

Abstract
The intestinal tract is lined by a single layer of columnar epithelial cells that forms a dynamic, permeable barrier allowing for selective absorption of nutrients, while restricting access to pathogens and food-borne antigens. Precise regulation of epithelial barrier function is therefore required for maintaining mucosal homeostasis and depends, in part, on barrier-forming elements within the epithelium and a balance between pro- and anti-inflammatory factors in the mucosa. Pathologic states, such as inflammatory bowel disease, are associated with a leaky epithelial barrier, resulting in excessive exposure to microbial antigens, recruitment of leukocytes, release of soluble mediators, and ultimately mucosal damage. An inflammatory microenvironment affects epithelial barrier properties and mucosal homeostasis by altering the structure and function of epithelial intercellular junctions through direct and indirect mechanisms. We review our current understanding of complex interactions between the intestinal epithelium and immune cells, with a focus on pathologic mucosal inflammation and mechanisms of epithelial repair. We discuss leukocyte–epithelial interactions, as well as inflammatory mediators that affect the epithelial barrier and mucosal repair. Increased knowledge of communication networks between the epithelium and immune system will lead to tissue-specific strategies for treating pathologic intestinal inflammation.
Keywords: Epithelium, Mucosal Barrier, Inflammation, Mucosal Repair
The human intestinal mucosa is composed of a simple columnar epithelium, surface mucus layers, and underlying immune cell containing lamina propria (LP) and has an estimated surface area of 32 m2.1 Besides its essential role in the selective absorption of nutrients, water, and electrolytes, the intestinal epithelium provides a dynamic physical barrier that separates mucosal tissues from luminal commensal bacteria, pathogens, and dietary antigens. The intestinal epithelium, therefore, forms an important interface between the body interior and exterior. Intestinal epithelial cells (IECs) and LP immune cells coordinate the development of an adaptive response that leads to either tolerance of food antigens and the commensal flora or response against pathogenic microbes.2
Delicate and complex interactions between epithelial cells, LP immune cells, and luminal microbiota determine mucosal homeostasis. Dysregulation of these interactions results in translocation of luminal antigens, immune responses, and epithelial barrier compromise that can perpetuate pathologic mucosal inflammation. For example, compromised epithelial barrier and accompanying excessive immune responses to gut microbiota contribute to the pathogenesis of inflammatory bowel diseases (IBDs).3 However, dysregulated barrier function is not the only factor that contributes to these types of disorders. Healthy relatives of patients with Crohn’s disease (CD) have increased intestinal epithelial permeability without disease, indicating that additional factors are required for pathogenesis.4,5 Despite these observations, disease severity correlates with mucosal recruitment of polymorphonuclear neutrophils (PMNs) that traverse the epithelium and disrupt barrier function. Furthermore, recruitment of leukocytes into the subepithelial space creates a local milieu enriched in soluble pro- and anti-inflammatory mediators that impair epithelial barrier function, which can perpetuate injury (inflammatory flares) while also promoting healing.6 We review mechanisms of epithelial barrier compromise during mucosal inflammation.
The mucus layer and luminal microbiota have important roles in regulating intestinal mucosal homeostasis and contribute to pathogenesis.7–9 Here we discuss the evidence supporting direct and indirect contributions of immune cells and inflammatory mediators to regulation of epithelial barrier function, emphasizing key structural components of the intestinal barrier, specifically, the intercellular junction molecular complexes.
Intercellular Junctions
The intestinal epithelium is dynamically renewed within a week. It is truly remarkable that barrier properties are not only maintained but modified as crypt-progenitor stem cells proliferate, differentiate, and are shed at the luminal surface (Figure 1A).10 Intestinal epithelial stem cells generate multiple cell lineages, including absorptive enterocytes (80% of the cells), mucus-producing goblet cells, enteroendocrine cells, and antimicrobial peptide-producing Paneth cells. Microfold cells are found over mucosal lymphoid aggregates and mediate transcytosis of antigens across the epithelium. Intra-epithelial lymphocytes are found between IECs and have roles in immune regulation. IECs express pattern-recognition receptors that bind to highly conserved structures on commensal bacteria and pathogens. Pattern-recognition receptors activate intracellular signaling pathways that affect epithelial responses, such as generation of protein and lipid mediators, which are released into the epithelial milieu to regulate homeostasis.11,12 The contribution of epithelial cells to maintenance of immune tolerance in the gut has been a focus of a number of reviews and will not be discussed here.13
Figure 1.

Structure of the intestinal mucosa and intercellular junction complexes. (A) The intestinal mucosal barrier is composed of a monolayer of columnar epithelial cells that is separated from luminal contents by mucus and is in close contact with a lamina propria containing immune cells, capillaries, and lymphatic vessels. In the small intestine, the epithelium is folded to form luminal structures termed villi and crypts, while the colon lacks villi and only has crypts. The epithelium is continuously renewed with progenitor stem cells in the crypt base that proliferate, differentiate, and migrate along the crypt–luminal axis followed by regulated cell death at the luminal surface. The epithelium includes absorptive enterocytes, mucus-producing goblet cells, enteroendocrine cells, and antimicrobial peptides-producing Paneth cells. AMPs, antimicrobial peptides; IEL, intraepithelial lymphocyte. (B) Epithelial intercellular junctional complexes consist of an apical junctional complex (comprising TJs and subjacent AJs) followed by DMs. The TJ and AJ is associated with an underlying perijunctional actin-myosin cytoskeleton, while DM associates with intermediate filaments. TJ, AJ, and DM complexes include transmembrane proteins that associate with TJ-associated scaffold and signaling molecules as indicated. aPKC, atypical protein kinase C; CLMP, CAR-like membrane protein; MAGI-1, membrane-associated guanylate kinase; Par-3/Par-6, partitioning defective 3/-6.
Epithelial barrier function is mediated by a series of intercellular junctions that include an apical tight junction (TJ), subjacent adherens junction (AJ), and desmosomes. Given their close structural and functional proximity, the TJ and AJ are collectively referred to as the apical junctional complex (AJC). The backbone of intercellular junctions consists of transmembrane proteins that associate with cytoplasmic plaque proteins anchored to the cytoskeleton (Figure 1B).
Tight Junctions
TJs reside at the interface between the apical and basolateral plasma membrane domains. In addition to controlling the movement of fluids and solutes in the paracellular space, TJs contribute to the establishment of cell polarity.14,15 By electron microscopy, TJs are visualized as areas of close plasma membrane appositions between adjacent cells. Freeze-fracture electron microscopy has identified anastomosing multi-strand structures at sites of intercellular contact that vary in different epithelia and endothelia.15 The complexity of TJ strands correlates positively with the tightness of the paracellular barrier. Three main families of transmembrane proteins constitute the TJ backbone: claudins, TJ-associated MARVEL domain-containing proteins, and members of the cortical thymocyte marker in Xenopus family, such as junctional adhesion molecules (JAMs).
Epithelial paracellular permeability is determined by a high-capacity, charge- and size-selective pore pathway that is permeable for ions and small solutes with a molecular radius of <4 Å and a low-capacity, charge- and size-independent leak pathway that is involved in the paracellular trafficking of macromolecules.16,17 The pore and leak pathways are mediated by specific molecular constituents of the TJ.
Claudins
Claudins are required for TJ function and are part of a large family that contains 27 tetraspan integral membrane proteins in humans. They consist of 2 extracellular loop domains with cytoplasmic N- and C-terminal domains. Claudins oligomerize in cis and trans, offering diverse combinations and the complement of claudins in TJ strands influences cellular barrier function.18,19 Claudins regulate the TJ pore pathway by forming either anion-selective or cation-selective paracellular channels. Based on effects on epithelial permeability, they have been grouped into tight claudins (1, 3, 4, 5, and 18), which increase barrier tightness, and leaky claudins (2, 10, and 15), which contribute to increased paracellular permeability.20 A recent report on the crystal structure of claudin 15 has shed light on the molecular details of claudin dimerization and ion binding.21 Studies are needed to resolve higher-order structures of different claudin oligomers within TJ strands that control paracellular permeability.
Claudins are expressed differentially along the length of the gastrointestinal tract as well as within the crypt-luminal axis. Claudins 2, 10, 13, and 15 are most frequently expressed in the crypt base, whereas claudins 3, 4, and 7 are expressed at the luminal surface22,23 (Table 1). This topographic separation of intestinal claudins is perturbed during mucosal inflammation and has been proposed to contribute to barrier disruption in the inflamed gut. In addition to enrichment in TJs, some claudins (1, 3, 4, 5, and 7) localize in the lateral plasma membrane of differentiated enterocytes. Although little is known about the function of nonjunctional claudins, it is likely that lateral-membrane pools of claudins are essential for signaling events that control epithelial homeostasis. The intracellular C-terminal domain of claudins binds to TJ-associated PSD95–DlgA–ZO1 homology (PDZ)–containing proteins, such as zonula occludens (ZO) proteins and multi-PDZ domain protein 1 (MPDZ or MUPP1). These interactions have been proposed to contribute to recruitment of scaffold and signaling proteins to TJs.24 Several recent studies using knockout mouse models have begun to dissect the roles of individual claudin isoforms. For example, deletion of claudin 7 from intestinal epithelial cells in mice increased the paracellular flux of small molecules, such as the bacteria-derived N-formyl-L-methionyl-L-leucyl-L-phenylalanine peptides (438 Da), resulting in mucosal inflammation.25,26 In agreement with several in vitro studies, knockout of leaky claudin-2 in mice decreased paracellular cation permeability in the small intestine.27,28 Claudin 2 knockout mice (Cldn2−/−) have exaggerated intestinal mucosal inflammation and susceptibility to colitis, whereas mice that overexpress a Cldn2 transgene are protected from experimental colitis, despite increased intestinal mucosal permeability.28,29 The apparent dissociation between the effects of claudin 2 on the mucosal barrier and those on intestinal inflammation indicates that claudin 2 regulates intestinal epithelial homeostasis by other, undetermined mechanisms.30 This concept is supported by the observation that claudin 15-knockout mice develop mega-intestine due to greatly increased intestinal epithelial cell proliferation.31 Double loss of claudins 2 and 15 is associated with defective nutrient absorption and lethality shortly after birth.32
Table 1.
