

Abstract
Marine natural products with antibiotic activity have been a rich source of drug discovery; however, the emergence of antibiotic-resistant bacterial strains has turned attention towards the discovery of alternative innovative strategies to combat pathogens. In many pathogenic bacteria, the expression of virulence factors is under the regulation of quorum sensing (QS). QS inhibitors (QSIs) present a promising alternative or potential synergistic treatment since they disrupt the signaling pathway used for intra- and interspecies coordination of expression of virulence factors. This review covers the set of molecules showing QSI activity that were isolated from marine organisms, including plants (algae), animals (sponges, cnidarians, and bryozoans), and microorganisms (bacteria, fungi, and cyanobacteria). The compounds found and the methods used for their isolation are the emphasis of this review.
Keywords: quorum-sensing inhibitors, quorum quenching, marine natural products, antimicrobial resistance, N-acyl homoserine lactones
1. Introduction
Overuse of antibiotics has led to a decrease in the effectiveness of the drugs currently available to combat life-threatening and debilitating diseases. As a survival mechanism, many pathogenic microbes have developed mechanisms for resisting the toxic effect of antimicrobials. Antimicrobial resistance can be encoded by “resistance genes” and susceptible strains can become resistant either through mutations in existing genes or by acquiring “resistance genes” from another organism that is already resistant [1,2,3]. There is an urgent need to discover new antimicrobial compounds and identify new methods for disease prevention and treatment [4]. Different factors can be important to avoid the emergence of new resistant traits: (a) understanding the basic concepts of infection and effective treatment; (b) keeping adequate infection control measures in hospitals to prevent the emergence of nosocomial resistant strains [5,6,7]; and (c) discovering new drugs and/or novel inhibitory pathways for fighting systemic infections [8].
An increased understanding of the mechanism underlying bacterial pathogenesis and intercellular microbial communication has revealed potential alternative/complementary strategies to treat bacteria-mediated diseases [9,10]. Specifically, the strategy relates to interference of intercellular signaling to alter the pathogen’s ability to coordinate and invade their hosts, whether human, animal, or plant. Quorum sensing (QS) acts as a tool that synchronizes physiological activities of bacteria based on cell density and by means of genetic regulation [11]. In many cases, the responses elicited by QS signals contribute directly to pathogenesis through the coordinated production of virulence determinants, such as toxins, proteases, and other immune-evasive factors. Additionally, QS can contribute to behaviors that enable bacteria to resist antimicrobial compounds, such as biofilm development [12]. If the QS signal communication that coordinates these pathogenic behaviors is blocked, it is theorized that bacteria would lose their ability to mount an organized assault on the host and thus would be less able to form organized community structures that promote resistance to antibiotics [12,13,14]. Communication interference was not a human invention; rather, it exists naturally in the microbial world and is employed to gain an advantage over competitors [15]. Different types of QS communication systems rely on diverse small, secreted signaling molecules also known as autoinducers (AIs), which can act much like hormones in higher organisms, to initiate coordinated responses across a population. Autoinducers belong to different categories including, among others, the well-studied N-acylated homoserine lactones (AHLs), sometimes referred to as autoinducer-1 (AI-1), used by many Gram-negative bacteria (Figure 1); oligopeptide-based signals, used by Gram-positive bacteria; and a shared furanone-based system (autoinducer-2, AI-2), used by both Gram-negative and Gram-positive bacteria. There are also other QS signals that go beyond these classes, including Pseudomonas quinolone signal (PQS), diffusible signal factor (DSF), and autoinducer-3 (AI-3) [16].
Figure 1.
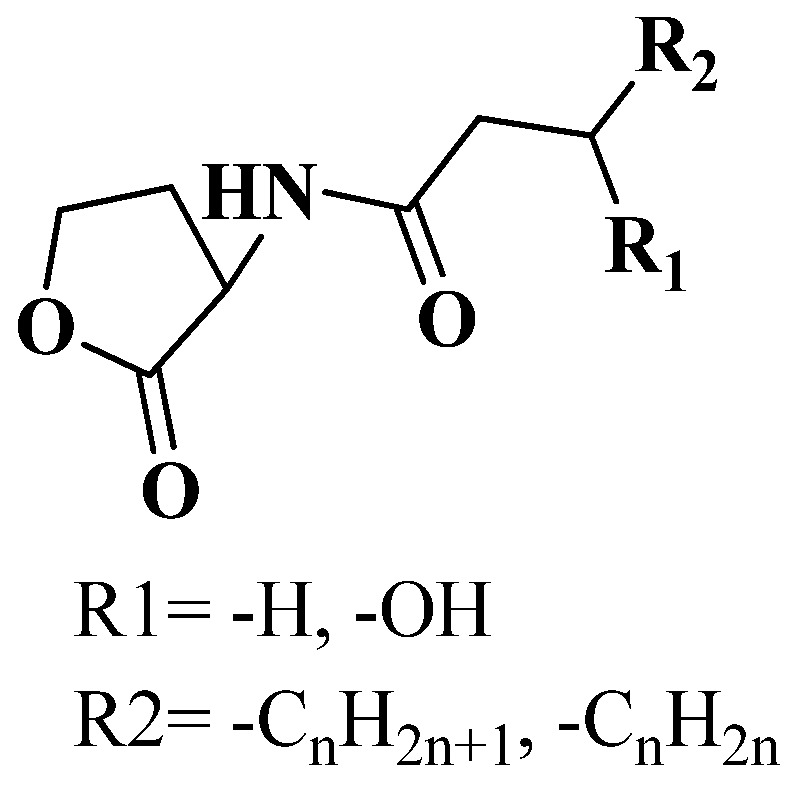
N-acyl homoserine lactones, found in many Gram-negative bacteria, vary by substitution at the C3 position (R1) and the length of the acyl chain (R2).
Despite the presence of diverse quorum-sensing network architectures, the key steps of signal supply and signal response are conserved in any QS system [12,17]; hence, QS inhibitors (QSI) can be categorized into two functional classes: signal-supply inhibitors and signal-response inhibitors, depending on whether a drug inhibits the function of signal generation or the response [18]. In this review, we focus on interference with AHL-QS systems. Although QSI has been shown to be an effective anti-infective strategy in different host–microbe systems, recent studies have suggested that QSI may also pose selective pressure. For example, the standard laboratory culture conditions using nutrient-rich media may not correctly mimic natural infection conditions, and resistance to QSI drugs may develop or not based on the specific environment the pathogen is experiencing [19]. It has also been proposed that QS blockade may increase the prevalence of more virulent traits among nosocomial infections [20]. For example, genetic knockouts in P. aeruginosa QS signal response show that signal-blind cheats are able to exploit a protease that is produced by signal-responsive individuals during well-mixed growth in both in vitro and animal models [21,22]. Further, resistance to one well-characterized QSI compounds, the brominated furanone C-30, was shown to be accomplished by the overexpression of the MexAB–OprM efflux pump [23]. While these and other recent studies provide evidence that QSI, as it is presently understood by the scientific community, is not the magic and final solution to antibiotic resistance in microbial pathogens, they provide motivation for more investigations into QSI compounds and mechanisms of action that can lead in the future to an efficient use of QSI as therapeutic treatment.
