI] [PubMed] [Google Scholar]
- 95.Orive G, et al. Nat Med. 2003;9(1):104. doi: 10.1038/nm0103-104. [DOI] [PubMed] [Google Scholar]
- 96.Chang TMS. Can J Physiol Pharm. 1969;47(12):1043. doi: 10.1139/y69-170. [DOI] [PubMed] [Google Scholar]
- 97.Chang TMS, et al. Nature. 1968;218(5138):243. doi: 10.1038/218243a0. [DOI] [PubMed] [Google Scholar]
- 98.Chang TMS. Nature. 1971;229(5280):117. doi: 10.1038/229117a0. [DOI] [PubMed] [Google Scholar]
- 99.Chang TMS. Biochem Biophys Res Commun. 1971;44(6):1531. doi: 10.1016/s0006-291x(71)80260-7. [DOI] [PubMed] [Google Scholar]
- 100.Chang TMS, et al. Ther Apher. 2000;4(2):108. doi: 10.1046/j.1526-0968.2000.004002108.x. [DOI] [PubMed] [Google Scholar]
- 101.Chang TMS, et al. J Biomed Mater Res. 1968;2(2):187. doi: 10.1002/jbm.820020202. [DOI] [PubMed] [Google Scholar]
- 102.Jones J, et al. Am J Emerg Med. 1987;5(4):305. doi: 10.1016/0735-6757(87)90358-5. [DOI] [PubMed] [Google Scholar]
- 103.Li M, et al. Curr Opin Chem Biol. 2014;22:1. doi: 10.1016/j.cbpa.2014.05.018. [DOI] [PubMed] [Google Scholar]
- 104.Dinsmore AD, et al. Science. 2002;298(5595):1006. doi: 10.1126/science.1074868. [DOI] [PubMed] [Google Scholar]
- 105.Li M, et al. Chem Sci. 2011;2(9):1739. [Google Scholar]
- 106.Binks BP, et al. Nat Mater. 2006;5(11):865. doi: 10.1038/nmat1757. [DOI] [PubMed] [Google Scholar]
- 107.Subramaniam AB, et al. Soft Matter. 2011;7(6):2600. [Google Scholar]
- 108.Wang CY, et al. Chemphyschem. 2007;8(8):1157. doi: 10.1002/cphc.200700147. [DOI] [PubMed] [Google Scholar]
- 109.He YQ, et al. ACS Appl Mater Interfaces. 2013;5(11):4843. doi: 10.1021/am400582n. [DOI] [PubMed] [Google Scholar]
- 110.Bachinger A, et al. Monatsh Chem. 2010;141(6):685. [Google Scholar]
- 111.Zhou J, et al. J Colloid Interface Sci. 2012;367:213. doi: 10.1016/j.jcis.2011.11.001. [DOI] [PubMed] [Google Scholar]
- 112.He J, et al. J Am Chem Soc. 2012;134(28):11342. doi: 10.1021/ja3032295. [DOI] [PubMed] [Google Scholar]
- 113.Li M, et al. Nat Chem. 2013;5(6):529. doi: 10.1038/nchem.1644. [DOI] [PubMed] [Google Scholar]
- 114.Li M, et al. Small. 2014;10(16):3291. doi: 10.1002/smll.201400639. [DOI] [PubMed] [Google Scholar]
- 115.Ferris JP, et al. Nature. 1996;381(6577):59. doi: 10.1038/381059a0. [DOI] [PubMed] [Google Scholar]
- 116.Hughes J, et al. BMC Evol Biol. 2007;7 doi: 10.1186/1471-2148-7-227. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 117.Oparin AI. The Origin of Life. Dover Publications, Inc; New York City: 1953. [Google Scholar]
- 118.Koga S, et al. Nat Chem. 2011;3(9):720. doi: 10.1038/nchem.1110. [DOI] [PubMed] [Google Scholar]
- 119.Almeida JD, et al. Lancet. 1975;2(7941):899. doi: 10.1016/s0140-6736(75)92130-3. [DOI] [PubMed] [Google Scholar]
- 120.Jennings R, et al. Med Microbiol Immunol. 1974;160(4):295. doi: 10.1007/BF02121445. [DOI] [PubMed] [Google Scholar]
- 121.Grgacic EVL, et al. Methods. 2006;40(1):60. doi: 10.1016/j.ymeth.2006.07.018. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 122.de Jonge J, et al. Biochem J. 2007;405:41. doi: 10.1042/BJ20061756. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 123.Daemen T, et al. Adv Drug Deliv Rev. 2005;57(3):451. doi: 10.1016/j.addr.2004.09.005. [DOI] [PubMed] [Google Scholar]
- 124.Thon JN, et al. Semin Hematol. 2010;47(3):220. doi: 10.1053/j.seminhematol.2010.03.005. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 125.Ruggeri ZM. Nat Med. 2002;8(11):1227. doi: 10.1038/nm1102-1227. [DOI] [PubMed] [Google Scholar]