Roles and Location of the Main Claudins of the Gastrointestinal Tract
| Claudins | Expression
|
Function
|
|||
|---|---|---|---|---|---|
| Stomach | Small intestine | Colon | Barrier-forming | Pore-forming | |
| Cld-1 (TJ+basolateral membrane) | ND | Crypt+villus | Crypt | + | NA |
| Cld-2 (TJ) | NA | Crypt base | Crypt base | NA | Cations (Na+) water |
| Cld-3 (TJ+basolateral membrane) | Surface | Crypt: low Villus: high |
Crypt base: low Crypt surface: high |
+ | NA |
| Cld-4 (TJ+basolateral membrane) | Gastric gland | Villus apex | Crypt surface | + | Anions (Cl−) when associated with Cld-8 |
| Cld-5 (TJ+basolateral membrane) | Uniformly | Crypt+villus | Crypt base | + | NA |
| Cld-7 (TJ+basolateral membrane) | NA | Crypt: low Villus: high |
Crypt base: low Crypt surface: high |
NA | Cations (Na+) |
| Cld-8 (TJ) | NA | Crypt+villus | Mid-crypt | + | Anions (Cl−) |
| Cld-10 (TJ) | NA | Crypt: high Villus: low |
Crypt base | NA | Cations |
| Cld-12 (TJ) | ND | Crypt+villus | Crypt | + | Calcium |
| Cld-13 (TJ) | NA | NA | Crypt | + | NA |
| Cld-15 (TJ) | NA | Crypt: high Villus: low |
Crypt base | NA | Cations (Na+) |
| Cld-18 (TJ) | Uniformly | ND | ND | + | Cations (Na+, H+) |
| Cld-23 (TJ) | ND | ND | ND | ND | ND |
Pathogenic bacteria of the gastrointestinal tract have been reported to target specific claudins, causing TJ disassembly and barrier defects. Clostridium perfringens enterotoxin (CPE), responsible for food poisoning in humans, was initially shown to bind claudins 3 and 4, inducing internalization of those claudins from the TJ and compromising barrier function.33,34 Additionally, claudins 6, 7, 8, 9, and 14 have been shown to associate with CPE.35 Most recently, the crystal structure of the claudin 19–CPE complex identified the amino acids that mediate the interactions between CPE and specific claudins these observations provided insight into the mechanisms by which CPE disrupts cis and trans claudin interactions.36 Claudin-targeting CPE, or its derivative peptides, could reversibly open TJs and be used in transepithelial delivery of drugs. Analogous to CPE, Helicobacter pylori can induce the loss of claudins 4 and 5 from gastric epithelial TJs.37 In addition to interacting with bacteria, claudins can influence viral protein entry into cells. For example, claudin 1 functions as a co-receptor for hepatitis C virus.38
Tight Junction-Associated MARVEL Domain-Containing Proteins
Members of this tetraspan family of proteins include occludin, MARVELD2 (found at tricellular contacts), and MARVELD3.39 Occludin was the first identified TJ transmembrane protein, but there is controversy over its role in controlling epithelial barrier function. Well-developed TJ strands were reported in occludin-deficient epithelial cells.40,41 Occludin-deficient mice are viable, exhibiting normal intestinal epithelial-barrier function.42,43 Synthetic peptides mimicking the first or second extracellular domain of occludin attenuate TJ assembly. However, other studies have reported that occludin expression can influence paracellular flux, which requires its C-terminal cytoplasmic domain.44–50
Analogous to occludin, co-expression of MARVEL (membrane-associating) domain containing 3 (MARVELD3) with claudin 1 increases the number of TJ freeze-fracture strands.51 MARVELD3 has been shown to affect cell behavior and survival by coupling TJs to MEKK1–JNK signaling.52 The tetraspan TJ-associated MARVEL domain-containing proteins family members MARVELD2 and lipolysis-stimulated lipoprotein receptor containing an immunoglobulin-like domain, localize to tricellular TJs, and contribute to barrier regulation at these sites.53–55 TJ-associated MARVEL domain-containing proteins interact with intestinal pathogens that disrupt the intestinal epithelial barrier. For example, Shigella targets MARVELD2 and spreads throughout the intestinal mucosa via tricellular TJs.56 N-3-(oxododecanoyl)-homoserine lactone, a quorum-sensing molecule produced by intestinal Gram-negative bacteria, influences TJ barrier function by down-regulating MARVELD2 and occludin expression.57 Furthermore, lipolysis-stimulated lipoprotein receptor serves as an epithelial receptor for Clostridium difficile transferase and Clostridium perfringens iota toxin.58
Cortical Thymocyte Marker in Xenopus Family Proteins
The classical cortical thymocyte marker in the Xenopus family of adhesion molecules includes F11 receptor (JAM-A), JAM-B, and JAM-C. JAM proteins share a single transmembrane domain with 2 extracellular immunoglobulin-like loops (V-C2-type Ig-domains) and a cytoplasmic tail (40–50 residues) containing C-terminus class II PDZ-binding motif.59 Among the classical JAM proteins, JAM-A is expressed in IECs and has been implicated in several aspects of epithelial homeostasis, including barrier function, cell proliferation, and migration.60 Based on crystallographic structural studies, JAM-A is reported to form homodimers both in cis and in trans.61,62 JAM-A has an important role in the regulation of epithelial barrier function. Knockout mice and JAM-A–deficient human epithelial cells have barrier defects that are associated with increased expression of leaky claudins (claudins 10 and 15) and with alterations in the peri-junctional actin cytoskeleton.63–65 JAM-A–deficient mice do not have symptoms of intestinal disease because of mucosal compensatory and protective adaptive immune responses that are secondary to a leaky intestinal epithelial barrier.65 However, they have increased susceptibility to colitis, supporting a role for JAM-A in regulation of intestinal epithelial barrier function and mucosal homeostasis.64
JAM-A can interact through its C-terminal PDZ-binding motif with several TJ scaffold proteins, such as ZO proteins, par-3 family cell polarity regulator (PARD3/Par3), afadin, and MUPP1.59 Those molecular interactions mediate JAM-A regulation of TJ assembly, epithelial homeostasis, and barrier function. Furthermore, by associating with PARD3 and the polarity complex (Par3, PARD6A/Par6, and aPKC), JAM-A has been implicated in controlling cell polarity.66 It has been shown that JAMA forms a signaling complex through association with ZO-2, afadin, and PDZ-GEF1 to control activation of RAP2C, which in turn regulates epithelial permeability to high-molecular-weight solutes by modulating levels of RhoA and apical actomyosin cytoskeletal contraction.67 Furthermore, reovirus surface protein σ1 binds to the extracellular domain of JAM-A, resulting in the loss of TJ-associated JAM-A and increased permeability.67,68
Coxsackie and adenovirus receptor (CAR) is another cortical thymocyte marker in Xenopus family member that is closely related to the JAM proteins. CAR possesses a single transmembrane domain with 2 extracellular immunoglobulin-like loops (V-C2 type Ig domains) and a cytoplasmic tail (107 residues) containing a class I PDZ-binding motif at the C-terminus.69 CAR localizes in TJs, where it physically interacts with ZO-1 and MUPP1 to regulate epithelial barrier function.69 Loss of CAR does not affect the formation of the TJ protein complex in CARdeficient mice.70 However, CAR has been reported to control AJ stability by regulating E-cadherin (CDH1) at the cell surface, which is mediated by protein kinase C δ-dependent phosphorylation of CAR.71
Tight Junction Plaque and Signaling Proteins
More than 50 proteins have been identified in TJs, and the list is growing. PDZ domain-containing proteins, actin-binding proteins, signaling complexes (such as kinases, phosphatases, and small GTPases) associate with TJ transmembrane proteins and contribute to the formation of the TJ plaque by recruiting adapter proteins, signaling molecules, and transcriptional regulators.72,73 Among the PDZ domain-containing proteins, the membrane-associated guanylate kinase family of proteins ZO-1, ZO-2, and ZO-3 interact directly with the C-terminal PDZ-binding motif of claudins, JAM-A, and CAR.74 In contrast, the C-terminus domain of occludin binds to the guanylate kinase and acidic domains of ZO-1. Additionally, the ZO proteins serve as a link between TJ transmembrane proteins and the actomyosin cytoskeleton, with important roles in control of TJ function.75
In addition to ZO proteins, other PDZ domain-containing proteins in the TJ include scaffold proteins (such as afadin or ALL1-fused gene from chromosome 6 protein),76 MUPP1,77 MAGI1,78 polarity complex proteins (Par3, Par6, scribble),79 and signaling proteins, including kinases (eg, atypical protein kinase C), phosphatases (PP2A, PTEN),80 and guanine nucleotide exchange factors that activate small GTPases (PDZ-GEFs67). Additionally, proteins that lack PDZ domains, such as cingulin bind to JAM-A, occludin, ZO-1, and the actin cytoskeleton.81–84 Small GTPases (RAP2, Rho, and Rac), kinases such as myosin light chain kinase (MLCK), and phosphatases (eg, PP2A, PTPN285), are recruited to the TJ via the scaffolding proteins and regulate peri-junctional actomyosin dynamics and barrier function.86,87
Adherens Junction and Desmosomes
The assembly and maintenance of the intestinal epithelial barrier is not limited to TJs, but depends on additive effects and crosstalk among TJs, AJs, and desmosomes.88–90 The AJ is an ancient junctional complex that initiates and maintains epithelial cell–cell contacts. The key transmembrane protein in the epithelial AJ is E-cadherin (CDH1), which mediates calcium-dependent homotypic intercellular adhesions. On the cytoplasmic face of the AJ, E-cadherin associates with p120 (CTNND1), β-catenin (CTNNB1), and α-catenin (CTNNA1), forming a complex that is anchored to cortical actin filaments.91 An early study involving transgenic mice expressing a dominant-negative form of N-cadherin in the intestinal epithelium reported disruption of the mucosal barrier and the development of CD-like intestinal inflammation.92 In addition, studies involving conditional knockout mice with intestinal epithelial-specific deletion of E-cadherin93 or p12094 resulted in disruption of epithelial architecture and spontaneous mucosal inflammation.
A critical common mechanism that regulates the assembly and stability of AJ and TJ involves interactions with the cortical actin cytoskeleton. AJ and TJ cytosolic plaques contain a number of actin-binding proteins (α-catenin, vinculin, ZO family, afadin, and cingulin) that anchor junctional complexes to underlying F-actin bundles.95,96 Studies that used either pharmacologic or genetic tools to disrupt actin filaments demonstrated an important role of the actin cytoskeleton in controlling AJ and TJ function.96,97 A key mechanism that regulates peri-junctional actin cytoskeleton dynamics and AJ/TJ biogenesis involves the activity of nonmuscle myosin II (NM II) motor protein. NM II stabilizes cytoskeletal structures by bundling actin filaments and mediates cytoskeletal rearrangement by generating forces for filament movement.98 Both inhibition and activation of NM II disrupts the epithelial barrier and triggers AJ and TJ disassembly.99–101 Likewise, recent studies using knockout and transgenic mouse models revealed that both the inhibition of the actin motor, via the intestinal epithelial-specific knockout of NM IIA,102 and overactivation of NM II, via the overexpression of constitutively active MLCK in the intestinal epithelium,103 resulted in increased intestinal permeability. These findings suggest that balanced NM II activity controls integrity of the gut barrier.
Desmosomes (DMs) provide mechanical strength to the epithelium. DM transmembrane cadherins include desmoglein and desmocollin proteins. Among the 7 DM cadherins (4 desmogleins and 3 desmocollins), human IECs express only desmoglein 2 and desmocollin 2, which associate with plaque proteins, including plakophilin and plakoglobin, which in turn bind to the desmoplakin that anchors the DM protein complex to intermediate filaments.104 Although little is known about the role of DM proteins in controlling the intestinal epithelial barrier, important regulatory roles for the adhesive structures of the DM are easy to envision because the intestinal epithelium is under substantial mechanical stress (stretching and compression) generated by peristaltic movement of the gut.
Effects of Inflammation
Mucosal inflammatory diseases are typically associated with compromised epithelial barrier function. It is generally believed that the trafficking of immune cells in concert with the release of different mediators represent key mechanisms of barrier disruption in the inflamed intestinal mucosa. Furthermore, it is increasingly appreciated that the luminal microflora promotes mucosal inflammatory responses, especially under conditions of altered epithelial permeability (recently reviewed in Donaldson et al105). We summarize the direct effects of leukocytes, as well as the indirect effects of mediators released by leukocytes and epithelial cells, on the epithelial barrier and homeostasis.
Direct Effects of Leukocytes
Leukocyte transepithelial migration
PMNs are the first responders to infection or mucosal injury, responding to transepithelial gradients of microbial and cell-derived chemoattractants. In addition to microbe-derived chemoattractants, such as formylated peptides, the engagement of epithelial pattern-recognition receptors by luminal antigens results in the release of inflammatory cytokines and chemokines, such as interleukin (IL)1, IL6, IL8, tumor necrosis factor (TNF), interferon (IFN) gamma, and CCL2, leading to the recruitment and activation of leukocytes. Neutrophils, for example, emigrate from blood vessels, reach the sub-epithelial space, and migrate across the epithelial monolayer toward the intestinal lumen.
Although the transendothelial migration of leukocytes has been studied extensively,106 many details of the molecular mechanisms that promote neutrophil transepithelial migration (TEpM) are unclear. Lack of understanding of the molecular details of TEpM is surprising, because disease symptoms and pathologic epithelial permeability are closely correlated to this event. In vitro approaches, involving Transwell inserts with polarized epithelial monolayers cultured on permeable filters, have been used to characterize interactions between PMN and epithelia.107 Epithelial monolayers cultured on the underside of Transwell filters have been used to model PMN-mediated TEpM in a physiologically relevant basolateral to apical direction. These studies have increased our understanding of the effects of PMN–epithelial interactions on barrier function and wound healing (Figure 2A). Monoclonal antibody–based approaches, along with Transwell-based PMN transmigration assays, have been used to study events that control PMN TEpM. In vitro approaches and complementary in vivo observations have confirmed that large-scale PMN TEpM results in the disruption of barrier function through the creation of microdiscontinuities in the epithelium108,109 (Figure 2B). There are circumstances, however, where TEpM does not result in barrier compromise, which would suggest that, under certain conditions, PMN-epithelial signaling events might actually preserve barrier function.110
Figure 2.