QSI metabolites, sometimes also referred to as quorum-quenching (QQ) compounds, have been recovered from extracts of marine algae (e.g., [24]), invertebrates (e.g., [25]), terrestrial plants (e.g., [26,27]) and bacteria (e.g., [28]). Also, the enzymatic inactivation of QS signals was reported in bacterial extracts (e.g., [29,30]), mammalian cells [31], and plants [27]. Although QS and QQ processes were both first discovered in marine organisms [32,33], information on QQ processes in the marine environment is scarce when compared to its terrestrial counterparts but some indirect evidence suggests QSI as a frequent phenomenon in the marine environment. Numerous reports are emerging that provide empirical data demonstrating QSI activity from various marine sources including sponges, algae, bryozoan, and coral-associated bacteria [16]. Various methods have been implemented to identify strains that produce QSI compounds, followed by lead molecule purification. QS biosensors, which are genetically modified strains that express reporter genes (e.g., lacZ, gfp or luxCDABEG) in response to the presence of specific QS signals [34], are also valuable tools for the identification of QSI molecules. Here, we provide the list of the most widely used AHL-based biosensors that were applied for bioassay-guided isolation and identification of QSI lead molecules from marine sources (Table 1). The aim of this review is to give an overview of marine-derived secondary metabolites with QSI activity that have been described to date using AHL-based biosensor assays. The compounds were categorized based on their source of isolation.
Table 1.
AHL-based biosensors used for the identification of active QSI molecules from the sea.
| Strain/Plasmid | QS System | Reporter System | Detection Range | References |
|---|---|---|---|---|
| Chromobacterium violaceum CV026 | CviI/R | Violacein pigment | C6-HSL, 3-oxo-C6-HSL, C8-HSL, 3-oxo-C8-HSL | [35,36,37,38] |
| Chromobacterium violaceum CV017 | CviI/R | Violacein pigment | 3-oxo-C6-HSL, C8-HSL, 3-oxo-C8-HSL | [39,40,41] |
| Escherichia coli pSB403 | LuxI/R (V. fisheri) | luxCDABE | 3-oxo-C6-HSL, C6-HSL, 3-oxo-C8-HSL, C8-HSL | [37,42] |
| Escherichia coli pSB536 | AhyI/R (A. hydrophyla) | luxCDABE | C4-HSL | [24] |
| Escherichia coli pSB401 | LuxI/R (V. fisheri) | luxCDABE | 3-oxo-C6-HSL, C6-HSL, 3-oxo-C8-HSL, C8-HSL | [39,40,43,44,45,46,47] |
| Escherichia coli pSB1075 | LasI/R (P. aeruginosa) | luxCDABE | 3-oxo-C12-HSL, 3-oxo-C10-HSL, C12-HSL | [39,40,41] |
| QSIS2 | LasI/R (P. aeruginosa) | luxCDABE | 3-oxo-C12-HSL, 3-oxo-C10-HSL, C12-HSL | [25] |
| Vibrio harveyi JMH 612 | LuxPQ (Vibrio harveyi) | luxQ | 3-OH-C4-HSL | [48,49] |
| Agrobacterium tumifaciencens pZLR4 | TraI/R (A. tumefaciens) | β-galactosidase | All 3-oxo-HSLs | [41,50,51] |
| Escherichia coli pKDT17 | LasI/R (P. aeruginosa) | β-galactosidase | 3-oxo-C12-HSL, C12-HSL, C10-HSL, 3-oxo-C10-HSL | |
| QSIS1 | LuxI/R (V. fisheri) | β-galactosidase | 3-oxo-C6-HSL, C6-HSL, C8-HSL, C10-HSL | [25] |
| pAS-C8 | CepI/R (B. cepacia) | gfp | C8-HSL | [42] |
| pKR-C12 | LasI/R (P. aeruginosa) | gfp | 3-oxo-C12-HSL, 3-oxo-C10-HSL | [42] |
| Escherichia coli JB525 | LuxI/R (V. fisheri) | gfp | 3-oxo-C6-HSL, C6-HSL, C8-HSL, C10-HSL | [35] |
| QSIS3 | LuxI/R (V. fisheri) | gfp | 3-oxo-C6-HSL, C6-HSL, C8-HSL, C10-HSL | [25] |
| Tn5-Las | LasI/R (P. aeruginosa) | gfp | 3-oxo-C12-HSL, 3-oxo-C10-HSL | [25] |
2. Bacterial AHL-Based Biosensors
An AHL-mediated QS system uses AHLs as signal molecules. AHLs consist of an invariant homoserine lactone (HSL) ring attached to an acyl chain with varying length (C4-C20) and various substituent (methylene, keto, or hydroxy groups) at the C3 position (Figure 1). AHLs are most commonly synthesized by members of the LuxI family of AHL synthases with a few exceptions such as Vibrio harveyi and Vibrio fischeri, which are synthesized by LuxM and AinS, respectively [52,53]. The identification of a large number of AHL QS systems was facilitated by several qualitative and quantitative methods to detect AHLs based on bacterial biosensors. These biosensors do not produce AHLs and contain a functional response-regulator (e.g., LuxR-type) protein cloned together with a cognate target promoter (often the promoter of the cognate AHL synthase gene), which positively regulates the transcription of a reporter gene (e.g., bioluminescence, β-galactosidase, green-fluorescent protein (gfp), and violacein pigment production) [34]. The same biosensors were later utilized also for the screening of QSI molecules. Selection of suitable biosensors plays a crucial role in screening and isolation of QSI compounds and depends on the type of study that is being carried out. The following section indicates some of the selective biosensors commonly employed for screening and identification of active QSI lead molecules based on their phenotypic expression.
2.1. Pigment-Based Biosensors
One of the most widely used biosensor in this category is based on Chromobacterium violaceum, a Gram-negative bacterium that produces the visible purple pigment violacein, whose production is under the regulation of the CviI/CviR QS system. McClean and collaborators developed an AHL-deficient and nonpigmented mutant of this species, the C. violaceum strain CV026, by mini-T5 mutagenesis in cviI and in a QS repressor locus (Smr mini-Tn5 HgrcviI::Tn5xylE Kmr) [54]. This mutant will appear white unless provided with exogenous cognate AHL, which will result in the production of violacein and turn purple. Therefore, strain CV026 can be used as a biosensor to detect the presence of a range of AHLs, as well as being useful for screens of QSI molecules via the addition of exogenous AHLs and the detection of reduced violet pigmentation in the presence of QSI compounds in the tested extract. This pigment-based biosensor has been used both qualitatively and quantitatively for QSI compound searching; for example, using the overlaid well diffusion assay [54] or the violacein quantification assay in a 96-well plate [55]. CV026 was also previously used with Thin Layer Chromatography (TLC) for QSI screens, yet its detection limit for AHLs, C4-C8 acyl side chain length enables the search for QSI compounds that specifically affect the response to these signals [56]. Another C. violaceum-based mutant strain obtained through transposon insertion (Smr mini-Tn5 Hgr) of the wild strain C. violaceum ATCC 31532, and termed CV017, which results in the overproduction of violacein [57], has also been employed for bioactive guided isolation of QSIs (Table 1).
