

It is becoming increasingly evident that steady-state mRNA levels do not always reflect the cellular proteome composition. Pathways that regulate mRNA translation determine the repertoire of proteins being expressed. Deregulation of processes involved in translation control participate in the development and progression of different diseases including cancer.1 Tumoral cells modulate translational programming to increase the activity of components of the translation machinery. This boosts the overall protein synthesis capacity and favors the expression of specific oncogenic networks. Hence, therapeutic strategies that target components of the translation machinery to decrease protein synthesis rates and rewire translation programs hold promise in a broad range of cancer types.1 Various compounds have been shown to affect protein synthesis and are investigated as possible cancer therapeutics. Among these, we recently described that the HIV aspartyl protease inhibitors (HIV-PIs), a family of antiviral drugs, regulate pathways that modulate mRNA translation.2
The HIV-PIs were designed to specifically target the HIV protease. These safe and orally available molecules have been successfully used for more than 2 decades to decrease viral loads in infected patients. Intriguingly, early studies indicated that patients treated with these drugs had a significant reduction in HIV-associated tumors.3 Further studies have demonstrated that Nelfinavir (NFR), one of the first HIV-PIs to be approved by the FDA, showed promising anticancer properties in mice and in non-infected cancer patients. This led to phase I and phase II clinical trials that were designed to evaluate NFR repositioning in various cancer types.3 Nevertheless, the mechanisms underlying NFR and HIV-PIs anti-tumoral properties remained poorly understood.3
In a previous study we showed that NFR targets translation program by inducing the integrated stress response (ISR).4 This response is driven by the phosphorylation of the translation initiation factor eIF2α to decrease overall translation initiation and increase production of stress factors including the transcription factor ATF4. While the ISR is a key response that modulates cellular homeostasis, this pathway did not affect cellular viability in presence of the HIV-PIs. In contrast, we found that NFR-mediated toxicity relied on the activation of the kinase eEF2K2 (Fig. 1).
Figure 1.
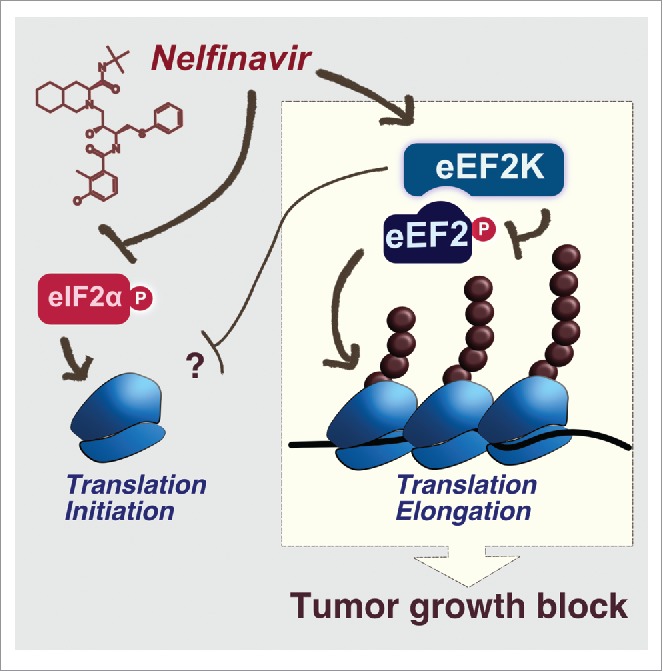
Translational control by Nelfinavir. Nelfinavir inhibits translation initiation by triggering the phosphorylation of eIF2α. Moreover, Nelfinavir promotes the activation of the kinase eEF2K leading to the phosphorylation of the elongation factor eEF2, thereby blocking its ability to promote translation elongation. In addition eEF2K activation may affect a yet to be defined step in the initiation process. EEF2K activation by Nelfinavir dampens tumor growth.
The eukaryotic elongation Factor 2 Kinase (eEF2K), belongs to a small group of proteins containing an α-kinase catalytic domain.5 It mainly functions as a negative modulator of protein synthesis through its ability to phosphorylate and inhibit its unique known target: the translation elongation factor eEF2. This factor mediates GTP-dependent tRNA-mRNA duplex translocation through the ribosome. Therefore its inhibition negatively impacts protein synthesis. The eEF2K pathway restores cellular homeostasis upon conditions of nutrient or energy depletion by decreasing translation rates at the stage of elongation. Within tumors, decreased protein synthesis by eEF2K activation has been proposed to contribute to tumor adaptation under nutrient depletion and stress conditions including hypoxia.6
In contrast to its possible oncogenic role, increased activation of eEF2K by treatment with rapamycin (an inhibitor of mTORC1), has been shown to dampen tumor growth in a model of colorectal cancer.7 In line with these findings we found that NFR and other HIV-PIs elicited a sustained eEF2K activation that decreased protein synthesis, impaired cell proliferation and promoted cell-death. Moreover, eEF2K deficiency within tumors impaired NFR-mediated anticancer activity in immunocompromised mice, indicating that pharmacological eEF2K activation drives a tumor intrinsic response that impairs tumor growth.2
The molecular mechanisms by which NFR triggers eEF2K activation are still unidentified. Main known pathways involved in eEF2K activation, including those downstream of rapamycin, were found to be dispensable, suggesting that NFR mode of action defines a new mechanism of eEF2K activation. In addition, NFR did not promote the expected increase of the polysomal fraction, but led to the accumulation of free ribosomes in an eEF2K dependent manner, indicating that eEF2K activation may affect other steps of the translation process beyond its role in regulating elongation. This possibility may reveal a new facet of eEF2K function and therefore needs to be addressed in details.