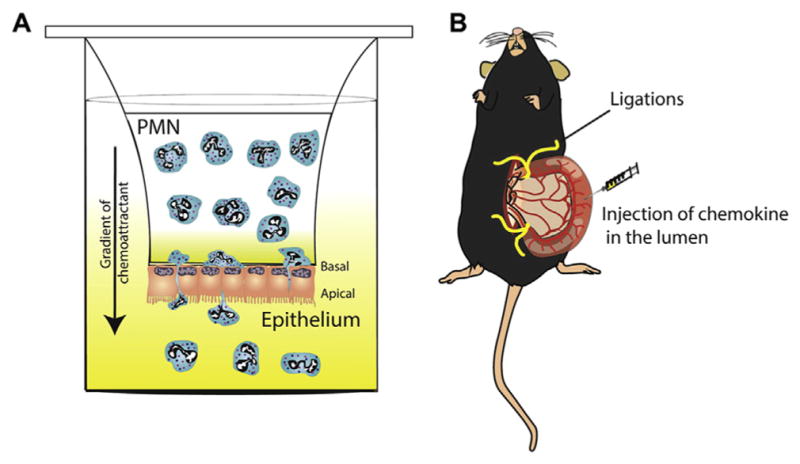
In vitro and in vivo models of neutrophil trans-epithelial migration. (A) Schematic of an in vitro model of PMN TEpM across a monolayer of intestinal epithelial cells in response to a chemoattractant (such as N-formyl-L-methionyl-L-leucyl-L-phenylalanine peptides, leukotriene B4 or CXCL1). Epithelial cells are grown on collagen-coated Transwell filters with pore sizes sufficient for PMNs to migrate through and in an inverted conformation designed to model the physiologically relevant polarity of migration, from basolateral to apical. Transmigrated PMN are collected in the bottom of the well and quantified. (B) In vivo “ileal loop” model of PMN TEpM. A vascularized 2–4 cm long ileal portion of the small intestine near the ileo–cecal junction is exteriorized and ligated at both ends without compromising the blood supply. Chemoattractant is injected into the lumen to induce PMN migration. The loop content is collected and PMN quantified by flow cytometry, while ileal loop mucosa can be analyzed histologically.
Current TEpM studies highlight a multistep process involving adhesive interactions between surface proteins (receptors) on IECs and counter-receptors on neutrophils (Figure 3). Sequential steps encompass initial adhesion to the basal membrane, migration between cells, and subsequent interactions of the post-migrated PMNs with the apical epithelial surface. Compared with leukocyte diapedesis across endothelia, which can be transcellular (through the cell body), or paracellular (between the cells), neutrophil TEpM occurs only via the paracellular route in a basolateral-to-apical direction. To migrate through the paracellular space, PMNs must sequentially cross DM, AJ, and TJs before reaching the intestinal lumen.
Figure 3.

Multistep model of neutrophil transepithelial migration across intestinal mucosa. Migration of neutrophils out of blood vessels and across the epithelium during mucosal inflammation is a multistep process involving many surface adhesion molecules on the epithelium and neutrophils: (1) basolateral adhesion, (2) transmigration, and (3) apical adhesion. In addition, neutrophils secrete proteases that promote cleavage of E-cadherin, desmoglein-2, and JAML. TJs contains CAR, which binds to JAML. AJs contain E-cadherin. DMs contain desmoglein-2 (Dsg-2). ADAM10, Disintegrin and metalloproteinase domain containing protein 10; AMP, adenosine monophosphate; DC, dendritic cell; ICAM-1, intercellular adhesion molecule-1; Mϕ, macrophage; MMP, matrix metalloprotease; NETs, neutrophil extracellular traps; SIRPα, signal-regulatory protein α.
The initial step in TEpM involves the adhesion of PMNs to the basal aspect of the epithelium through interactions between integrins (integrin subunit αM [ITGAM or CD11b] and integrin subunit β2 [ITGB2 or CD18]) and fucosylated ligand(s) that have not yet been identified.107,111 Although the integrin CD11b/CD18 on PMNs appears to be an important mediator of early steps in the transmigration response, there have been reports of adhesion independent of this integrin, which varies with chemoattractant signals and epithelial types.112 The rate of PMN migration along the paracellular space is regulated by the epithelial transmembrane glycoprotein CD47, which binds to leukocyte signal regulatory protein α.113,114 Unlike antibody-mediated ligation of CD11b/CD18, which results in sustained inhibition of TEpM, blockage of CD47 delays TEpM but does not diminish the total number of transmigrated PMNs.
Little is known about how PMNs migrate across inter-cellular junctions, but migration across TJs appears to involve interactions between epithelial TJ-associated CAR and JAM-like (JAML) protein on neutrophils.115 Unlike transendothelial migration, the role of JAM-A in regulating TEpM is unclear.63
Once PMNs have crossed TJs, they gain access to the apical or luminal aspect of the epithelium. Adhesive interactions of post-migrated PMNs with the apical surface have been reported and might serve to not only retain PMNs within intestinal crypts but also to elicit epithelial functional responses. PMNs have been shown to bind apically expressed epithelial molecules CD44v6 and CD55 (decay accelerating factor), which have been implicated in neutrophil detachment from the apical surface. Specific leukocyte counter receptors have not been identified. The selective expression of intercellular adhesion molecule 1 on the apical membrane of IECs binds PMN CD11b/CD18 to activate signals that increase epithelial permeability through the MLCK-dependent contraction of the peri-junctional actomyosin ring; this leads to recruitment of additional neutrophils to promote inflammation.116
Proteolytic and Non-Proteolytic Opening of Epithelial Junctions by Polymorphonuclear Neutrophil
Activated neutrophils release an arsenal of proteases, including elastase, proteinase, and matrix metalloproteinases that have potent effects on epithelial barrier function. The effects of protease release by migrating PMNs are not necessarily detrimental. There is evidence that proteolytic events have important roles in guiding TEpM in a regulated and polarized manner. During TEpM, PMN contact with the basolateral epithelial membrane results in elastase and proteinase-3–dependent cleavage and activation of the protease-activated receptors (PAR) 1 and PAR2, leading to signaling events that activate MLCK-dependent contraction of the actomyosin cytoskeleton, thereby increasing paracellular permeability.117 The end result is PAR1- and PAR2-dependent facilitation of TEpM in the physiologically relevant basolateral–apical direction, but not in the reverse direction.
The PMN serine proteinase elastase promotes cleavage of E-cadherin, thereby contributing to the disruption of AJ complexes and increasing the paracellular space.118 Although it is logical to assume that the disruption of AJ complexes would help pave the way for transmigrating PMNs, such cleavage events result in proliferative signals that are important for epithelial repair.119 There is evidence that analogous PMN-dependent proteolytic events occur at the level of the DM, which can facilitate TEpM. Matrix metalloproteinase 9 and disintegrin and metalloproteinase domain containing protein 10 induce the cleavage of the DM cadherin desmoglein 2, resulting in the disruption of DM complexes and impaired intestinal epithelial-barrier function.120
Functional Responses to Transepithelial Migration of Polymorphonuclear Neutrophil
An increasing number of studies highlight examples of inflammatory and pro-resolution signals that occur in the epithelium at various stages of PMN transmigration in response to intimate epithelial contact with PMNs. For example, although JAML mediates passage of PMN across TJs, it is also shed from activated PMNs in a zinc-containing endopeptidase-dependent manner. The shed ectodomain binds to epithelial CAR, leading to inhibition of epithelial proliferation and wound healing.121 Soluble JAML, released from migrating leukocytes, impedes epithelial restitution and promotes inflammation. That observation might be relevant to pathologic human conditions, such as active ulcerative colitis (UC), in which poorly healing colonic ulcers are associated with massive PMN infiltration and, presumably, abundant cleavage of JAML. Therapeutic strategies might be developed to target the JAML–CAR receptor–ligand interaction.
In an analogous fashion, there are examples of epithelial protective signals generated by direct interactions between PMNs and epithelial cells. Post-migrated PMNs that are in close contact with the apical epithelial surface release 5′ adenosine monophosphate, which is converted to adenosine through the action of an apically expressed ectonucleotidase CD73. Adenosine is a potent inducer of electrogenic chloride secretion that causes diarrhea and might represent a purging response to flush the intestine of noxious agents and microbes.122,123 Post-migrated PMNs also bind epithelial intercellular adhesion molecule 1, resulting in pro-restitutive signals. The engagement of epithelial intercellular adhesion molecule 1 enhances intestinal wound healing through the activation of Akt signaling via β-catenin.116 That observation is analogous to proliferative responses observed in the lung after PMN elastase-mediated cleavage of E-cadherin, which require β-catenin, during TEpM.119 Neutrophil TEpM therefore has temporal effects on epithelial function, which increases further leukocyte recruitment and stimulates reparative responses.
Indirect effects of inflammation on the epithelial barrier, mediated by inflammatory cytokines
Epithelial barrier defects are a common manifestation of different gastrointestinal disorders, including IBD and celiac disease. Correlation of a leaky gut epithelium with liver pathogenesis and systemic disorders, such as diabetes and allergies, have been proposed but are not yet well understood. In recent years, there has been an increased appreciation of molecular mechanisms regulating disruption of the epithelial barrier in the inflamed gut and the role played by inflammatory cytokines (Figure 4).
Figure 4.

Schematic representation of the epithelial responses to pro- and anti-inflammatory cytokines. The imbalance between pro-inflammatory and anti-inflammatory cytokines is a hallmark of chronic gut inflammation, such as in inflammatory bowel disease. Overall, pro-inflammatory cytokines increase intestinal paracellular permeability, while anti-inflammatory cytokines and pro-resolving mediators stimulate epithelial repair and barrier function. Interestingly, some pro-inflammatory mediators, such as IL17A, IL23, IL36γ, and IFNγ have been reported to promote barrier protective responses (see text). Molecular mechanisms to be identified. Dsg-2, desmoglein-2.
In chronic inflammatory diseases of the intestinal mucosa, the gut epithelium is exposed to a multitude of inflammatory cytokines secreted by activated immune cells.124,125 A large complement of immune mediators has been identified in the mucosa of patients with IBD, varying with the disease type and activity. High mucosal levels of IFNγ, TNF, IL6, IL22, and IL17 have been reported in people with CD. Increased mucosal release of IL13, IL5, and IFNγ has been observed in people with UC.6 Therapeutic strategies for those disorders involve combinations of antibodies and receptor inhibitors to dampen the immune response and promote repair. Such therapeutic antibodies against cytokines include infliximab, adalimumab, certolizumab, etanercept, fontolizumab, and ustekinumab.126–132 Given the complement of cytokine changes during pathogenesis of CD and UC, it is not surprising that such therapies have been effective in select individuals only. Strategies to analyze the cytokine and chemokine profiles of patients with IBD might be developed to create combined personalized therapies.
The intestinal epithelium responds to a broad array of cytokines that influence homeostasis (proliferation, differentiation, and apoptosis) and barrier function. Inflammatory cytokines compromise epithelial barrier function by several mechanisms, including increased epithelial apoptosis.133 However, cytokines have major effects on epithelial barrier function, even in the presence of apoptosis inhibitors, supporting a role of additional mechanisms that contribute barrier compromise during inflammation.134
Decreased Expression of Junctional Proteins Under Conditions of Inflammation
Mucosal inflammation is frequently associated with the decreased expression of junctional proteins. For example, decreased expression of different molecular junctional constituents has been documented in the intestinal mucosa of patients with IBD, especially within the inflamed areas of the gut.135,136 Several studies have demonstrated that in vitro exposure of epithelial cells to IFNγ and TNF decreases protein levels of occludin, ZO-1, E-cadherin, and p120.137–140 However, other studies have not observed such cytokine-dependent effects.141 These differences are likely related to the complement and duration of cytokine exposure.