- 126.Semple JW, et al. Nat Rev Immunol. 2011;11(4):264. doi: 10.1038/nri2956. [DOI] [PubMed] [Google Scholar]
- 127.Savage B, et al. Cell. 1996;84(2):289. doi: 10.1016/s0092-8674(00)80983-6. [DOI] [PubMed] [Google Scholar]
- 128.Doshi N, et al. Adv Mater. 2012;24(28):3864. doi: 10.1002/adma.201200607. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 129.Tsai RK, et al. J Cell Biol. 2008;180(5):989. doi: 10.1083/jcb.200708043. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 130.Tsai RK, et al. Blood Cell Mol Dis. 2010;45(1):67. doi: 10.1016/j.bcmd.2010.02.016. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 131.Hu CMJ, et al. Proc Natl Acad Sci U S A. 2011;108(27):10980. doi: 10.1073/pnas.1106634108. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 132.Hu CMJ, et al. Nat Nanotechnol. 2013;8(5):336. doi: 10.1038/nnano.2013.54. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 133.Fang RH, et al. Nano Lett. 2014;14(4):2181. doi: 10.1021/nl500618u. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 134.Gao WW, et al. Nano Lett. 2015;15(2):1403. doi: 10.1021/nl504798g. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 135.Hu C-MJ, et al. Nature. 2015 [Google Scholar]
- 136.Furman NET, et al. Nano Lett. 2013;13(7):3248. doi: 10.1021/nl401376w. [DOI] [PubMed] [Google Scholar]
- 137.Parodi A, et al. Nat Nanotechnol. 2013;8(1):61. doi: 10.1038/nnano.2012.212. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 138.Young KD. Microbiol Mol Biol Rev. 2006;70(3):660. doi: 10.1128/MMBR.00001-06. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 139.Frojmovic MM, et al. Physiol Rev. 1982;62(1):185. doi: 10.1152/physrev.1982.62.1.185. [DOI] [PubMed] [Google Scholar]
- 140.Guest MM, et al. Science. 1963;142(3597):1319. doi: 10.1126/science.142.3597.1319. [DOI] [PubMed] [Google Scholar]
- 141.Mitragotri S, et al. Nat Mater. 2009;8(1):15. doi: 10.1038/nmat2344. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 142.Doshi N, et al. Proc Natl Acad Sci U S A. 2009;106(51):21495. doi: 10.1073/pnas.0907127106. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 143.Haghgooie R, et al. Macromol Rapid Commun. 2010;31(2):128. doi: 10.1002/marc.200900302. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 144.Merkel TJ, et al. Proc Natl Acad Sci U S A. 2011;108(2):586. doi: 10.1073/pnas.1010013108. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 145.Geisbert TW, et al. Nat Med. 2004;10(12):S110. doi: 10.1038/nm1142. [DOI] [PubMed] [Google Scholar]
- 146.Shortridge KF, et al. Virology. 1998;252(2):331. doi: 10.1006/viro.1998.9488. [DOI] [PubMed] [Google Scholar]
- 147.Roberts PC, et al. Virology. 1998;240(1):127. doi: 10.1006/viro.1997.8916. [DOI] [PubMed] [Google Scholar]
- 148.Singh R, et al. Proc Natl Acad Sci U S A. 2006;103(9):3357. doi: 10.1073/pnas.0509009103. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 149.Geng Y, et al. Nat Nanotechnol. 2007;2(4):249. doi: 10.1038/nnano.2007.70. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 150.Kaehr B, et al. Proc Natl Acad Sci U S A. 2012;109(43):17336. doi: 10.1073/pnas.1205816109. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 151.Huang C, et al. Adv Mater. 2015;27(2):310. doi: 10.1002/adma.201402213. [DOI] [PubMed] [Google Scholar]
- 152.Lee ES, et al. Angew Chem Int Ed Engl. 2008;47(13):2418. doi: 10.1002/anie.200704121. [DOI] [PMC free article] [PubMed] [Google Scholar]