
Figure 1.
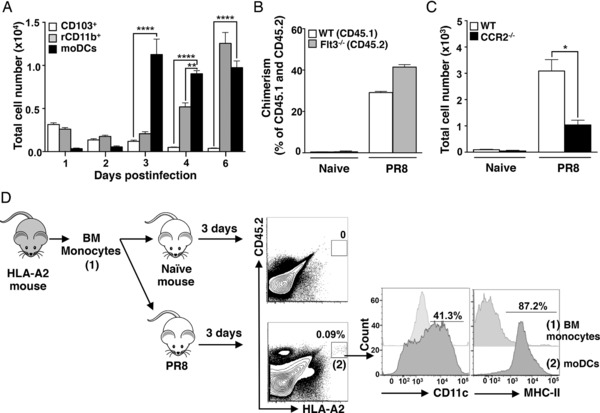
Influenza infection promotes recruitment of monocytes and triggers their differentiation into moDCs. (A) WT mice were infected intranasally with 250 PFU of PR8 and absolute numbers of CD103+ DCs, lung resident CD11b+ DCs (rCD11b) and moDCs were determined at the indicated time points (n = 3 mice per time point) in the lungs by flow cytometry. (B) WT/Flt3−/− mixed BM chimeric mice were infected intranasally with 250 PFU of PR8 and the frequency of moDCs was evaluated in the lungs 4 dpi by flow cytometry. The frequency of moDCs in the CD45.1 (WT) or CD45.2 (Flt3−/−) gates is shown. Data are shown as mean ± standard error of the mean of n = 4 mice. (C) WT and CCR2−/− mice were infected intranasally with 250 PFU of PR8. Absolute cell number of moDCs in the singlet population was determined at 4 dpi in the lungs. Data are shown as mean ± SEM of n = 4 mice. (A–C) Graphs depict one representative experiment of at least three experiments. (D) Monocytes were sorted as SSC‐Alow CD11c− MHC‐II− CD11b+ Ly6C+ cells from the BM of donor HLA‐A2+ transgenic mice. Purified monocytes were injected in naïve mice or in mice infected with PR8 for 3 days. Twenty‐four hours later, the phenotype of donor cells (0.09% of the singlet population and 3% of the CD11c+ MHC class IIhi cell population) was determined in the lungs by flow cytometry. Data shown are from a single experiment performed with n = 5 mice. Two independent experiments were performed. In all cases, data are shown as mean ± SEM. Asterisks represent statistical significance as follows: *p < 0.05; **p < 0.005; ****p < 0.0001 as assessed by one‐way ANOVA followed by Bonferroni's posttest.