One mechanism that likely contributes to a leaky intestinal epithelial barrier in the inflamed gut is claudin switching. The mechanism involves decreased expression of several tight claudins, such as claudins 4 and 7, and the upregulation of leaky claudins, such as claudins 2 and 15. In vitro model systems have shown that TNF and IL13 increase the expression of the pore-forming TJ protein claudin 2.142,143 Additionally, IL6 and IL17 affect expression of claudin 2.144,145 Most TJs contain a number of claudin family members that have distinct compatibility profiles, which affect their ability to associate with each other and, ultimately, paracellular ion and water permeability. It is therefore not surprising that increased expression of a single claudin can affect the balance of other claudin members within TJs and thus alter paracellular permeability. Studies have been performed to better understand complex claudin dynamics in inflammation. For example, interferon gamma-induced increases in claudin 2 affect dynamics of barrier-forming claudin 4 through competitive interactions between claudins 2 and 4 for residence within the TJ.146 In an analogous fashion, loss of JAM-A in mice results in compromised intestinal epithelial barrier in parallel with increased expression of the pore-forming claudins 15 and 10.64
Trafficking of Junction Proteins
Stimulation of junctional protein endocytosis can disrupt the intestinal epithelial barrier in response to inflammatory mediators.147 IFNγ and TNF induce selective endocytosis of the TJ proteins occludin, claudin 1, claudin 4, and JAM-A in the intestinal epithelium, in vitro and in vivo.148,149 More robust stimuli, such as oxidative stress and extracellular calcium depletion, induce AJ disassembly and E-cadherin internalization.150,151 The types of endocytic pathways that are involved in TJ protein internalization in the inflamed intestinal epithelium depend on the inciting stimulus as well as cell type. IFNγ for example, mediates the endocytosis of TJ transmembrane proteins by macropinicytosis, whereas TNFα promotes endocytosis of occludin by caveolar-mediated pathways.148,149,152
It should be noted that activation of the endocytic molecular machinery alone is not sufficient to induce junctional disassembly, which is facilitated by rearrangements of the perijunctional actin cytoskeleton.97 Such cytoskeletal rearrangements can be induced by increased F-actin turnover and NM II activity.100 NM II activation is especially important for cytokine-induced AJC disassembly and mediates IFNγ–induced macropinocytosis and the TNF-induced caveolar endocytosis of TJ proteins.149,153 Activation of NM II in intestinal epithelial cells exposed to cytokines involves phosphorylation of regulatory myosin light chain (RMLC), mediated by activation of RhoA GTPase and the downstream Rho-associated kinase, which inhibits myosin light chain phosphatase activity.154 The other pathway involves activation of MLCK, which directly phosphorylates Ser18 and Thr19 of RMLC.155 Activation of RMLC signaling has been observed in response to exposure of epithelial cells to cytokines, including IFNγ, TNF, IL13, and IL1B; this results in AJC remodeling and compromised barrier function.156
In vivo models of intestinal inflammation have verified findings from in vitro studies of the RMLC, emphasizing the importance of those regulatory pathways in the control of actin-myosin dynamics and barrier function during inflammation. Consistent with the requirement for balanced NM II activity in the maintenance of a normal intestinal epithelial barrier, barrier integrity also requires a balance of Rho signaling via Rho-associated kinase. Activation and the inhibition of Rho GTPase induces TJ disassembly and IEC barrier disruption.157,158
Trans interactions between integral membrane AJ and TJ proteins inhibit junctional endocytosis.159 The disruption of those interactions would therefore be expected to accelerate the disassembly and internalization of junctional complexes. In the inflamed gut, this is likely to be accelerated by the protease-mediated cleavage of adhesive AJ and DM proteins.160 For example, IL1B and TNF promote the cleavage of the extracellular domains of intestinal epithelial desmoglein 2, desmocollin 2, and E-cadherin in a disintegrin and metalloproteinase domain containing protein 10 and matrix metalloproteinase 9–dependent manner.120 Such cadherin-cleaved ectodomains have signaling properties that influence epithelial homeostasis. In addition to perturbing intercellular adhesion, shed desmoglein 2 and E-cadherin ectodomains have been shown to promote epithelial proliferation by activation of HER2 and HER3 signaling. Cadherin ectodomain fragments have been detected in the inflamed intestinal mucosa of mice with colitis, as well as in the mucosa of people with UC, suggesting potential utility as biomarkers of active inflammation.161
A much less appreciated mechanism that might contribute to intestinal barrier disruption in the inflamed gut involves attenuated trafficking of proteins to the AJC. Because components of epithelial junctions are synthesized in the endoplasmic reticulum and delivered to the plasma membrane via multistep, vesicle-mediated exocytosis,91 it is reasonable to assume that defects in the trafficking machinery might contribute to epithelial barrier compromise during inflammation. Reticulon 4 (RTN4 or NOGO-B) is a structural endoplasmic reticulum protein that affects protein movement from the endoplasmic reticulum to the Golgi, vesicle formation, and trafficking of proteins such as E-cadherin and α-catenin. Decreased reticulon 4 expression has been observed in the intestinal mucosa of people with CD and in IL10-knockout mice with spontaneous colitis.162 Another study highlighted the role of a polarity protein Par3 in the regulation of TJ protein occludin trafficking and barrier function that was disrupted after exposure to inflammatory cytokines.163
Inflammation and Restoration of Mucosal Barrier Function
Although the deleterious effects of neutrophils on tissue homeostasis are commonly emphasized, the arrival of neutrophils and monocytes to sites of inflammation initiates anti-inflammatory signals that promote resolution. Specifically, leukocytes and epithelial cells promote resolution of inflammation by releasing factors such as annexin 1, resolvins, lipoxins, maresins, and protectins. These are released in a spatiotemporal manner and signal via G-protein–coupled receptors to coordinate resolution of inflammation and repair. Recruitment of additional neutrophils stimulates clearance of dead PMNs and microbes by macrophages and promotes repair of the epithelial barrier.164–166 Resolvin E1 reduces leukocyte infiltration, decreases inflammatory cytokines (Il12 subunit p40 and TNF), and prevents development of colitis in mice given 2,4,6-trinitrobenzene sulfonic acid.167 Engagement of the chemerin chemokine-like receptor 1 by Resolvin E1 inhibits recruitment of PMNs and mediates clearance of neutrophils, via induction of the anti-adhesion protein CD55.168 The release of such mediators does not appear to impair normal antimicrobial functions of PMNs because decreased infiltration is associated with increased phagocytic function.169,170
Similarly, highly regulated spatiotemporal interactions among mucosal cytokines, including inflammatory and anti-inflammatory mediators, not only serve in host defense but also ensure epithelial barrier repair. Resolution of inflammation and repair are active at specific times, in a spatially restricted manner, to ensure recovery of epithelial barrier function. During inflammation and repair, abundant inflammatory and anti-inflammatory mediators are released into the epithelial milieu and subsequently bind epithelial receptors to exert biologic responses. Concentrations of such mediators in the immediate vicinity of the epithelium are not known. However, it is reasonable to assume that during mucosal inflammation, recruitment of large numbers of leukocytes would result in greatly increased subepithelial levels of inflammatory mediators that are far higher than observed in the blood or bulk tissue.
Inflammatory cytokines can have detrimental effects on the epithelial barrier, although they also stimulate the synthesis of mediators that help restore mucosal homeostasis. The overall effects of cytokines on the epithelial barrier are a complex combination of their direct effects on the epithelium and their effects on innate and adaptive immune cells. Given the crucial role of immunity in host protection, the double duty of cytokines in eradication of pathogens and fortification of the epithelial barrier might have conserved evolutionary advantages. For example, inflammatory cytokines increase expression of casein kinase 2α by intestinal epithelial cells,171 which protects IECs from apoptosis by influencing caspase 9 and caspase 3 activity.172 In addition, creatine kinase phosphorylates and facilitates TJ occludin assembly, thereby contributing to TJ remodeling and barrier regulation.173 In keratinocytes, creatine kinase has been reported to phosphorylate α-catenin and E-cadherin,174 thereby strengthening intercellular adhesion.175 As another example, production of IL23 during intestinal inflammation has been reported to induce production of IL17A and IL22, which in turn promote recruitment of innate and adaptive immune cells, host defense, and mucosal homeostasis.176 Although IL17A is considered to be an inflammatory cytokine, it also protects the barrier via changes in occludin.177,178 It is, however, unclear how IL17A affects AJC protein remodeling and barrier function.
Another cytokine that is potently induced by IL23 is IL22.179,180 This cytokine is up-regulated after gastrointestinal infection or damage and promotes tissue regeneration, barrier formation, and antimicrobial defense.181 IL22 effects are mediated by binding to the IL22RA1–IL10R2 receptor complex on epithelial cells resulting in STAT3 activation, induction of cellular proliferation and, ultimately, barrier fortification.181–183 Although IL22 is produced by innate lymphoid cells and Th22 lymphocytes,179 recent evidence suggests that, in response to IL23 and TNF, infiltrating PMNs are also significant producers of IL22, promoting antimicrobial defense, including the antibacterial peptides RegIIIβ and S100A8, and the restitution of colonic epithelial integrity during acute colitis.176 Thus, inflammatory cytokines that are associated with intestinal inflammation and barrier damage can also paradoxically contribute to barrier repair and fortification.
The dynamic interactions between inflammatory cytokines and barrier function were highlighted in recent studies that demonstrated a role for the IL36 receptor (an IL1 family member) in promoting intestinal epithelial barrier repair through recruitment of neutrophils and release of IL22.184,185 The protective effects of IL36 ligands in the epithelium might depend, in part, on IL22-mediated induction of the antimicrobial peptides S100A8/9 and regenerating islet-derived 3 gamma (REG3G). IL10 is also involved in the epithelial restitution induced during intestinal inflammation. IFNγ has been reported to induce intestinal epithelial cell expression of the IL10 receptor and signaling, resulting in barrier-protective effects.186–190 Cytokines mediate induction of barrier protective antimicrobial peptides, which highlights a critical balance that must be maintained between the mucosa and luminal microbiota. Similarly, the luminal microbiota play key roles in regulation of epithelial homeostasis. Microbial composition and products modulate epithelial barrier function by acting directly on IECs (as reported for microbial-derived short-chain fatty acids191) or indirectly, by contributing to the recruitment of immune cells that produce IL17 and IL22.192,193
There is accumulating evidence that certain inflammatory cytokines contribute to barrier protection. These functions need to be considered when developing therapies that target inflammatory cytokines for the treatment of intestinal inflammation.
Conclusions
The intestinal epithelial barrier is at the interface between luminal microbes and the mucosal immune system and, therefore, helps maintain mucosal homeostasis. This barrier is not only determined by complex interactions between many TJ proteins, but is also regulated by proteins within other intercellular junction complexes. Precise regulation of barrier function and efficient repair after injury are essential for preventing pathologic inflammation. During mucosal inflammation, recruited leukocytes directly interact with the epithelium, resulting in the release of inflammatory mediators, which have potent effects on barrier function and mucosal homeostasis. Cytokines released into the epithelial milieu target intercellular junction proteins and alter barrier function. Spatiotemporal release of pro-inflammatory and anti-inflammatory mediators, as well as pro-resolution proteins and lipids ensure restoration of mucosal homeostasis. Increasing our understanding of the mechanisms of inflammation-dependent alterations in epithelial permeability will provide new ideas for development of therapeutics to improve mucosal healing and barrier function during disease.
Acknowledgments
The authors thank Timothy L. Denning (Georgia State University) for helpful comments and Robin Kunkel (University of Michigan) for assistance in figure preparation. The authors also acknowledge the many significant contributions made by other investigators over the years who were not able to be referenced because of space and length restrictions.
Funding
Research programs in the authors laboratories are supported by the NIH (DK072564, DK061379, DK079392 to Charles A. Parkos and DK055679, DK059888 to Asma Nusrat) and Department of Defense PR121194.
Abbreviations used in this paper
- AJ
adherens junction
- AJC
apical junctional complex
- CAR
Coxsackie and adenovirus receptor
- CD
Crohn’s disease
- CPE
Clostridium perfringens enterotoxin
- DM
desmosome
- IBD
inflammatory bowel disease
- IEC
intestinal epithelial cell
- IFN
interferon
- IL
interleukin
- JAM
junctional adhesion molecule
- JAML
junctional adhesion molecule–like
- LP
lamina propria
- MLCK
myosin light chain kinase
- MUPP-1
multi-PDZ domain protein 1
- NM II
nonmuscle myosin II
- PAR
protease-activated receptor
- PDZ
PSD95–DlgA–ZO1
- PMN
polymorphonuclear neutrophil
- RMLC
regulatory myosin light chain
- TEpM
transepithelial migration
- TJ
tight junction
- TNF
tumor necrosis factor
- UC
ulcerative colitis
- ZO
zonula occludens
Footnotes
Conflicts of interest
The authors disclose no conflicts.
References
Author names in bold designate shared co-first authorship.