2.2. Bioluminescence-Based Biosensor
Bioluminescence is a common reporter activity used to quantify gene expression at high sensitivity and over a large dynamic range in real time and non-destructively [58]. The lux genes essential for luminescence are arranged in a single operon designated as luxCDABE. The luxCDE genes encode for a fatty acid reductase complex involved in synthesis of the fatty aldehyde substrate for the luminescence reaction catalyzed by the luciferase luxAB subunits [59]. Although all the lux genes isolated so far were derived from Gram-negative bacteria, their functional properties can vary between bacterial species of origin. For example, the luxCDABE operon from Photorhabdus luminescens permits a greater flexibility and ease of use in Gram-negative bacteria than the luxAB or luxCDABE systems derived from Vibrio fischeri or Vibrio harveyi, due to its wide temperature activity dependency [60]. Among others, the two most widely used bioluminescence-based QS reporters are plasmids pSB401 and pSB1075. Plasmid pSB401 was constructed using the P. luminescens luxCDABE operon controlled by the PluxI gene together with the V. fischeri luxR DNA fragment, and when transformed in E. coli it emits luminescence in response to the exogenous addition of AHLs with medium (C6-C8) acyl side chain length. LasR-based reporter plasmid pSB1075 contains the lasR gene, and the promoter of lasI− controlling the expression for reporter operon luxCDABE and emits luminescence in response to AHLs with long (≥C10) acyl side chain. Another plasmid, called pSB403, was constructed with the same arrangement as pSB401, and the advantage of being cloned into a wider host range replicon so it can be harbored in several other Gram-negative bacteria other than E. coli [58]. Swift et al. [61] constructed a similar biosensor (pSB536), sensitive to C4-AHL, using the ahyR AHL sensor/regulator of Aeromonas hydrophyla and the cognate ahyI gene promoter fused to luxCDABE. A quite different case is the bioluminescence-based reporter biosensor QSIS2, which was specifically developed for QSI assays. It is based on lasI-rhlI double mutant harboring the gene promoter of lasB controlling the expression of sacB1, encoding an AHL-induced killing system. The bacterial cell also harbors the plasmid pSU2007, which encodes the constitutively expressed luxCDABE, thus giving rise to bioluminescence expression in cases where the biosensor growth is rescued by the presence of a QSI molecule [62]. Another biosensor is the V. harveyi JMH 612 double mutant (luxQ:cqs Svh) enabling it to detect bioluminescence at very low cell density; it can thus be used for density-dependent bioluminescence assay [63].
QSI assays using bioluminescence-based biosensors can be visualized quantitatively/qualitatively by using bioluminescence microscopy or can be measured quantitatively using a luminometer. The general principle behind QSI assays is similar (with a few exceptions e.g., QSIS2) for the biosensors mentioned above. For a quantitative analysis, overnight cultures of these biosensors are inoculated in fresh medium at low OD (e.g., 0.01) and supplemented with the relevant cognate AHL to stimulate QS-induced expression of the reporter gene. QSI test extracts or purified molecules are added to the same biosensor cultures and kept for incubation. The bioluminescence is recorded over time (usually every 30 min for 1–16 h), and the production of bioluminescence is measured and compared to the same culture kept in the absence of the QSI test extract or purified compound (solvent of QSI extract/compound is added as a control).
2.3. gfp-Based Biosensors
The green fluorescent protein (GFP) of the jellyfish Aequorea victoria is a useful tool for non-invasive, real-time detection of gene expression at the single-cell level without the addition of any chemical substrates. The gfp gene encodes for a 27 kD monomer having a complex spectrum with a major excitation peak at 395 nm, and a minor peak at 475 nm [64]. Fluorescence of GFP producing strains can be easily quantified using, for example, 96-well plate assays. GFP-fluorescence for AHL-QS discovery can also be detected by microscopy and TLC assays. The GFP-based AHL sensor plasmid pKR-C12 contains a lasB-gfp (ASV) translational fusion together with a Plac constitutively expressed lasR gene, which encodes for the LasR receptor protein cloned in the broad host-range plasmid pBBR1MCS-5 [65]. Another GFP-based AHL sensor is the plasmid pAS-C8, which is based on the QS system of Burkholderia cepacia and contains PcepI-gfp (ASV) fusion together with the cepR regulator gene placed under control of Plac. pAS-C8 biosensor is most sensitive to C8-HSL [65,66]. These biosensors can be used for qualitative and quantitative screens, where epifluorescence microscope detects green fluorescent cells as early as 15 min after the addition of its cognate AHL [67]. Biosensor E. coli JB525 has been used for quantitative estimation of QSI molecules. This biosensor is composed by E. coli harboring the gfp plasmid pJBA132. The plasmid expresses the LuxR AHL receptor of Vibrio fischeri, and once activated by exogenously provided AHLs, LuxR induces the expression of the translational fusion PluxI-gfp (ASV) [67]. The QSIS3 system was constructed in host E. coli and is based on Vibrio fischeri LuxR QS system. The npt (kanamycin resistance gene) and gfp genes are controlled by the temperature-sensitive cI repressor, which in turn is regulated by QS through PluxI. In the presence of a QSI molecule, de-repression of antibiotic resistance leads to growth of the biosensor strain, which in turn produces GFP fluorescence, as the promotorless gfp gene is incorporated immediately downstream of the npt gene [62]. Hentzer et al. constructed a QS reporter system harboring the lasB-gfp (ASV) fusion in a P. aeruginosa PAO1 Tn5-Las background where the expression of unstable Gfp (ASV) is regulated by the QS-controlled PlasB. Upon reaching quorum, the biosensor produces fluorescence, yet in the presence of an exogenous QSI compound the decrease in fluorescence expression will be proportional to the concentration and efficacy of the QS inhibitor [68].
2.4. β-Galactosidase-Based Biosensor
β-galactosidase (β-gal) is a well-characterized bacterial enzyme containing one tetramer with a large subunit size of 1023 amino acids (the monomer is 116 kDa). β-gal can efficiently accelerate the hydrolysis of various β-galactosides, and has been used extensively as an endpoint control for assessing variability in reporter protein activity. The substrate 5-bromo-4-chloro-3-indolyl galactoside (X-Gal), a galactose sugar with a glycosidic linkage to an indolyl molecule, has been used widely as a reporter substrate. It remains colorless until the chromophore is linked to the galactose; however, upon hydrolysis in the presence of the enzyme β-gal, it produces a blue insoluble derivative, which is used in the reporter assay to visualize the activity [69]. Of all the AHL biosensors constructed using this reporter gene, the Agrobacterium tumefaciens-based sensor is particularly well suited and often used for TLC analysis and QS signals. A. tumefaciens strain NT1 carrying plasmid pZLR4 is one of the commonly used biosensors. NT1 does not harbor the Ti plasmid and accordingly does not produce native AHLs. Plasmid pZLR4 contains traR and a fusion of lacZ (encoding for β-gal) to traG, which is expressed from a TraR-dependent promoter, thus producing a blue colony appearance in the presence of exogenous AHLs [70]. In contrast, a colorless colony appears in the presence of a QSI molecule in the extract. This sensor can also be used in overlay techniques, similarly to the C. violaceum CV026 biosensor (as previously described) [71]. Another E. coli plasmid sensor based on the β-gal reporter is pKDT17, which was constructed by adding the Plac-lasR sequence into plasmid pTS400, which contains a lasB::lacZ translational fusion. Rasmussen et al. developed a unique suicide-based reporter system, QSIS1, to identify QSI molecules. This system expresses the luxR gene and a translational fusion of PluxI with phlA (from S. liquefaciens MG1), the latter producing a toxic compound. When QS is active, the phlA is expressed, and the biosensor is killed. Accordingly, the system is used as a biosensor for QSI compounds, in the presence of such molecules, QS is inhibited, phlA is not expressed, and the QSIS1 is alive expressing lacZ and appearing as a blue zone surrounding the sample containing the QSI compounds [62].
3. Marine Microorganisms
Marine microbes are particularly attractive in terms of their potential application in pharmaceutical industry sector because they (a) have not been extensively exploited as their terrestrial counterparts; and (b) possess potent bioactive compounds in order to be effective in the diluted marine seawater environment [72]. It is estimated that less than 1% of potentially useful chemicals from marine environments has been screened so far, reflecting the very low percentage of microbial products originating from marine microorganisms [73]. The exploration of marine microbial secondary metabolites has led to the discovery of numerous biologically active compounds that possess antibiotic, antitumor, and other pharmacological activities which are currently being used for the treatment of various diseases in humans, animals, and plants [74,75]. Natural products with antibiotic activity that derive from bacteria or fungi are almost always products of secondary metabolic pathways [76].