Another important question is to understand how eEF2K activation dampens tumor growth. One hypothesis is that tumor cells are particularly sensitive to the general decrease in mRNA translation. Because the expression of eEF2K and its activity have been reported to be increased in many tumors,6 it is possible that pharmacological eEF2K activation and subsequent decrease of mRNA translation, may exceed the bearable threshold of overall decrease in protein synthesis. Therefore, in stressed tissues, eEF2K activation originally aimed at reducing the energy consuming processes of mRNA translation, may exacerbate the toxicity of eEF2K activating compound such as NFR. It is also possible that, rather than its function in reducing global protein translation, eEF2K drives specific alteration of the repertoire of translated genes that may directly impact tumor growth. Whether eEF2K activation preferentially affects factors involved in survival and proliferation remains to be addressed. A global approach analyzing how eEF2K activation rewires the translational landscape may provide hints on how the pharmacological activation of this stress kinase affects oncogenic processes.
No doubt that these questions will spark a renewed interest in the therapeutic benefits of targeting translation programs in cancer.
Disclosure of potential conflicts of interest
The authors declare no conflicts of interest.
References
- [1].Bhat M, Robichaud N, Hulea L, Sonenberg N, Pelletier J, Topisirovic I. Targeting the translation machinery in cancer. Nat Rev Drug Discov 2015; 14:261-78; PMID:25743081; http://dx.doi.org/ 10.1038/nrd4505 [DOI] [PubMed] [Google Scholar]
- [2].De Gassart A, Demaria O, Panes R, Zaffalon L, Ryazanov AG, Gilliet M, Martinon F. Pharmacological eEF2K activation promotes cell death and inhibits cancer progression. EMBO Rep 2016; 17:1471-84; PMID:27572820; http://dx.doi.org/ 10.15252/embr.201642194 [DOI] [PMC free article] [PubMed] [Google Scholar]
- [3].Koltai T. Voltage-gated sodium channel as a target for metastatic risk reduction with re-purposed drugs. F1000Res 2015; 4:9; PMID:26097685; http://dx.doi.org/ 10.12688/f1000research.6789.1 [DOI] [PMC free article] [PubMed] [Google Scholar]
- [4].De Gassart A, Bujisic B, Zaffalon L, Decosterd LA, Di Micco A, Frera G, Tallant R, Martinon F. An inhibitor of HIV-1 protease modulates constitutive eIF2a dephosphorylation to trigger a specific integrated stress response. Proc Natl Acad Sci U S A 2016; 113:E117-26; PMID:26715744; http://dx.doi.org/ 10.1073/pnas.1514076113 [DOI] [PMC free article] [PubMed] [Google Scholar]
- [5].Kenney JW, Moore CE, Wang X, Proud CG. Eukaryotic elongation factor 2 kinase, an unusual enzyme with multiple roles. Adv Biol Regul 2014; 55:15-27; PMID:24853390; http://dx.doi.org/ 10.1016/j.jbior.2014.04.003 [DOI] [PubMed] [Google Scholar]
- [6].Leprivier G, Remke M, Rotblat B, Dubuc A, Mateo A-RF, Kool M, Agnihotri S, El-Naggar A, Yu B, Somasekharan SP, et al. . The eEF2 kinase confers resistance to nutrient deprivation by blocking translation elongation. Cell 2013; 153:1064-79; PMID:23706743; http://dx.doi.org/ 10.1016/j.cell.2013.04.055 [DOI] [PMC free article] [PubMed] [Google Scholar]
- [7].Faller WJ, Jackson TJ, Knight JRP, Ridgway RA, Jamieson T, Karim SA, Jones C, Radulescu S, Huels DJ, Myant KB, et al. . mTORC1-mediated translational elongation limits intestinal tumour initiation and growth. Nature 2015; 517:497-500; PMID:25383520; http://dx.doi.org/ 10.1038/nature13896 [DOI] [PMC free article] [PubMed] [Google Scholar]