- 1.Helander HF, Fandriks L. Surface area of the digestive tract—revisited. Scand J Gastroenterol. 2014;49:681–689. doi: 10.3109/00365521.2014.898326. [DOI] [PubMed] [Google Scholar]
- 2.Abraham C, Medzhitov R. Interactions between the host innate immune system and microbes in inflammatory bowel disease. Gastroenterology. 2011;140:1729–1737. doi: 10.1053/j.gastro.2011.02.012. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 3.Ordas I, Eckmann L, Talamini M, et al. Ulcerative colitis. Lancet. 2012;380:1606–1619. doi: 10.1016/S0140-6736(12)60150-0. [DOI] [PubMed] [Google Scholar]
- 4.Katz KD, Hollander D, Vadheim CM, et al. Intestinal permeability in patients with Crohn’s disease and their healthy relatives. Gastroenterology. 1989;97:927–931. doi: 10.1016/0016-5085(89)91499-6. [DOI] [PubMed] [Google Scholar]
- 5.Peeters M, Geypens B, Claus D, et al. Clustering of increased small intestinal permeability in families with Crohn’s disease. Gastroenterology. 1997;113:802–807. doi: 10.1016/s0016-5085(97)70174-4. [DOI] [PubMed] [Google Scholar]
- 6.Neurath MF. Cytokines in inflammatory bowel disease. Nat Rev Immunol. 2014;14:329–342. doi: 10.1038/nri3661. [DOI] [PubMed] [Google Scholar]
- 7.Kamada N, Seo SU, Chen GY, et al. Role of the gut microbiota in immunity and inflammatory disease. Nat Rev Immunol. 2013;13:321–335. doi: 10.1038/nri3430. [DOI] [PubMed] [Google Scholar]
- 8.Kim YS, Ho SB. Intestinal goblet cells and mucins in health and disease: recent insights and progress. Curr Gastroenterol Rep. 2010;12:319–330. doi: 10.1007/s11894-010-0131-2. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 9.Pelaseyed T, Bergstrom JH, Gustafsson JK, et al. The mucus and mucins of the goblet cells and enterocytes provide the first defense line of the gastrointestinal tract and interact with the immune system. Immunol Rev. 2014;260:8–20. doi: 10.1111/imr.12182. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 10.van der Flier LG, Clevers H. Stem cells, self-renewal, and differentiation in the intestinal epithelium. Annu Rev Physiol. 2009;71:241–260. doi: 10.1146/annurev.physiol.010908.163145. [DOI] [PubMed] [Google Scholar]
- 11.Lavelle EC, Murphy C, O’Neill LA, et al. The role of TLRs, NLRs, and RLRs in mucosal innate immunity and homeostasis. Mucosal Immunol. 2010;3:17–28. doi: 10.1038/mi.2009.124. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 12.Stadnyk AW. Intestinal epithelial cells as a source of inflammatory cytokines and chemokines. Can J Gastroenterol. 2002;16:241–246. doi: 10.1155/2002/941087. [DOI] [PubMed] [Google Scholar]
- 13.Peterson LW, Artis D. Intestinal epithelial cells: regulators of barrier function and immune homeostasis. Nat Rev Immunol. 2014;14:141–153. doi: 10.1038/nri3608. [DOI] [PubMed] [Google Scholar]
- 14.Cereijido M, Valdes J, Shoshani L, et al. Role of tight junctions in establishing and maintaining cell polarity. Annu Rev Physiol. 1998;60:161–177. doi: 10.1146/annurev.physiol.60.1.161. [DOI] [PubMed] [Google Scholar]
- 15.Tsukita S, Furuse M, Itoh M. Multifunctional strands in tight junctions. Nat Rev Mol Cell Biol. 2001;2:285–293. doi: 10.1038/35067088. [DOI] [PubMed] [Google Scholar]
- 16.Watson CJ, Rowland M, Warhurst G. Functional modeling of tight junctions in intestinal cell monolayers using polyethylene glycol oligomers. Am J Physiol Cell Physiol. 2001;281:C388–C397. doi: 10.1152/ajpcell.2001.281.2.C388. [DOI] [PubMed] [Google Scholar]
- 17.Van Itallie CM, Holmes J, Bridges A, et al. The density of small tight junction pores varies among cell types and is increased by expression of claudin-2. J Cell Sci. 2008;121:298–305. doi: 10.1242/jcs.021485. [DOI] [PubMed] [Google Scholar]
- 18.Koval M. Differential pathways of claudin oligomerization and integration into tight junctions. Tissue Barriers. 2013;1:e24518. doi: 10.4161/tisb.24518. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 19.Furuse M, Fujita K, Hiiragi T, et al. Claudin-1 and -2: novel integral membrane proteins localizing at tight junctions with no sequence similarity to occludin. J Cell Biol. 1998;141:1539–1550. doi: 10.1083/jcb.141.7.1539. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 20.Anderson JM, Van Itallie CM. Physiology and function of the tight junction. Cold Spring Harb Perspect Biol. 2009;1:a002584. doi: 10.1101/cshperspect.a002584. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 21.Suzuki H, Nishizawa T, Tani K, et al. Crystal structure of a claudin provides insight into the architecture of tight junctions. Science. 2014;344:304–307. doi: 10.1126/science.1248571. [DOI] [PubMed] [Google Scholar]
- 22.Capaldo CT, Nusrat A. Claudin switching: physiological plasticity of the tight junction. Semin Cell Dev Biol. 2015;42:22–29. doi: 10.1016/j.semcdb.2015.04.003. [DOI] [PubMed] [Google Scholar]
- 23.Gunzel D, Yu AS. Claudins and the modulation of tight junction permeability. Physiol Rev. 2013;93:525–569. doi: 10.1152/physrev.00019.2012. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 24.Furuse M. Molecular basis of the core structure of tight junctions. Cold Spring Harb Perspect Biol. 2010;2:a002907. doi: 10.1101/cshperspect.a002907. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 25.Ding L, Lu Z, Foreman O, et al. Inflammation and disruption of the mucosal architecture in claudin-7-deficient mice. Gastroenterology. 2012;142:305–315. doi: 10.1053/j.gastro.2011.10.025. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 26.Tanaka H, Takechi M, Kiyonari H, et al. Intestinal deletion of Claudin-7 enhances paracellular organic solute flux and initiates colonic inflammation in mice. Gut. 2015;64:1529–1538. doi: 10.1136/gutjnl-2014-308419. [DOI] [PubMed] [Google Scholar]
- 27.Tamura A, Hayashi H, Imasato M, et al. Loss of claudin-15, but not claudin-2, causes Na+ deficiency and glucose malabsorption in mouse small intestine. Gastroenterology. 2011;140:913–923. doi: 10.1053/j.gastro.2010.08.006. [DOI] [PubMed] [Google Scholar]
- 28.Ahmad R, Chaturvedi R, Olivares-Villagomez D, et al. Targeted colonic claudin-2 expression renders resistance to epithelial injury, induces immune suppression, and protects from colitis. Mucosal Immunol. 2014;7:1340–1353. doi: 10.1038/mi.2014.21. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 29.Nishida M, Yoshida M, Nishiumi S, et al. Claudin-2 regulates colorectal inflammation via myosin light chain kinase-dependent signaling. Dig Dis Sci. 2013;58:1546–1559. doi: 10.1007/s10620-012-2535-3. [DOI] [PubMed] [Google Scholar]
- 30.Luettig J, Rosenthal R, Barmeyer C, et al. Claudin-2 as a mediator of leaky gut barrier during intestinal inflammation. Tissue Barriers. 2015;3:e977176. doi: 10.4161/21688370.2014.977176. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 31.Tamura A, Kitano Y, Hata M, et al. Megaintestine in claudin-15-deficient mice. Gastroenterology. 2008;134:523–534. doi: 10.1053/j.gastro.2007.11.040. [DOI] [PubMed] [Google Scholar]
- 32.Wada M, Tamura A, Takahashi N, et al. Loss of claudins 2 and 15 from mice causes defects in paracellular Na+ flow and nutrient transport in gut and leads to death from malnutrition. Gastroenterology. 2013;144:369–380. doi: 10.1053/j.gastro.2012.10.035. [DOI] [PubMed] [Google Scholar]
- 33.Katahira J, Sugiyama H, Inoue N, et al. Clostridium per-fringens enterotoxin utilizes two structurally related membrane proteins as functional receptors in vivo. J Biol Chem. 1997;272:26652–26658. doi: 10.1074/jbc.272.42.26652. [DOI] [PubMed] [Google Scholar]
- 34.Fernandez Miyakawa ME, Pistone Creydt V, Uzal FA, et al. Clostridium perfringens enterotoxin damages the human intestine in vitro. Infect Immun. 2005;73:8407–8410. doi: 10.1128/IAI.73.12.8407-8410.2005. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 35.Veshnyakova A, Protze J, Rossa J, et al. On the interaction of Clostridium perfringens enterotoxin with claudins. Toxins (Basel) 2010;2:1336–1356. doi: 10.3390/toxins2061336. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 36.Saitoh Y, Suzuki H, Tani K, et al. Tight junctions. Structural insight into tight junction disassembly by Clostridium perfringens enterotoxin. Science. 2015;347:775–778. doi: 10.1126/science.1261833. [DOI] [PubMed] [Google Scholar]
- 37.Fedwick JP, Lapointe TK, Meddings JB, et al. Helicobacter pylori activates myosin light-chain kinase to disrupt claudin-4 and claudin-5 and increase epithelial permeability. Infect Immun. 2005;73:7844–7852. doi: 10.1128/IAI.73.12.7844-7852.2005. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 38.Evans MJ, von Hahn T, Tscherne DM, et al. Claudin-1 is a hepatitis C virus co-receptor required for a late step in entry. Nature. 2007;446:801–805. doi: 10.1038/nature05654. [DOI] [PubMed] [Google Scholar]
- 39.Raleigh DR, Marchiando AM, Zhang Y, et al. Tight junction-associated MARVEL proteins marveld3, tricellulin, and occludin have distinct but overlapping functions. Mol Biol Cell. 2010;21:1200–1213. doi: 10.1091/mbc.E09-08-0734. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 40.Moroi S, Saitou M, Fujimoto K, et al. Occludin is concentrated at tight junctions of mouse/rat but not human/guinea pig Sertoli cells in testes. Am J Physiol. 1998;274:C1708–C1717. doi: 10.1152/ajpcell.1998.274.6.C1708. [DOI] [PubMed] [Google Scholar]
- 41.Saitou M, Fujimoto K, Doi Y, et al. Occludin-deficient embryonic stem cells can differentiate into polarized epithelial cells bearing tight junctions. J Cell Biol. 1998;141:397–408. doi: 10.1083/jcb.141.2.397. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 42.Saitou M, Furuse M, Sasaki H, et al. Complex phenotype of mice lacking occludin, a component of tight junction strands. Mol Biol Cell. 2000;11:4131–4142. doi: 10.1091/mbc.11.12.4131. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 43.Schulzke JD, Gitter AH, Mankertz J, et al. Epithelial transport and barrier function in occludin-deficient mice. Biochim Biophys Acta. 2005;1669:34–42. doi: 10.1016/j.bbamem.2005.01.008. [DOI] [PubMed] [Google Scholar]
- 44.Al-Sadi R, Khatib K, Guo S, et al. Occludin regulates macromolecule flux across the intestinal epithelial tight junction barrier. Am J Physiol Gastrointest Liver Physiol. 2011;300:G1054–G1064. doi: 10.1152/ajpgi.00055.2011. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 45.Buschmann MM, Shen L, Rajapakse H, et al. Occludin OCEL-domain interactions are required for maintenance and regulation of the tight junction barrier to macromolecular flux. Mol Biol Cell. 2013;24:3056–3068. doi: 10.1091/mbc.E12-09-0688. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 46.Everett RS, Vanhook MK, Barozzi N, et al. Specific modulation of airway epithelial tight junctions by apical application of an occludin peptide. Mol Pharmacol. 2006;69:492–500. doi: 10.1124/mol.105.017251. [DOI] [PubMed] [Google Scholar]
- 47.Nusrat A, Brown GT, Tom J, et al. Multiple protein interactions involving proposed extracellular loop domains of the tight junction protein occludin. Mol Biol Cell. 2005;16:1725–1734. doi: 10.1091/mbc.E04-06-0465. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 48.Wong V, Gumbiner BM. A synthetic peptide corresponding to the extracellular domain of occludin perturbs the tight junction permeability barrier. J Cell Biol. 1997;136:399–409. doi: 10.1083/jcb.136.2.399. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 49.Fanning AS, Jameson BJ, Jesaitis LA, et al. The tight junction protein ZO-1 establishes a link between the transmembrane protein occludin and the actin cytoskeleton. J Biol Chem. 1998;273:29745–29753. doi: 10.1074/jbc.273.45.29745. [DOI] [PubMed] [Google Scholar]
- 50.Furuse M, Itoh M, Hirase T, et al. Direct association of occludin with ZO-1 and its possible involvement in the localization of occludin at tight junctions. J Cell Biol. 1994;127:1617–1626. doi: 10.1083/jcb.127.6.1617. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 51.Cording J, Berg J, Kading N, et al. In tight junctions, claudins regulate the interactions between occludin, tricellulin and marvelD3, which, inversely, modulate claudin oligomerization. J Cell Sci. 2013;126:554–564. doi: 10.1242/jcs.114306. [DOI] [PubMed] [Google Scholar]