3.1. QSI from Marine Bacteria
Halobacillus salinus C42 isolated from a seagrass sample collected in 2002 from Point Judith Salt Pond, South Kingstown, RI led to the isolation of two phenethylamide metabolites, N-(2′-phenylethyl) isobutyramide (1) and 2,3-methyl-N-(2′-phenylethyl)-butyramide (2) (Figure 2), with potential to inhibit QS. Theses metabolites were quantitatively assessed for QS inhibition using Chromobacterium violaceum CV026, Vibrio harveyi BB120 and Escherichia coli JB525 at a non-inhibitory concentration below 200 µg/mL. These compounds behaved as antagonists of bacterial QS by competing with N-acyl homoserine lactones for receptor binding [35]. From the same location, the analysis of another Gram-positive bacterium, Bacillus cereus D28, yielded a cyclic dipeptide, cyclo(l-Pro-l-Tyr) (3) (Figure 2), using bioassay guided fractionation. QSI activity was measured using disk diffusion assays with V. harveyi BB120 and C. violaceum ATCC 12472 [28].
Figure 2.

Structures of phenethylamides (1–2), cyclic dipeptides (3–7), tyrosol (8) and tyrosol acetate (9).
Marinobacter sp. SK-3, isolated from a microbial mat sample collected from wadi Muqshin in southeastern Oman, resulted in the isolation of four diketopiperazines (DKPs), cyclo(l-Pro-l-Phe) (4), cyclo(l-Pro-l-Leu) (5), cyclo(l-Pro-l-isoLeu) (6), and cyclo(l-Pro-d-Phe) (7) (Figure 2), with QSI properties when screened using C. violaceum CV017 and Escherichia coli-based QS reporters (pSB401 and pSB1075). 4 and 6 inhibited QS-dependent production of violacein by C. violaceum CV017. 4, 5, and 6 reduced the QS-dependent luminescence of the reporter E. coli pSB401 induced by 3-oxo-C6-HSL [39]. Cyclo(Pro-Leu) (5) was also identified in an active subfraction of the extract from Staphylococcus saprophyticus by GC-MS analysis with moderate QSI activity [77]. The chemical study of the organic extract of Oceanobacillus profundus isolated from the Caribbean soft coral Antillogorgia elisabethae, yielded compounds tyrosol (8) and tyrosol acetate (9) (Figure 2), as responsible for the QSI activity detected using C. violaceum ATCC 31532 [78].
3.2. QSI from Marine Fungi
Marine fungal strains are potent producers of polyketide-derived alkaloids, terpenes, peptides, and mixed biosynthesis compounds that represent chemical groups of secondary metabolites produced by fungi [79]. A fermentation broth of Penicillium sp. SCS-KFD08 associated with marine animal Sipunculus nudus from Haikou Bay, China resulted in the isolation of six QSI active compounds (10–15) (Figure 3), using C. violaceum CV026 as a biosensor. These compounds exhibited strong QSI activity at a dosage of 50 µg/well using the well diffusion assay. At a non-inhibitory concentration, compounds 10 and 15 strongly reduced the violacein production by up to 49.1% and 45.5%, respectively, at a concentration of 300 µM [36]. Kojic acid (16) (Figure 3), previously isolated from marine-derived Alternaria sp. from the surface of the marine green alga Ulva pertusa [80] inhibited QS-dependent violacein production of the reporter C. violaceum CV017 at MIC of 239.25 µM and reduced the bioluminescence of reporter E. coli pSB401 induced by 3-oxo-C6-HSL at concentrations above 36 μM [40]. Meleagrin (17) (Figure 3), a known bacterial enoyl-acyl carrier protein reductase (FabI) inhibitor from the marine fungus Penicillum chrysogenium [81], inhibited QS of the bacterial reporter C. violaceum CV017 with MIC of 138.42 µM [40].
Figure 3.

Structures of Aculene E (10), Penicitor B (11), aculene C (12), aculene D (13), aspergillumarins A (14), aspergillumarins B (15), kojic acid (16) and meleagrin (17).
3.3. QSI from Marine Cyanobacteria
Cyanobacteria are an ancient group of organisms well known for their prolific production of bioactive natural products [82,83]. Since 2013, there has been an increase in the number of new metabolites reported from cyanobacteria [84]. Metabolites from the well-studied genus Lyngbya have been examined for production of QSI inhibitory compounds in addition to many other bioactive screens. Extracts of L. majuscula collected at various sites in Florida yielded 8-epi-malyngamide C (18), malyngamide C (19) and malyngolide (20) and a new small cyclopropane-containing fatty acid, lyngbyoic acid (21) (Figure 4). Compounds 18 and 19 were able to reduce 3-oxo-C12-HSL induced signaling in a LasR-based QS reporter (pSB1075) at a concentration of 10 µM [85]. 20 was identified as a QS inhibitor using the pSB1075 reporter with an EC50 value of 12.2 ± 1.6 µM. It was also found to inhibit LasR-regulated phenotypic production of elastase by P. aeruginosa PAO1 with EC50 = 10.6 ± 1.8 µM [41]. Compound 21 antagonized 3-oxo-C12-HSL induced bioluminescence of the pSB1075-based sensor with an IC50 of approximately 100 µM. 21 also reduced production of QS regulated pyocyanin and elastase in wild-type P. aeruginosa [24]. Blennothrix cantharidosmum collected at Duke of York Island in Papua New Guinea in 2005 resulted in the isolation of tumonoic acids (22–25) (Figure 5), which inhibited bioluminescence in wild-type strain Vibrio harveyi, with compound 23 having the strongest activity, IC50 of 62 μM [84].
Figure 4.

Structures of 8-epi-malyngamide C (18), malyngamide C (19), malyngolide (20) and lyngbyoic acid (21).
Figure 5.

Structures of tumonoic acids (22–25) and honaucins A–C (26–28).
Leptolyngbya crossbyana led to the isolation of three γ-butyrolactones, honaucins A–C (26–28) (Figure 5), from corals collected on the Hawaiian coast. Honaucins A–C presented a dose-dependent QSI by reducing bioluminescence expression in Vibrio harveyi BB120, with IC50 values of 5.6, 17.6, and 14.6 μM, respectively. 26–28 also exhibited potent inhibition of lipopolysaccharide-stimulated nitric oxide production in the murine macrophage cell line RAW264.7 [86].
Pitinoic acid A (29) (Figure 6), was isolated from a Guamanian cyanobacterium as an inhibitor of QS in P. aeruginosa and significantly reduced the transcript levels of lasB and the pyocyanin biosynthetic member phzG1 at 1 mM and 100 μM, respectively. Additionally, it also reduced levels of LasB and pyocyanin in the culture supernatants as evaluated by an enzymatic assay for LasB and quantitative evaluation using UV absorbance for pyocyanin at 1 mM [87]. Two lipopeptides, microcolins A (30) and B (31) and a peptide lyngbyastatin 3 (32) (Figure 6), from Lyngbya majuscula inhibited the QS of the bacterial reporter C. violaceum CV017 with an MIC of 15.23 µM, 43.21 µM, and 12 µM, respectively [40]. Recently, lyngbic acid (33) (Figure 6), has been isolated from a polymicrobial disease consortium dominated by the filamentous cyanobacterium Roseofilum reptotaenium with strong QSI activity against Vibrio harveyi-based QS reporters [48].
Figure 6.

Structures of Pitinoic acid A (29) microcolins A (30) and B (31) and a peptide lyngbyastatin 3 (32) and lyngbic acid (33).
