
Fig. 8. Model for RecQ and RecJ role in daughter-strand gap repair of spontaneous/endogenous DNA.
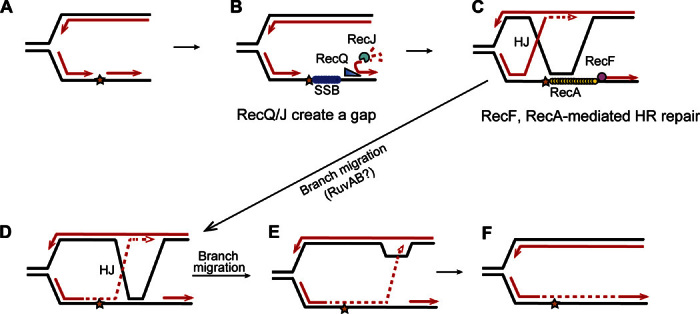
This model follows the ideas of Rupp and Howard-Flanders (95) and integrates the data (i) that most spontaneous RDG/HJ foci and intrachromosomal HR events are RecA- and RecF-dependent HR-HJs (Figs. 3, A and B, and 5B and fig. S10), reflecting RecA loading at single-strand gaps, and largely RecQ- and RecJ-dependent (Figs. 4, A to C, and 5B and fig. S10), with RecQ/J acting in the RecA-dependent pathway (Fig. 4C); (ii) that RecQ/J act before HJ formation, shown by their promotion of HJs even after RDG is produced (Fig. 4B); and (iii) that RecQ/J promote apparent RecA loading in assays of spontaneous RecA-GFP foci (Fig. 4, D and E) thought to reflect RecA-DNA nucleoprotein filaments (46). Model: During vegetative growth and DNA replication, (A) when a synthesis tract of lagging strand DNA (an Okazaki fragment, arrowheads, 3′ ends) encounters a replication-blocking lesion (star) in its template strand, (B) the adjacent Okazaki fragment can be unwound by RecQ DNA helicase, which translocates on DNA in the 3′ to 5′ direction as shown by Hishida et al. (69), exposing a 5′-ssDNA end, which is then degraded by RecJ 5′-ssDNA–dependent exonuclease. This creates a single-stranded gap, which can be coated by SSB [E. coli single-stranded DNA binding protein, like human replication protein A (96)]. (C) RecF is guided to a 5′ end of a ssDNA-dsDNA junction (43) and loads RecA onto SSB-coated ssDNA (42). The RecA-DNA nucleoprotein complex then promotes strand exchange (40, 41) so that the blocked 3′ Okazaki fragment end can displace the identical sequence (black line) in the sister chromosome, and prime continued DNA synthesis (dashed red line) using the new strand of the sister as a template. Lines, strands of DNA; red lines, new DNA strands; black lines, old strands; dashed red line, new DNA synthesis after strand exchange. (D) Branch migration of the HJ rightward, past the lesion, places newly synthesized DNA across from the lesion. Further branch rightward returns the newly synthesized strand to its original template (E) and returns the 3′ end to the original duplex (F), removing the HJ and D loop. The lesion is bypassed and, once it is part of dsDNA, can be repaired by excision repair pathways, which can act only when the damaged base or nucleotide is present in dsDNA [base excision repair and nucleotide excision repair; reviewed by Friedberg et al. (97)]. An alternative mode of resolution of the HJ is endonucleolytic cleavage, for example, by RuvC (not shown).