- 52.Steed E, Elbediwy A, Vacca B, et al. MarvelD3 couples tight junctions to the MEKK1-JNK pathway to regulate cell behavior and survival. J Cell Biol. 2014;204:821–838. doi: 10.1083/jcb.201304115. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 53.Furuse M, Izumi Y, Oda Y, et al. Molecular organization of tricellular tight junctions. Tissue Barriers. 2014;2:e28960. doi: 10.4161/tisb.28960. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 54.Ikenouchi J, Furuse M, Furuse K, et al. Tricellulin constitutes a novel barrier at tricellular contacts of epithelial cells. J Cell Biol. 2005;171:939–945. doi: 10.1083/jcb.200510043. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 55.Krug SM, Amasheh S, Richter JF, et al. Tricellulin forms a barrier to macromolecules in tricellular tight junctions without affecting ion permeability. Mol Biol Cell. 2009;20:3713–3724. doi: 10.1091/mbc.E09-01-0080. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 56.Fukumatsu M, Ogawa M, Arakawa S, et al. Shigella targets epithelial tricellular junctions and uses a noncanonical clathrin-dependent endocytic pathway to spread between cells. Cell Host Microbe. 2012;11:325–336. doi: 10.1016/j.chom.2012.03.001. [DOI] [PubMed] [Google Scholar]
- 57.Eum SY, Jaraki D, Bertrand L, et al. Disruption of epithelial barrier by quorum-sensing N-3-(oxododecanoyl)-homoserine lactone is mediated by matrix metal-loproteinases. Am J Physiol Gastrointest Liver Physiol. 2014;306:G992–G1001. doi: 10.1152/ajpgi.00016.2014. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 58.Papatheodorou P, Carette JE, Bell GW, et al. Lipolysis-stimulated lipoprotein receptor (LSR) is the host receptor for the binary toxin Clostridium difficile transferase (CDT) Proc Natl Acad Sci U S A. 2011;108:16422–16427. doi: 10.1073/pnas.1109772108. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 59.Ebnet K, Suzuki A, Ohno S, et al. Junctional adhesion molecules (JAMs): more molecules with dual functions? J Cell Sci. 2004;117:19–29. doi: 10.1242/jcs.00930. [DOI] [PubMed] [Google Scholar]
- 60.Luissint AC, Nusrat A, Parkos CA. JAM-related proteins in mucosal homeostasis and inflammation. Semin Immunopathol. 2014;36:211–226. doi: 10.1007/s00281-014-0421-0. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 61.Kostrewa D, Brockhaus M, D’Arcy A, et al. X-ray structure of junctional adhesion molecule: structural basis for homophilic adhesion via a novel dimerization motif. EMBO J. 2001;20:4391–4398. doi: 10.1093/emboj/20.16.4391. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 62.Prota AE, Campbell JA, Schelling P, et al. Crystal structure of human junctional adhesion molecule 1: implications for reovirus binding. Proc Natl Acad Sci U S A. 2003;100:5366–5371. doi: 10.1073/pnas.0937718100. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 63.Liu Y, Nusrat A, Schnell FJ, et al. Human junction adhesion molecule regulates tight junction resealing in epithelia. J Cell Sci. 2000;113(Pt 13):2363–2374. doi: 10.1242/jcs.113.13.2363. [DOI] [PubMed] [Google Scholar]
- 64.Laukoetter MG, Nava P, Lee WY, et al. JAM-A regulates permeability and inflammation in the intestine in vivo. J Exp Med. 2007;204:3067–3076. doi: 10.1084/jem.20071416. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 65.Khounlotham M, Kim W, Peatman E, et al. Compromised intestinal epithelial barrier induces adaptive immune compensation that protects from colitis. Immunity. 2012;37:563–573. doi: 10.1016/j.immuni.2012.06.017. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 66.Itoh M, Sasaki H, Furuse M, et al. Junctional adhesion molecule (JAM) binds to PAR-3: a possible mechanism for the recruitment of PAR-3 to tight junctions. J Cell Biol. 2001;154:491–497. doi: 10.1083/jcb.200103047. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 67.Monteiro AC, Sumagin R, Rankin CR, et al. JAM-A associates with ZO-2, afadin, and PDZ-GEF1 to activate Rap2c and regulate epithelial barrier function. Mol Biol Cell. 2013;24:2849–2860. doi: 10.1091/mbc.E13-06-0298. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 68.Barton ES, Forrest JC, Connolly JL, et al. Junction adhesion molecule is a receptor for reovirus. Cell. 2001;104:441–451. doi: 10.1016/s0092-8674(01)00231-8. [DOI] [PubMed] [Google Scholar]
- 69.Coyne CB, Bergelson JM. CAR: a virus receptor within the tight junction. Adv Drug Deliv Rev. 2005;57:869–882. doi: 10.1016/j.addr.2005.01.007. [DOI] [PubMed] [Google Scholar]
- 70.Pazirandeh A, Sultana T, Mirza M, et al. Multiple phenotypes in adult mice following inactivation of the cox-sackievirus and adenovirus receptor (Car) gene. PLoS One. 2011;6:e20203. doi: 10.1371/journal.pone.0020203. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 71.Morton PE, Hicks A, Nastos T, et al. CAR regulates epithelial cell junction stability through control of E-cadherin trafficking. Sci Rep. 2013;3:2889. doi: 10.1038/srep02889. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 72.Matter K, Balda MS. Epithelial tight junctions, gene expression and nucleojunctional interplay. J Cell Sci. 2007;120:1505–1511. doi: 10.1242/jcs.005975. [DOI] [PubMed] [Google Scholar]
- 73.Paris L, Tonutti L, Vannini C, et al. Structural organization of the tight junctions. Biochim Biophys Acta. 2008;1778:646–659. doi: 10.1016/j.bbamem.2007.08.004. [DOI] [PubMed] [Google Scholar]
- 74.Aijaz S, Balda MS, Matter K. Tight junctions: molecular architecture and function. Int Rev Cytol. 2006;248:261–298. doi: 10.1016/S0074-7696(06)48005-0. [DOI] [PubMed] [Google Scholar]
- 75.Fanning AS, Ma TY, Anderson JM. Isolation and functional characterization of the actin binding region in the tight junction protein ZO-1. FASEB J. 2002;16:1835–1837. doi: 10.1096/fj.02-0121fje. [DOI] [PubMed] [Google Scholar]
- 76.Yamamoto T, Harada N, Kano K, et al. The Ras target AF-6 interacts with ZO-1 and serves as a peripheral component of tight junctions in epithelial cells. J Cell Biol. 1997;139:785–795. doi: 10.1083/jcb.139.3.785. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 77.Ullmer C, Schmuck K, Figge A, et al. Cloning and characterization of MUPP1, a novel PDZ domain protein. FEBS Lett. 1998;424:63–68. doi: 10.1016/s0014-5793(98)00141-0. [DOI] [PubMed] [Google Scholar]
- 78.Ide N, Hata Y, Nishioka H, et al. Localization of membrane-associated guanylate kinase (MAGI)-1/BAI-associated protein (BAP) 1 at tight junctions of epithelial cells. Oncogene. 1999;18:7810–7815. doi: 10.1038/sj.onc.1203153. [DOI] [PubMed] [Google Scholar]
- 79.Joberty G, Petersen C, Gao L, et al. The cell-polarity protein Par6 links Par3 and atypical protein kinase C to Cdc42. Nat Cell Biol. 2000;2:531–539. doi: 10.1038/35019573. [DOI] [PubMed] [Google Scholar]
- 80.Gonzalez-Mariscal L, Tapia R, Chamorro D. Crosstalk of tight junction components with signaling pathways. Biochim Biophys Acta. 2008;1778:729–756. doi: 10.1016/j.bbamem.2007.08.018. [DOI] [PubMed] [Google Scholar]
- 81.Bazzoni G, Martinez-Estrada OM, Orsenigo F, et al. Interaction of junctional adhesion molecule with the tight junction components ZO-1, cingulin, and occludin. J Biol Chem. 2000;275:20520–20526. doi: 10.1074/jbc.M905251199. [DOI] [PubMed] [Google Scholar]
- 82.Cordenonsi M, D’Atri F, Hammar E, et al. Cingulin contains globular and coiled-coil domains and interacts with ZO-1, ZO-2, ZO-3, and myosin. J Cell Biol. 1999;147:1569–1582. doi: 10.1083/jcb.147.7.1569. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 83.Cordenonsi M, Turco F, D’Atri F, et al. Xenopus laevis occludin. Identification of in vitro phosphorylation sites by protein kinase CK2 and association with cingulin. Eur J Biochem. 1999;264:374–384. doi: 10.1046/j.1432-1327.1999.00616.x. [DOI] [PubMed] [Google Scholar]
- 84.D’Atri F, Citi S. Cingulin interacts with F-actin in vitro. FEBS Lett. 2001;507:21–24. doi: 10.1016/s0014-5793(01)02936-2. [DOI] [PubMed] [Google Scholar]
- 85.Scharl M, Paul G, Weber A, et al. Protection of epithelial barrier function by the Crohn’s disease associated gene protein tyrosine phosphatase n2. Gastroenterology. 2009;137:2030–2040. e5. doi: 10.1053/j.gastro.2009.07.078. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 86.Citalan-Madrid AF, Garcia-Ponce A, Vargas-Robles H, et al. Small GTPases of the Ras superfamily regulate intestinal epithelial homeostasis and barrier function via common and unique mechanisms. Tissue Barriers. 2013;1:e26938. doi: 10.4161/tisb.26938. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 87.McCole DF. Phosphatase regulation of intercellular junctions. Tissue Barriers. 2013;1:e26713. doi: 10.4161/tisb.26713. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 88.Hartsock A, Nelson WJ. Adherens and tight junctions: structure, function and connections to the actin cytoskeleton. Biochim Biophys Acta. 2008;1778:660–669. doi: 10.1016/j.bbamem.2007.07.012. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 89.Holthofer B, Windoffer R, Troyanovsky S, et al. Structure and function of desmosomes. Int Rev Cytol. 2007;264:65–163. doi: 10.1016/S0074-7696(07)64003-0. [DOI] [PubMed] [Google Scholar]
- 90.Takai Y, Nakanishi H. Nectin and afadin: novel organizers of intercellular junctions. J Cell Sci. 2003;116:17–27. doi: 10.1242/jcs.00167. [DOI] [PubMed] [Google Scholar]
- 91.Ivanov AI, Naydenov NG. Dynamics and regulation of epithelial adherens junctions: recent discoveries and controversies. Int Rev Cell Mol Biol. 2013;303:27–99. doi: 10.1016/B978-0-12-407697-6.00002-7. [DOI] [PubMed] [Google Scholar]
- 92.Hermiston ML, Gordon JI. Inflammatory bowel disease and adenomas in mice expressing a dominant negative N-cadherin. Science. 1995;270:1203–1207. doi: 10.1126/science.270.5239.1203. [DOI] [PubMed] [Google Scholar]
- 93.Schneider MR, Dahlhoff M, Horst D, et al. A key role for E-cadherin in intestinal homeostasis and Paneth cell maturation. PLoS One. 2010;5:e14325. doi: 10.1371/journal.pone.0014325. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 94.Smalley-Freed WG, Efimov A, Burnett PE, et al. p120-catenin is essential for maintenance of barrier function and intestinal homeostasis in mice. J Clin Invest. 2010;120:1824–1835. doi: 10.1172/JCI41414. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 95.Madara JL. Intestinal absorptive cell tight junctions are linked to cytoskeleton. Am J Physiol. 1987;253:C171–C175. doi: 10.1152/ajpcell.1987.253.1.C171. [DOI] [PubMed] [Google Scholar]
- 96.Meng W, Takeichi M. Adherens junction: molecular architecture and regulation. Cold Spring Harb Perspect Biol. 2009;1:a002899. doi: 10.1101/cshperspect.a002899. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 97.Ivanov AI, Parkos CA, Nusrat A. Cytoskeletal regulation of epithelial barrier function during inflammation. Am J Pathol. 2010;177:512–524. doi: 10.2353/ajpath.2010.100168. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 98.Vicente-Manzanares M, Ma X, Adelstein RS, et al. Non-muscle myosin II takes centre stage in cell adhesion and migration. Nat Rev Mol Cell Biol. 2009;10:778–790. doi: 10.1038/nrm2786. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 99.Ivanov AI, Bachar M, Babbin BA, et al. A unique role for nonmuscle myosin heavy chain IIA in regulation of epithelial apical junctions. PLoS One. 2007;2:e658. doi: 10.1371/journal.pone.0000658. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 100.Ivanov AI, McCall IC, Parkos CA, et al. Role for actin filament turnover and a myosin II motor in cytoskeleton-driven disassembly of the epithelial apical junctional complex. Mol Biol Cell. 2004;15:2639–2651. doi: 10.1091/mbc.E04-02-0163. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 101.Ivanov AI, Samarin SN, Bachar M, et al. Protein kinase C activation disrupts epithelial apical junctions via ROCK-II dependent stimulation of actomyosin contractility. BMC Cell Biol. 2009;10:36. doi: 10.1186/1471-2121-10-36. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 102.Naydenov NG, Feygin A, Wang D, et al. Nonmuscle myosin IIA regulates intestinal epithelial barrier in vivo and plays a protective role during experimental colitis. Sci Rep. 2016;6:24161. doi: 10.1038/srep24161. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 103.Su L, Shen L, Clayburgh DR, et al. Targeted epithelial tight junction dysfunction causes immune activation and contributes to development of experimental colitis. Gastroenterology. 2009;136:551–563. doi: 10.1053/j.gastro.2008.10.081. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 104.Green KJ, Simpson CL. Desmosomes: new perspectives on a classic. J Invest Dermatol. 2007;127:2499–2515. doi: 10.1038/sj.jid.5701015. [DOI] [PubMed] [Google Scholar]