4. QSI from Marine Sponges
Sponges (phylum Porifera) are sessile marine filter feeders evolutionarily considered as the oldest animals and the richest natural product source. Up to 2013, approximately 4851 discovered compounds contributing to nearly 30% of all marine natural products belong to marine sponges [88]. In 2014 alone, 283 new compound structures reported from sponges enabled the phylum to regain its position as the dominant source of new bioactive metabolites [75]. Compounds range from terpene skeletons, mixed polyketide-peptide biogenesis molecules to complex carbohydrate-based metabolites [89,90,91,92,93]. Many of these metabolites have been tested for their possible role as lead compounds in drug research. Extracts from Luffariella variabilis led to the isolation of three sesterterpene metabolites, manoalide (34), manoalide monoacetate (35), and secomanoalide (36) (Figure 7), with strong QS inhibition properties based on the QS reporter system QSIS1 and QSIS2. In addition, the activities for compound 34–36 were also quantitatively tested using biosensor P. aeruginosa PAO1 Tn5-Las with IC50 values of 0.658 mM, 1.123 mM, and 1.110 mM, respectively [25].
Figure 7.

Structures of manoalide (34), manoalide monoacetate (35), and secomanoalide (36).
QSI screening studies on marine invertebrates (39 extracts) including 26 sponges, seven soft corals, five algae, and one zooanthid collected in the Colombian Caribbean Sea and the Brazilian Coasts led to the isolation of five furanosesterterpenes (37–41) (Figure 8), from the sponge Ircinia felix. Compound (7Z,13Z,18R,20Z)-felixinin acetate (37), (8Z,13Z,18R,20Z)-strobilinin acetate (38), (7E,13Z,18R,20Z)-felixinin acetate (39) and (8E,13Z,18R,20Z)-strobilinin acetate (40) was isolated with distinct configuration along with compound (7E,12E,18R,20Z)-variabilin acetate (41) using C. violaceum biosensor [43]. These compounds were previously reported from I. felix based on their antimicrobial properties [94,95]. Similarly, Saurav et al. [96] screened marine sponges from the Mediterranean and Red Sea for QSI activity using the QSIS1 biosensor and C. violaceum CV026 systems and observed, QSI as a common phenomenon within half of the sponge species tested. Four sponge species extracts were observed to inhibit QS activity; more precisely, two Red Sea sponges, Suberites clavatus and Negombata magnifica, and two Mediterranean Sea species, Ircinia variabilis and Sarcotragus sp. However, the lead molecules responsible for this activity have not been characterized but it has been reported that sponge genera Ircinia sp. and Sarcotragus sp., both belonging to the family Irciniidae, are a potential source of QSI active lead molecules [97]. In addition, the investigation of the crude extract of Leucetta chagosensis led to the isolation of isonaamine D (42) (Figure 8), with mild but dose-dependent activity on the Vibrio harveyi JMH 612 bioreporter [49].
Figure 8.

Structures of furanosesterterpenes (37–41) and isonaamine D (42).
Dobretsov et al. [40] screened a large set of natural products for QSI activity. Five compounds (43–47) (Figure 9), deriving from various sponges were able to inhibit QS of C. violaceum CV017. Hymenialdisin (43), an alkaloid originally isolated from the sponge Axinella verrucosa [98] inhibited QS dependent luminescence in reporter strain pSB401 and pSB1075 induced by 3-oxo-C6-HSL and 3-oxo-C12-HSL, respectively. Several other alkaloids, ageliferin (44) isolated from Agelas conifer, mauritamide B (45) isolated from A. nakamurai, midpacamide (46), isolated from A. mauritama [99] and 4-(4,5-dibromo-1-methyl-1H-pyrrole-2-carboxamido) butanoic acid (47) isolated from Agelas sp. [100] inhibited QS of the bacterial reporter C. violaceum CV017 with an MIC of 11.29, 36.76, 458.61 and 271.73 µM, respectively.
Figure 9.

Structures of Hymenialdisin (43), ageliferin (44), mauritamide B (45), midpacamide (46) and butanoic acid (47).
On the other hand, one terpene, (+)avarol (48) and seven alkaloids (49–55) (Figure 10), from distinct marine sponges inhibited the QS of C. violaceum CV017, along with antibiotic properties [40,101].
Figure 10.

Structures of (+)avarol (48) and seven alkaloids (49–55).
5. QSI from Marine Algae
Marine algae act as a major food source in marine environments and produce a variety of compounds that are beneficial to human health [102,103]. These compounds include polyunsaturated fatty acids and carotenoids as well as other compounds with antibiotic and antifungal activity [104,105,106]. Several polyunsaturated fatty acids are also being studied in relation to their potential anticancer activity and treatment of the symptoms of cystic fibrosis [75,107,108]. The first QS antagonist compounds discovered from macro-algal source were isolated from D. pulchra and Ahnfeltiopsis flabelliformis. They include halogenated furanones, (56) [44,109], betonicine (57), cis-betonicine (58), floridoside (59), and isethionic acid (60) (Figure 11). They display an inhibition activity on QS mechanisms mediated by C8-AHL lactone and the TraR transcriptional activator protein [38,50,51,110]. ICR-FT/MS analysis revealed the presence of 2-dodecanoyloxyethanesulfonate (61) (Figure 11), in Asparagopsis taxiformis, as the lead molecule responsible for QSI activity based on Serratia liquefaciens MG44 and C. violaceum CV026 bioassays. This is in line with previous reports showing that sulfur-containing synthetic AHL-analogues can inhibit QS, such as the case of N-(propylsulfanylacetyl)-l-homoserine lactone, and N-(pentylsulfanylacetyl)-l-homoserine lactone [37,111].
Figure 11.

Structures of halogenated furanones, (56), betonicine (57), cis-betonicine (58), floridoside (59), and isethionic acid (60) and 2-dodecanoyloxyethanesulfonate (61).
6. QSI from Cnidarians
Cnidaria is a large, diverse, and ecologically important group of marine invertebrates (including jellyfish, sea anemones, and corals) that is characterized by specialized cells called cnidocytes or nematocytes, utilized for prey capture. Over 3000 natural products have been described from this phylum, mostly in the last two decades [112,113]. The ability of cnidarians to produce powerful toxins and venoms has also been well documented [114]. In terms of QSI compounds, four cembranoids (62–65) (Figure 12), were isolated from octocoral Eunicea knighti with QSI property. QSI assays were carried out using P. putida IsoF wild strain, E. coli pSB401, and C. violaceum (ATCC 31532) biosensor [45,46]. Another three cembranoid epimers at C-8 (66–68) (Figure 12), were isolated from the Colombian Caribbean octocoral Pseudoplexaura flagellosa based on inhibition of the production of violacein pigment of C. violaceum (ATCC 31532) biosensor [47].
Figure 12.

Structures of cembranoids (62–68).
7. QSI from Bryozoa
Bryozoans, also known as sea mats or sea mosses, are found in both freshwater and marine environments. Natural products and biological activity have been reported from all the bryozoan orders constituting the marine clade [115]. By using the biosensors P. putida (pKR-C12), P. putida (pAS-C8), and E. coli (pSB403) the antagonistic effect on N-acyl-homoserine lactone-dependent QS systems was investigated on metabolites from North Sea bryozoan Flustra foliacea, leading to the isolation of two QSI compounds, 69 and 70, (Figure 13), active at concentrations of 20 μg/mL [42].
Figure 13.

Structures of two brominated alkaloids (69, 70).
8. Concluding Remarks
The utilization of QSI could be a promising strategy for antivirulence therapy as it is less prone to the development of resistance. Moreover, natural heritable variation in QS genes and their expression level may result in differences in fitness under conditions that are different from those in standard laboratory, and can thus pose selective pressure on bacteria to develop resistance. To date more than 70 marine-derived QSI lead molecules with diverse molecular entities have been discovered using AHL-mediated biosensor, and many more compounds are expected to be discovered in the near future. It was evident from previous reports that several alkaloids from sponges exhibit QSI properties and hence it is noteworthy to screen previously isolated alkaloids from sponges. For example, Neopetrosia carbonaria, a rich producer of quinolinic type alkaloids and other nitrogen-containing compounds with cytotoxic and antimicrobial activity [116,117], could also possess QSI properties. Similarly, various acridine-type polar alkaloids were isolated from Xestospongia cf. carbonaria and Xestospongia cf. exigua with anticancer activity [118]. Consequently, it could be hypothesized that these alkaloids might possess QSI activity of interest; however, it has not yet been demonstrated.