- 105.Donaldson GP, Lee SM, Mazmanian SK. Gut biogeography of the bacterial microbiota. Nat Rev Microbiol. 2016;14:20–32. doi: 10.1038/nrmicro3552. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 106.Ley K, Laudanna C, Cybulsky MI, et al. Getting to the site of inflammation: the leukocyte adhesion cascade updated. Nat Rev Immunol. 2007;7:678–689. doi: 10.1038/nri2156. [DOI] [PubMed] [Google Scholar]
- 107.Parkos CA, Delp C, Arnaout MA, et al. Neutrophil migration across a cultured intestinal epithelium. Dependence on a CD11b/CD18-mediated event and enhanced efficiency in physiological direction. J Clin Invest. 1991;88:1605–1612. doi: 10.1172/JCI115473. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 108.Madara JL, Nash S, Parkos C. Neutrophil-epithelial cell interactions in the intestine. Adv Exp Med Biol. 1991;314:329–334. doi: 10.1007/978-1-4684-6024-7_22. [DOI] [PubMed] [Google Scholar]
- 109.Nusrat A, Parkos CA, Liang TW, et al. Neutrophil migration across model intestinal epithelia: monolayer disruption and subsequent events in epithelial repair. Gastroenterology. 1997;113:1489–1500. doi: 10.1053/gast.1997.v113.pm9352851. [DOI] [PubMed] [Google Scholar]
- 110.Parkos CA. Neutrophil-epithelial interactions: a double-edged sword. Am J Pathol. 2016;186:1404–1416. doi: 10.1016/j.ajpath.2016.02.001. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 111.Zen K, Liu Y, Cairo D, et al. CD11b/CD18-dependent interactions of neutrophils with intestinal epithelium are mediated by fucosylated proteoglycans. J Immunol. 2002;169:5270–5278. doi: 10.4049/jimmunol.169.9.5270. [DOI] [PubMed] [Google Scholar]
- 112.Blake KM, Carrigan SO, Issekutz AC, et al. Neutrophils migrate across intestinal epithelium using beta2 integrin (CD11b/CD18)-independent mechanisms. Clin Exp Immunol. 2004;136:262–268. doi: 10.1111/j.1365-2249.2004.02429.x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 113.Lee WY, Weber DA, Laur O, et al. The role of cis dimerization of signal regulatory protein alpha (SIRPalpha) in binding to CD47. J Biol Chem. 2010;285:37953–37963. doi: 10.1074/jbc.M110.180018. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 114.Liu Y, Merlin D, Burst SL, et al. The role of CD47 in neutrophil transmigration. Increased rate of migration correlates with increased cell surface expression of CD47. J Biol Chem. 2001;276:40156–40166. doi: 10.1074/jbc.M104138200. [DOI] [PubMed] [Google Scholar]
- 115.Zen K, Liu Y, McCall IC, et al. Neutrophil migration across tight junctions is mediated by adhesive interactions between epithelial coxsackie and adenovirus receptor and a junctional adhesion molecule-like protein on neutrophils. Mol Biol Cell. 2005;16:2694–2703. doi: 10.1091/mbc.E05-01-0036. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 116.Sumagin R, Robin AZ, Nusrat A, et al. Transmigrated neutrophils in the intestinal lumen engage ICAM-1 to regulate the epithelial barrier and neutrophil recruitment. Mucosal Immunol. 2014;7:905–915. doi: 10.1038/mi.2013.106. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 117.Chin AC, Lee WY, Nusrat A, et al. Neutrophil-mediated activation of epithelial protease-activated receptors-1 and -2 regulates barrier function and transepithelial migration. J Immunol. 2008;181:5702–5710. doi: 10.4049/jimmunol.181.8.5702. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 118.Ginzberg HH, Cherapanov V, Dong Q, et al. Neutrophil-mediated epithelial injury during transmigration: role of elastase. Am J Physiol Gastrointest Liver Physiol. 2001;281:G705–G717. doi: 10.1152/ajpgi.2001.281.3.G705. [DOI] [PubMed] [Google Scholar]
- 119.Zemans RL, Briones N, Campbell M, et al. Neutrophil transmigration triggers repair of the lung epithelium via beta-catenin signaling. Proc Natl Acad Sci U S A. 2011;108:15990–15995. doi: 10.1073/pnas.1110144108. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 120.Kamekura R, Nava P, Feng M, et al. Inflammation-induced desmoglein-2 ectodomain shedding compromises the mucosal barrier. Mol Biol Cell. 2015;26:3165–3177. doi: 10.1091/mbc.E15-03-0147. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 121.Weber DA, Sumagin R, McCall IC, et al. Neutrophil-derived JAML inhibits repair of intestinal epithelial injury during acute inflammation. Mucosal Immunol. 2014;7:1221–1232. doi: 10.1038/mi.2014.12. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 122.Colgan SP, Eltzschig HK, Eckle T, et al. Physiological roles for ecto-5’-nucleotidase (CD73) Purinergic Signal. 2006;2:351–360. doi: 10.1007/s11302-005-5302-5. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 123.Madara JL, Patapoff TW, Gillece-Castro B, et al. 5’-adenosine monophosphate is the neutrophil-derived paracrine factor that elicits chloride secretion from T84 intestinal epithelial cell monolayers. J Clin Invest. 1993;91:2320–2325. doi: 10.1172/JCI116462. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 124.Reinecker HC, Steffen M, Witthoeft T, et al. Enhanced secretion of tumour necrosis factor-alpha, IL-6, and IL-1 beta by isolated lamina propria mononuclear cells from patients with ulcerative colitis and Crohn’s disease. Clin Exp Immunol. 1993;94:174–181. doi: 10.1111/j.1365-2249.1993.tb05997.x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 125.Stallmach A, Giese T, Schmidt C, et al. Cytokine/chemokine transcript profiles reflect mucosal inflammation in Crohn’s disease. Int J Colorectal Dis. 2004;19:308–315. doi: 10.1007/s00384-003-0554-4. [DOI] [PubMed] [Google Scholar]
- 126.Ben-Horin S, Kopylov U, Chowers Y. Optimizing anti-TNF treatments in inflammatory bowel disease. Autoimmun Rev. 2014;13:24–30. doi: 10.1016/j.autrev.2013.06.002. [DOI] [PubMed] [Google Scholar]
- 127.Colombel JF, Sandborn WJ, Reinisch W, et al. Infliximab, azathioprine, or combination therapy for Crohn’s disease. N Engl J Med. 2010;362:1383–1395. doi: 10.1056/NEJMoa0904492. [DOI] [PubMed] [Google Scholar]
- 128.Colombel JF, Sandborn WJ, Rutgeerts P, et al. Adalimumab for maintenance of clinical response and remission in patients with Crohn’s disease: the CHARM trial. Gastroenterology. 2007;132:52–65. doi: 10.1053/j.gastro.2006.11.041. [DOI] [PubMed] [Google Scholar]
- 129.Evans AT, Lee SD. A review and expert opinion of the use of certolizumab for Crohn’s disease. Expert Opin Biol Ther. 2012;12:363–370. doi: 10.1517/14712598.2012.658770. [DOI] [PubMed] [Google Scholar]
- 130.Reinisch W, de Villiers W, Bene L, et al. Fontolizumab in moderate to severe Crohn’s disease: a phase 2, randomized, double-blind, placebo-controlled, multiple-dose study. Inflamm Bowel Dis. 2010;16:233–242. doi: 10.1002/ibd.21038. [DOI] [PubMed] [Google Scholar]
- 131.Sandborn WJ, Gasink C, Gao LL, et al. Ustekinumab induction and maintenance therapy in refractory Crohn’s disease. N Engl J Med. 2012;367:1519–1528. doi: 10.1056/NEJMoa1203572. [DOI] [PubMed] [Google Scholar]
- 132.Sandborn WJ, Hanauer SB, Katz S, et al. Etanercept for active Crohn’s disease: a randomized, double-blind, placebo-controlled trial. Gastroenterology. 2001;121:1088–1094. doi: 10.1053/gast.2001.28674. [DOI] [PubMed] [Google Scholar]
- 133.Abreu MT, Palladino AA, Arnold ET, et al. Modulation of barrier function during Fas-mediated apoptosis in human intestinal epithelial cells. Gastroenterology. 2000;119:1524–1536. doi: 10.1053/gast.2000.20232. [DOI] [PubMed] [Google Scholar]
- 134.Bruewer M, Luegering A, Kucharzik T, et al. Proinflammatory cytokines disrupt epithelial barrier function by apoptosis-independent mechanisms. J Immunol. 2003;171:6164–6172. doi: 10.4049/jimmunol.171.11.6164. [DOI] [PubMed] [Google Scholar]
- 135.Gassler N, Rohr C, Schneider A, et al. Inflammatory bowel disease is associated with changes of enterocytic junctions. Am J Physiol Gastrointest Liver Physiol. 2001;281:G216–G228. doi: 10.1152/ajpgi.2001.281.1.G216. [DOI] [PubMed] [Google Scholar]
- 136.Kucharzik T, Walsh SV, Chen J, et al. Neutrophil transmigration in inflammatory bowel disease is associated with differential expression of epithelial intercellular junction proteins. Am J Pathol. 2001;159:2001–2009. doi: 10.1016/S0002-9440(10)63051-9. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 137.Mankertz J, Tavalali S, Schmitz H, et al. Expression from the human occludin promoter is affected by tumor necrosis factor alpha and interferon gamma. J Cell Sci. 2000;113(Pt 11):2085–2090. doi: 10.1242/jcs.113.11.2085. [DOI] [PubMed] [Google Scholar]
- 138.Naydenov NG, Baranwal S, Khan S, et al. Novel mechanism of cytokine-induced disruption of epithelial barriers: Janus kinase and protein kinase D-dependent downregulation of junction protein expression. Tissue Barriers. 2013;1:e25231. doi: 10.4161/tisb.25231. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 139.Scharl M, Paul G, Barrett KE, et al. AMP-activated protein kinase mediates the interferon-gamma-induced decrease in intestinal epithelial barrier function. J Biol Chem. 2009;284:27952–27963. doi: 10.1074/jbc.M109.046292. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 140.Youakim A, Ahdieh M. Interferon-gamma decreases barrier function in T84 cells by reducing ZO-1 levels and disrupting apical actin. Am J Physiol. 1999;276:G1279–G1288. doi: 10.1152/ajpgi.1999.276.5.G1279. [DOI] [PubMed] [Google Scholar]
- 141.Wang F, Graham WV, Wang Y, et al. Interferon-gamma and tumor necrosis factor-alpha synergize to induce intestinal epithelial barrier dysfunction by upregulating myosin light chain kinase expression. Am J Pathol. 2005;166:409–419. doi: 10.1016/s0002-9440(10)62264-x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 142.Mankertz J, Amasheh M, Krug SM, et al. TNFalpha up-regulates claudin-2 expression in epithelial HT-29/B6 cells via phosphatidylinositol-3-kinase signaling. Cell Tissue Res. 2009;336:67–77. doi: 10.1007/s00441-009-0751-8. [DOI] [PubMed] [Google Scholar]
- 143.Weber CR, Raleigh DR, Su L, et al. Epithelial myosin light chain kinase activation induces mucosal interleukin-13 expression to alter tight junction ion selectivity. J Biol Chem. 2010;285:12037–12046. doi: 10.1074/jbc.M109.064808. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 144.Kinugasa T, Sakaguchi T, Gu X, et al. Claudins regulate the intestinal barrier in response to immune mediators. Gastroenterology. 2000;118:1001–1011. doi: 10.1016/s0016-5085(00)70351-9. [DOI] [PubMed] [Google Scholar]
- 145.Suzuki T, Yoshinaga N, Tanabe S. Interleukin-6 (IL-6) regulates claudin-2 expression and tight junction permeability in intestinal epithelium. J Biol Chem. 2011;286:31263–31271. doi: 10.1074/jbc.M111.238147. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 146.Capaldo CT, Farkas AE, Hilgarth RS, et al. Proinflammatory cytokine-induced tight junction remodeling through dynamic self-assembly of claudins. Mol Biol Cell. 2014;25:2710–2719. doi: 10.1091/mbc.E14-02-0773. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 147.Utech M, Mennigen R, Bruewer M. Endocytosis and recycling of tight junction proteins in inflammation. J Biomed Biotechnol. 2010;2010:484987. doi: 10.1155/2010/484987. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 148.Bruewer M, Utech M, Ivanov AI, et al. Interferon-gamma induces internalization of epithelial tight junction proteins via a macropinocytosis-like process. FASEB J. 2005;19:923–933. doi: 10.1096/fj.04-3260com. [DOI] [PubMed] [Google Scholar]