Acknowledgments
This study was supported by the Bi-lateral Italy–Israel R&D grant, MOST grant#10705-3 to Laura Steindler and Vittorio Venturi entitled: “A novel approach to fight antibiotic-resistant pathogens: acquisition of quorum sensing inhibitors from marine sponges” (from the Ministry of Science and Technology, Israel). The work was partially supported by the European Union’s Seventh Framework Program (FP7) 2007–2013 under Grant Agreement No. 311848 (Bluegenics). Kumar Saurav had a post-doctoral fellowship from the Israeli Council for Higher Education (VATAT) and the University of Haifa.
Conflicts of Interest
The authors declare no conflicts of interest.
References
- 1.Arias C.A., Murray B.E. Emergence and management of drug-resistant enterococcal infections. Expert Rev. Anti Infect. Ther. 2008;6:637–655. doi: 10.1586/14787210.6.5.637. [DOI] [PubMed] [Google Scholar]
- 2.Yong D., Toleman M.A., Giske C.G., Cho H.S., Sundman K., Lee K., Walsh T.R. Characterization of a new metallo-beta-lactamase gene, bla(NDM-1), and a novel erythromycin esterase gene carried on a unique genetic structure in Klebsiella pneumoniae sequence type 14 from India. Antimicrob. Agents Chemother. 2009;53:5046–5054. doi: 10.1128/AAC.00774-09. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 3.Furuya E.Y., Lowy F.D. Antimicrobial-resistant bacteria in the community setting. Nat. Rev. Microbiol. 2006;4:36–45. doi: 10.1038/nrmicro1325. [DOI] [PubMed] [Google Scholar]
- 4.Appelbaum P.C. 2012 and beyond: Potential for the start of a second pre-antibiotic era? J. Antimicrob. Chemother. 2012;67:2062–2068. doi: 10.1093/jac/dks213. [DOI] [PubMed] [Google Scholar]
- 5.Arias C.A., Murray B.E. The rise of the Enterococcus: Beyond vancomycin resistance. Nat. Rev. Microbiol. 2012;10:266–278. doi: 10.1038/nrmicro2761. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 6.Dijkshoorn L., Nemec A., Seifert H. An increasing threat in hospitals: Multidrug-resistant Acinetobacter baumannii. Nat. Rev. Microbiol. 2007;5:939–951. doi: 10.1038/nrmicro1789. [DOI] [PubMed] [Google Scholar]
- 7.Otto M. Staphylococcus epidermidis—The ‘accidental’ pathogen. Nat. Rev. Microbiol. 2009;7:555–567. doi: 10.1038/nrmicro2182. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 8.Martinez J.L., Baquero F. Interactions among strategies associated with bacterial infection: Pathogenicity, epidemicity, and antibiotic resistance. Clin. Microbiol. Rev. 2002;15:647–679. doi: 10.1128/CMR.15.4.647-679.2002. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 9.Hancock R.E.W., Nijnik A., Philpott D.J. Modulating immunity as a therapy for bacterial infections. Nat. Rev. Microboil. 2012;10:243–254. doi: 10.1038/nrmicro2745. [DOI] [PubMed] [Google Scholar]
- 10.Rasko D.A., Sperandio V. Anti-virulence strategies to combat bacteria-mediated disease. Nat. Rev. Drug Discov. 2010;9:117–128. doi: 10.1038/nrd3013. [DOI] [PubMed] [Google Scholar]
- 11.Winzer K., Hardie K.R., Williams P. Bacterial cell-to-cell communication: Sorry, can’t talk now—Gone to lunch! Curr. Opin. Microbiol. 2002;5:216–222. doi: 10.1016/S1369-5274(02)00304-1. [DOI] [PubMed] [Google Scholar]
- 12.Rutherford S.T., Bassler B.L. Bacterial Quorum Sensing: Its Role in Virulence and Possibilities for Its Control. Cold Spring Harb. Perspect. Med. 2012;2:a012427. doi: 10.1101/cshperspect.a012427. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 13.Bhardwaj A.K., Vinothkumar K., Rajpara N. Bacterial quorum sensing inhibitors: Attractive alternatives for control of infectious pathogens showing multiple drug resistance. Recent Pat. Anti Infect. Drug Discov. 2013;8:68–83. doi: 10.2174/1574891X11308010012. [DOI] [PubMed] [Google Scholar]
- 14.Fuqua C., Greenberg E.P. Listening in on bacteria: Acyl-homoserine lactone signalling. Nat. Rev. Mol. Cell Biol. 2002;3:685–695. doi: 10.1038/nrm907. [DOI] [PubMed] [Google Scholar]
- 15.Bhavsar A.P., Guttman J.A., Finlay B.B. Manipulation of host-cell pathways by bacterial pathogens. Nature. 2007;449:827–834. doi: 10.1038/nature06247. [DOI] [PubMed] [Google Scholar]
- 16.LaSarre B., Federle M.J. Exploiting Quorum Sensing To Confuse Bacterial Pathogens. Microbiol. Mol. Biol. Rev. 2013;77:73–111. doi: 10.1128/MMBR.00046-12. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 17.Smith J.M., Harper D. Animal Signals. OUP Oxford; New York, NY, USA: 2003. [Google Scholar]
- 18.Allen R.C., Popat R., Diggle S.P., Brown S.P. Targeting virulence: Can we make evolution-proof drugs? Nat. Rev. Microboil. 2014;12:300–308. doi: 10.1038/nrmicro3232. [DOI] [PubMed] [Google Scholar]
- 19.Defoirdt T., Boon N., Bossier P. Can Bacteria Evolve Resistance to Quorum Sensing Disruption? PLoS Pathog. 2010;6:e1000989. doi: 10.1371/journal.ppat.1000989. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 20.Kohler T., Perron G.G., Buckling A., van Delden C. Quorum sensing inhibition selects for virulence and cooperation in Pseudomonas aeruginosa. PLoS Pathog. 2010;6:e1000883. doi: 10.1371/journal.ppat.1000883. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 21.Wang M., Schaefer A.L., Dandekar A.A., Greenberg E.P. Quorum sensing and policing of Pseudomonas aeruginosa social cheaters. Proc. Natl. Acad. Sci. USA. 2015;112:2187–2191. doi: 10.1073/pnas.1500704112. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 22.Mellbye B., Schuster M. The sociomicrobiology of antivirulence drug resistance: A proof of concept. MBio. 2011;2:e00131. doi: 10.1128/mBio.00131-11. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 23.Maeda T., Garcia-Contreras R., Pu M., Sheng L., Garcia L.R., Tomas M., Wood T.K. Quorum quenching quandary: Resistance to antivirulence compounds. ISME J. 2012;6:493–501. doi: 10.1038/ismej.2011.122. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 24.Kwan J.C., Meickle T., Ladwa D., Teplitski M., Paul V., Luesch H. Lyngbyoic acid, a “tagged” fatty acid from a marine cyanobacterium, disrupts quorum sensing in Pseudomonas aeruginosa. Mol. Biosyst. 2011;7:1205–1216. doi: 10.1039/c0mb00180e. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 25.Skindersoe M.E., Ettinger-Epstein P., Rasmussen T.B., Bjarnsholt T., de Nys R., Givskov M. Quorum Sensing Antagonism from Marine Organisms. Mar. Biotechnol. 2008;10:56–63. doi: 10.1007/s10126-007-9036-y. [DOI] [PubMed] [Google Scholar]
- 26.Chong Y.M., Yin W.F., Ho C.Y., Mustafa M.R., Hadi A.H.A., Awang K., Narrima P., Koh C.-L., Appleton D.R., Chan K.-G. Malabaricone C from Myristica cinnamomea Exhibits Anti-Quorum Sensing Activity. J. Nat. Prod. 2011;74:2261–2264. doi: 10.1021/np100872k. [DOI] [PubMed] [Google Scholar]
- 27.Koh C.-L., Sam C.-K., Yin W.-F., Tan L.Y., Krishnan T., Chong Y.M., Chan K.-G. Plant-Derived Natural Products as Sources of Anti-Quorum Sensing Compounds. Sensors. 2013;13:6217–6228. doi: 10.3390/s130506217. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 28.Teasdale M.E., Donovan K.A., Forschner-Dancause S.R., Rowley D.C. Gram-Positive Marine Bacteria as a Potential Resource for the Discovery of Quorum Sensing Inhibitors. Mar. Biotechnol. 2011;13:722–732. doi: 10.1007/s10126-010-9334-7. [DOI] [PubMed] [Google Scholar]
- 29.Dong Y.H., Gusti A.R., Zhang Q., Xu J.L., Zhang L.H. Identification of quorum-quenching N-acyl homoserine lactonases from Bacillus species. Appl. Environ. Microbiol. 2002;68:1754–1759. doi: 10.1128/AEM.68.4.1754-1759.2002. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 30.Chen F., Gao Y., Chen X., Yu Z., Li X. Quorum Quenching Enzymes and Their Application in Degrading Signal Molecules to Block Quorum Sensing-Dependent Infection. Int. J. Mol. Sci. 2013;14:17477–17500. doi: 10.3390/ijms140917477. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 31.Yang F., Wang L.-H., Wang J., Dong Y.-H., Hu J.Y., Zhang L.-H. Quorum quenching enzyme activity is widely conserved in the sera of mammalian species. FEBS Lett. 2005;579:3713–3717. doi: 10.1016/j.febslet.2005.05.060. [DOI] [PubMed] [Google Scholar]
- 32.Givskov M., de Nys R., Manefield M., Gram L., Maximilien R., Eberl L., Molin S., Steinberg P.D., Kjelleberg S. Eukaryotic interference with homoserine lactone-mediated prokaryotic signalling. J. Bacteriol. 1996;178:6618–6622. doi: 10.1128/jb.178.22.6618-6622.1996. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 33.Nealson K.H., Platt T., Hastings J.W. Cellular Control of the Synthesis and Activity of the Bacterial Luminescent System. J. Bacteriol. 1970;104:313–322. doi: 10.1128/jb.104.1.313-322.1970. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 34.Steindler L., Venturi V. Detection of quorum-sensing N-acyl homoserine lactone signal molecules by bacterial biosensors. FEMS Microbiol. Lett. 2007;266:1–9. doi: 10.1111/j.1574-6968.2006.00501.x. [DOI] [PubMed] [Google Scholar]
- 35.Teasdale M.E., Liu J., Wallace J., Akhlaghi F., Rowley D.C. Secondary Metabolites Produced by the Marine Bacterium Halobacillus salinus That Inhibit Quorum Sensing-Controlled Phenotypes in Gram-Negative Bacteria. Appl. Environ. Microbiol. 2009;75:567–572. doi: 10.1128/AEM.00632-08. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 36.Kong F.D., Zhou L.M., Ma Q.Y., Huang S.Z., Wang P., Dai H.F., Zhao Y.X. Metabolites with Gram-negative bacteria quorum sensing inhibitory activity from the marine animal endogenic fungus Penicillium sp. SCS-KFD08. Arch. Pharm. Res. 2016;40:25–31. doi: 10.1007/s12272-016-0844-3. [DOI] [PubMed] [Google Scholar]
- 37.Jha B., Kavita K., Westphal J., Hartmann A., Schmitt-Kopplin P. Quorum Sensing Inhibition by Asparagopsis taxiformis, a Marine Macro Alga: Separation of the Compound that Interrupts Bacterial Communication. Mar. Drugs. 2013;11:253–265. doi: 10.3390/md11010253. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 38.Natrah F.M.I., Kenmegne M.M., Wiyoto W., Sorgeloos P., Bossier P., Defoirdt T. Effects of micro-algae commonly used in aquaculture on acyl-homoserine lactone quorum sensing. Aquaculture. 2011;317:53–57. doi: 10.1016/j.aquaculture.2011.04.038. [DOI] [Google Scholar]
- 39.Abed R.M.M., Dobretsov S., Al-Fori M., Gunasekera S.P., Sudesh K., Paul V.J. Quorum-sensing inhibitory compounds from extremophilic microorganisms isolated from a hypersaline cyanobacterial mat. J. Ind. Microbiol. Biotechnol. 2013;40:759–772. doi: 10.1007/s10295-013-1276-4. [DOI] [PubMed] [Google Scholar]
- 40.Dobretsov S., Teplitski M., Bayer M., Gunasekera S., Proksch P., Paul V.J. Inhibition of marine biofouling by bacterial quorum sensing inhibitors. Biofouling. 2011;27:893–905. doi: 10.1080/08927014.2011.609616. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 41.Dobretsov S., Teplitski M., Alagely A., Gunasekera S.P., Paul V.J. Malyngolide from the cyanobacterium Lyngbya majuscula interferes with quorum sensing circuitry. Environ. Microbiol. Rep. 2010;2:739–744. doi: 10.1111/j.1758-2229.2010.00169.x. [DOI] [PubMed] [Google Scholar]
- 42.Peters L., König G.M., Wright A.D., Pukall R., Stackebrandt E., Eberl L., Riedel K. Secondary Metabolites of Flustra foliacea and Their Influence on Bacteria. Appl. Environ. Microbiol. 2003;69:3469–3475. doi: 10.1128/AEM.69.6.3469-3475.2003. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 43.Quintana J., Brango-Vanegas J., Costa G.M., Castellanos L., Arévalo C., Duque C. Marine organisms as source of extracts to disrupt bacterial communication: Bioguided isolation and identification of quorum sensing inhibitors from Ircinia felix. Rev. Bras. Farmacogn. 2015;25:199–207. doi: 10.1016/j.bjp.2015.03.013. [DOI] [Google Scholar]
- 44.Manefield M., Welch M., Givskov M., Salmond G.P., Kjelleberg S. Halogenated furanones from the red alga, Delisea pulchra, inhibit carbapenem antibiotic synthesis and exoenzyme virulence factor production in the phytopathogen Erwinia carotovora. FEMS Microbiol. Lett. 2001;205:131–138. doi: 10.1111/j.1574-6968.2001.tb10936.x. [DOI] [PubMed] [Google Scholar]
- 45.Tello E., Castellanos L., Arevalo-Ferro C., Duque C. Cembranoid diterpenes from the Caribbean sea whip Eunicea knighti. J. Nat. Prod. 2009;72:1595–1602. doi: 10.1021/np9002492. [DOI] [PubMed] [Google Scholar]
- 46.Tello E., Castellanos L., Arévalo-Ferro C., Duque C. Disruption in Quorum-Sensing Systems and Bacterial Biofilm Inhibition by Cembranoid Diterpenes Isolated from the Octocoral Eunicea knighti. J. Nat. Prod. 2012;75:1637–1642. doi: 10.1021/np300313k. [DOI] [PubMed] [Google Scholar]
- 47.Tello E., Castellanos L., Arevalo-Ferro C., Rodríguez J., Jiménez C., Duque C. Absolute stereochemistry of antifouling cembranoid epimers at C-8 from the Caribbean octocoral Pseudoplexaura flagellosa. Revised structures of plexaurolones. Tetrahedron. 2011;67:9112–9121. doi: 10.1016/j.tet.2011.09.094. [DOI] [Google Scholar]