- 149.Marchiando AM, Shen L, Graham WV, et al. Caveolin-1-dependent occludin endocytosis is required for TNF-induced tight junction regulation in vivo. J Cell Biol. 2010;189:111–126. doi: 10.1083/jcb.200902153. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 150.Ivanov AI, Nusrat A, Parkos CA. Endocytosis of epithelial apical junctional proteins by a clathrin-mediated pathway into a unique storage compartment. Mol Biol Cell. 2004;15:176–188. doi: 10.1091/mbc.E03-05-0319. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 151.Kevil CG, Ohno N, Gute DC, et al. Role of cadherin internalization in hydrogen peroxide-mediated endothelial permeability. Free Radic Biol Med. 1998;24:1015–1022. doi: 10.1016/s0891-5849(97)00433-4. [DOI] [PubMed] [Google Scholar]
- 152.Van Itallie CM, Fanning AS, Holmes J, et al. Occludin is required for cytokine-induced regulation of tight junction barriers. J Cell Sci. 2010;123:2844–2852. doi: 10.1242/jcs.065581. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 153.Utech M, Ivanov AI, Samarin SN, et al. Mechanism of IFN-gamma-induced endocytosis of tight junction proteins: myosin II-dependent vacuolarization of the apical plasma membrane. Mol Biol Cell. 2005;16:5040–5052. doi: 10.1091/mbc.E05-03-0193. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 154.Amano M, Nakayama M, Kaibuchi K. Rho-kinase/ROCK: a key regulator of the cytoskeleton and cell polarity. Cytoskeleton (Hoboken) 2010;67:545–554. doi: 10.1002/cm.20472. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 155.Ikebe M, Hartshorne DJ, Elzinga M. Phosphorylation of the 20,000-dalton light chain of smooth muscle myosin by the calcium-activated, phospholipid-dependent protein kinase. Phosphorylation sites and effects of phosphorylation. J Biol Chem. 1987;262:9569–9573. [PubMed] [Google Scholar]
- 156.Al-Sadi R, Ye D, Said HM, et al. IL-1beta-induced increase in intestinal epithelial tight junction permeability is mediated by MEKK-1 activation of canonical NF-kappaB pathway. Am J Pathol. 2010;177:2310–2322. doi: 10.2353/ajpath.2010.100371. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 157.Hopkins AM, Walsh SV, Verkade P, et al. Constitutive activation of Rho proteins by CNF-1 influences tight junction structure and epithelial barrier function. J Cell Sci. 2003;116:725–742. doi: 10.1242/jcs.00300. [DOI] [PubMed] [Google Scholar]
- 158.Samarin SN, Ivanov AI, Flatau G, et al. Rho/ Rho-associated kinase-II signaling mediates disassembly of epithelial apical junctions. Mol Biol Cell. 2007;18:3429–3439. doi: 10.1091/mbc.E07-04-0315. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 159.Troyanovsky S. Adherens junction assembly. Subcell Biochem. 2012;60:89–108. doi: 10.1007/978-94-007-4186-7_5. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 160.Nava P, Kamekura R, Nusrat A. Cleavage of transmembrane junction proteins and their role in regulating epithelial homeostasis. Tissue Barriers. 2013;1:e24783. doi: 10.4161/tisb.24783. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 161.David JM, Rajasekaran AK. Dishonorable discharge: the oncogenic roles of cleaved E-cadherin fragments. Cancer Res. 2012;72:2917–2923. doi: 10.1158/0008-5472.CAN-11-3498. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 162.Rodriguez-Feo JA, Puerto M, Fernandez-Mena C, et al. A new role for reticulon-4B/NOGO-B in the intestinal epithelial barrier function and inflammatory bowel disease. Am J Physiol Gastrointest Liver Physiol. 2015;308:G981–G993. doi: 10.1152/ajpgi.00309.2014. [DOI] [PubMed] [Google Scholar]
- 163.Yu M, Yang S, Qiu Y, et al. Par-3 modulates intestinal epithelial barrier function through regulating intracellular trafficking of occludin and myosin light chain phosphorylation. J Gastroenterol. 2015;50:1103–1113. doi: 10.1007/s00535-015-1066-z. [DOI] [PubMed] [Google Scholar]
- 164.Buckley CD, Gilroy DW, Serhan CN. Proresolving lipid mediators and mechanisms in the resolution of acute inflammation. Immunity. 2014;40:315–327. doi: 10.1016/j.immuni.2014.02.009. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 165.Leoni G, Alam A, Neumann PA, et al. Annexin A1, formyl peptide receptor, and NOX1 orchestrate epithelial repair. J Clin Invest. 2013;123:443–454. doi: 10.1172/JCI65831. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 166.Leoni G, Gripentrog J, Lord C, et al. Human neutrophil formyl peptide receptor phosphorylation and the mucosal inflammatory response. J Leukoc Biol. 2015;97:87–101. doi: 10.1189/jlb.4A0314-153R. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 167.Arita M, Yoshida M, Hong S, et al. Resolvin E1, an endogenous lipid mediator derived from omega-3 eicosapentaenoic acid, protects against 2,4,6-trinitrobenzene sulfonic acid-induced colitis. Proc Natl Acad Sci U S A. 2005;102:7671–7676. doi: 10.1073/pnas.0409271102. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 168.Campbell EL, Louis NA, Tomassetti SE, et al. Resolvin E1 promotes mucosal surface clearance of neutrophils: a new paradigm for inflammatory resolution. FASEB J. 2007;21:3162–3170. doi: 10.1096/fj.07-8473com. [DOI] [PubMed] [Google Scholar]
- 169.Fournier BM, Parkos CA. The role of neutrophils during intestinal inflammation. Mucosal Immunol. 2012;5:354–366. doi: 10.1038/mi.2012.24. [DOI] [PubMed] [Google Scholar]
- 170.Serhan CN, Chiang N, Van Dyke TE. Resolving inflammation: dual anti-inflammatory and pro-resolution lipid mediators. Nat Rev Immunol. 2008;8:349–361. doi: 10.1038/nri2294. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 171.Koch S, Capaldo CT, Hilgarth RS, et al. Protein kinase CK2 is a critical regulator of epithelial homeostasis in chronic intestinal inflammation. Mucosal Immunol. 2013;6:136–145. doi: 10.1038/mi.2012.57. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 172.Duncan JS, Turowec JP, Duncan KE, et al. A peptide-based target screen implicates the protein kinase CK2 in the global regulation of caspase signaling. Sci Signal. 2011;4:ra30. doi: 10.1126/scisignal.2001682. [DOI] [PubMed] [Google Scholar]
- 173.Raleigh DR, Boe DM, Yu D, et al. Occludin S408 phosphorylation regulates tight junction protein interactions and barrier function. J Cell Biol. 2011;193:565–582. doi: 10.1083/jcb.201010065. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 174.Escobar DJ, Desai R, Ishiyama N, et al. alpha-Catenin phosphorylation promotes intercellular adhesion through a dual-kinase mechanism. J Cell Sci. 2015;128:1150–1165. doi: 10.1242/jcs.163824. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 175.Serres M, Filhol O, Lickert H, et al. The disruption of adherens junctions is associated with a decrease of E-cadherin phosphorylation by protein kinase CK2. Exp Cell Res. 2000;257:255–264. doi: 10.1006/excr.2000.4895. [DOI] [PubMed] [Google Scholar]
- 176.Zindl CL, Lai JF, Lee YK, et al. IL-22-producing neutrophils contribute to antimicrobial defense and restitution of colonic epithelial integrity during colitis. Proc Natl Acad Sci U S A. 2013;110:12768–12773. doi: 10.1073/pnas.1300318110. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 177.Lee JS, Tato CM, Joyce-Shaikh B, et al. Interleukin-23-independent IL-17 production regulates intestinal epithelial permeability. Immunity. 2015;43:727–738. doi: 10.1016/j.immuni.2015.09.003. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 178.Maxwell JR, Zhang Y, Brown WA, et al. Differential Roles for Interleukin-23 and Interleukin-17 in Intestinal Immunoregulation. Immunity. 2015;43:739–750. doi: 10.1016/j.immuni.2015.08.019. [DOI] [PubMed] [Google Scholar]
- 179.Dudakov JA, Hanash AM, van den Brink MR. Interleukin-22: immunobiology and pathology. Annu Rev Immunol. 2015;33:747–785. doi: 10.1146/annurev-immunol-032414-112123. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 180.Shih VF, Cox J, Kljavin NM, et al. Homeostatic IL-23 receptor signaling limits Th17 response through IL-22-mediated containment of commensal microbiota. Proc Natl Acad Sci U S A. 2014;111:13942–13947. doi: 10.1073/pnas.1323852111. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 181.Schreiber F, Arasteh JM, Lawley TD. Pathogen resistance mediated by IL-22 signaling at the epithelial-microbiota interface. J Mol Biol. 2015;427:3676–3682. doi: 10.1016/j.jmb.2015.10.013. [DOI] [PubMed] [Google Scholar]
- 182.Backert I, Koralov SB, Wirtz S, et al. STAT3 activation in Th17 and Th22 cells controls IL-22-mediated epithelial host defense during infectious colitis. J Immunol. 2014;193:3779–3791. doi: 10.4049/jimmunol.1303076. [DOI] [PubMed] [Google Scholar]
- 183.Sugimoto K, Ogawa A, Mizoguchi E, et al. IL-22 ameliorates intestinal inflammation in a mouse model of ulcerative colitis. J Clin Invest. 2008;118:534–544. doi: 10.1172/JCI33194. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 184.Medina-Contreras O, Harusato A, Nishio H, et al. Cutting edge: IL-36 receptor promotes resolution of intestinal damage. J Immunol. 2016;196:34–38. doi: 10.4049/jimmunol.1501312. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 185.Scheibe K, Backert I, Wirtz S, et al. IL-36R signalling activates intestinal epithelial cells and fibroblasts and promotes mucosal healing in vivo. Gut. Jan 18; doi: 10.1136/gutjnl-2015-310374. pii: gutjnl-2015–310374. http://dx.doi.org/10.1136/gutjnl-2015-310374. [Epub ahead of print] [DOI] [PubMed]
- 186.Jarry A, Bossard C, Bou-Hanna C, et al. Mucosal IL-10 and TGF-beta play crucial roles in preventing LPS-driven, IFN-gamma-mediated epithelial damage in human colon explants. J Clin Invest. 2008;118:1132–1142. doi: 10.1172/JCI32140. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 187.Kominsky DJ, Campbell EL, Ehrentraut SF, et al. IFN-gamma-mediated induction of an apical IL-10 receptor on polarized intestinal epithelia. J Immunol. 2014;192:1267–1276. doi: 10.4049/jimmunol.1301757. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 188.Lu J, Philpott DJ, Saunders PR, et al. Epithelial ion transport and barrier abnormalities evoked by superantigen-activated immune cells are inhibited by interleukin-10 but not interleukin-4. J Pharmacol Exp Ther. 1998;287:128–136. [PubMed] [Google Scholar]
- 189.Madsen KL, Lewis SA, Tavernini MM, et al. Interleukin 10 prevents cytokine-induced disruption of T84 monolayer barrier integrity and limits chloride secretion. Gastroenterology. 1997;113:151–159. doi: 10.1016/s0016-5085(97)70090-8. [DOI] [PubMed] [Google Scholar]
- 190.Suenaert P, Maerten P, Van Assche G, et al. Effects of T cell-induced colonic inflammation on epithelial barrier function. Inflamm Bowel Dis. 2010;16:1322–1331. doi: 10.1002/ibd.21211. [DOI] [PubMed] [Google Scholar]
- 191.Kelly CJ, Zheng L, Campbell EL, et al. Crosstalk between microbiota-derived short-chain fatty acids and intestinal epithelial HIF augments tissue barrier function. Cell Host Microbe. 2015;17:662–671. doi: 10.1016/j.chom.2015.03.005. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 192.Ivanov II, Atarashi K, Manel N, et al. Induction of intestinal Th17 cells by segmented filamentous bacteria. Cell. 2009;139:485–498. doi: 10.1016/j.cell.2009.09.033. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 193.Yu LC, Wang JT, Wei SC, et al. Host-microbial interactions and regulation of intestinal epithelial barrier function: From physiology to pathology. World J Gastrointest Pathophysiol. 2012;3:27–43. doi: 10.4291/wjgp.v3.i1.27. [DOI] [PMC free article] [PubMed] [Google Scholar]