- 48.Meyer J.L., Gunasekera S.P., Scott R.M., Paul V.J., Teplitski M. Microbiome shifts and the inhibition of quorum sensing by Black Band Disease cyanobacteria. ISME J. 2016;10:1204–1216. doi: 10.1038/ismej.2015.184. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 49.Mai T., Tintillier F., Lucasson A., Moriou C., Bonno E., Petek S., Magré K., Al Mourabit A., Saulnier D., Debitus C. Quorum sensing inhibitors from Leucetta chagosensis Dendy, 1863. Lett. Appl. Microbiol. 2015;61:311–317. doi: 10.1111/lam.12461. [DOI] [PubMed] [Google Scholar]
- 50.Liu H.B., Koh K.P., Kim J.S., Seo Y., Park S. The effects of betonicine, floridoside, and isethionic acid from the red alga Ahnfeltiopsis flabelliformis on quorum-sensing activity. Biotechnol. Bioprocess Eng. 2008;13:458–463. doi: 10.1007/s12257-008-0145-x. [DOI] [Google Scholar]
- 51.Kim J.S., Kim Y.H., Seo Y.W., Park S. Quorum sensing inhibitors from the red alga, Ahnfeltiopsis flabelliformis. Biotechnol. Bioprocess. Eng. 2007;12:308–311. doi: 10.1007/BF02931109. [DOI] [Google Scholar]
- 52.Gilson L., Kuo A., Dunlap P.V. AinS and a new family of autoinducer synthesis proteins. J. Bacteriol. 1995;177:6946–6951. doi: 10.1128/jb.177.23.6946-6951.1995. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 53.Hanzelka B.L., Parsek M.R., Val D.L., Dunlap P.V., Cronan J.E., Greenberg E.P. Acylhomoserine Lactone Synthase Activity of the Vibrio fischeri AinS Protein. J. Bacteriol. 1999;181:5766–5770. doi: 10.1128/jb.181.18.5766-5770.1999. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 54.McClean K.H., Winson M.K., Fish L., Taylor A., Chhabra S.R., Camara M., Daykin M., Lamb J.H., Swift S., Bycroft B.W., et al. Quorum sensing and Chromobacterium violaceum: Exploitation of violacein production and inhibition for the detection of N-acylhomoserine lactones. Microbiology. 1997;143(Pt 12):3703–3711. doi: 10.1099/00221287-143-12-3703. [DOI] [PubMed] [Google Scholar]
- 55.Martinelli D., Grossmann G., Sequin U., Brandl H., Bachofen R. Effects of natural and chemically synthesized furanones on quorum sensing in Chromobacterium violaceum. BMC Microbiol. 2004;4:25. doi: 10.1186/1471-2180-4-25. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 56.Bandara M.B.K., Zhu H., Sankaridurg P.R., Willcox M.D.P. Salicylic Acid Reduces the Production of Several Potential Virulence Factors of Pseudomonas aeruginosa Associated with Microbial Keratitis. Investig. Ophthalmol. Vis. Sci. 2006;47:4453–4460. doi: 10.1167/iovs.06-0288. [DOI] [PubMed] [Google Scholar]
- 57.Chernin L.S., Winson M.K., Thompson J.M., Haran S., Bycroft B.W., Chet I., Williams P., Stewart G.S.A.B. Chitinolytic Activity in Chromobacterium violaceum: Substrate Analysis and Regulation by Quorum Sensing. J. Bacteriol. 1998;180:4435–4441. doi: 10.1128/jb.180.17.4435-4441.1998. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 58.Winson M.K., Swift S., Fish L., Throup J.P., Jorgensen F., Chhabra S.R., Bycroft B.W., Williams P., Stewart G.S. Construction and analysis of luxCDABE-based plasmid sensors for investigating N-acyl homoserine lactone-mediated quorum sensing. FEMS Microbiol. Lett. 1998;163:185–192. doi: 10.1111/j.1574-6968.1998.tb13044.x. [DOI] [PubMed] [Google Scholar]
- 59.Meighen E.A. Molecular biology of bacterial bioluminescence. Microbiol. Rev. 1991;55:123–142. doi: 10.1128/mr.55.1.123-142.1991. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 60.Meighen E.A., Szittner R.B. Multiple repetitive elements and organization of the lux operons of luminescent terrestrial bacteria. J. Bacteriol. 1992;174:5371–5381. doi: 10.1128/jb.174.16.5371-5381.1992. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 61.Swift S., Karlyshev A.V., Fish L., Durant E.L., Winson M.K., Chhabra S.R., Williams P., Macintyre S., Stewart G.S. Quorum sensing in Aeromonas hydrophila and Aeromonas salmonicida: Identification of the LuxRI homologs AhyRI and AsaRI and their cognate N-acylhomoserine lactone signal molecules. J. Bacteriol. 1997;179:5271–5281. doi: 10.1128/jb.179.17.5271-5281.1997. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 62.Rasmussen T.B., Bjarnsholt T., Skindersoe M.E., Hentzer M., Kristoffersen P., Kote M., Nielsen J., Eberl L., Givskov M. Screening for quorum-sensing inhibitors (QSI) by use of a novel genetic system, the QSI selector. J. Bacteriol. 2005;187:1799–1814. doi: 10.1128/JB.187.5.1799-1814.2005. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 63.Henke J.M., Bassler B.L. Three parallel quorum-sensing systems regulate gene expression in Vibrio harveyi. J. Bacteriol. 2004;186:6902–6914. doi: 10.1128/JB.186.20.6902-6914.2004. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 64.Tsien R.Y. The green fluorescent protein. Annu. Rev. Biochem. 1998;67:509–544. doi: 10.1146/annurev.biochem.67.1.509. [DOI] [PubMed] [Google Scholar]
- 65.Riedel K., Hentzer M., Geisenberger O., Huber B., Steidle A., Wu H., Hoiby N., Givskov M., Molin S., Eberl L. N-acylhomoserine-lactone-mediated communication between Pseudomonas aeruginosa and Burkholderia cepacia in mixed biofilms. Microbiology. 2001;147:3249–3262. doi: 10.1099/00221287-147-12-3249. [DOI] [PubMed] [Google Scholar]
- 66.Huber B., Riedel K., Hentzer M., Heydorn A., Gotschlich A., Givskov M., Molin S., Eberl L. The cep quorum-sensing system of Burkholderia cepacia H111 controls biofilm formation and swarming motility. Microbiology. 2001;147:2517–2528. doi: 10.1099/00221287-147-9-2517. [DOI] [PubMed] [Google Scholar]
- 67.Andersen J.B., Heydorn A., Hentzer M., Eberl L., Geisenberger O., Christensen B.B., Molin S., Givskov M. gfp-based N-acyl homoserine-lactone sensor systems for detection of bacterial communication. Appl. Environ. Microbiol. 2001;67:575–585. doi: 10.1128/AEM.67.2.575-585.2001. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 68.Hentzer M., Riedel K., Rasmussen T.B., Heydorn A., Andersen J.B., Parsek M.R., Rice S.A., Eberl L., Molin S., Hoiby N., et al. Inhibition of quorum sensing in Pseudomonas aeruginosa biofilm bacteria by a halogenated furanone compound. Microbiology. 2002;148:87–102. doi: 10.1099/00221287-148-1-87. [DOI] [PubMed] [Google Scholar]
- 69.Alam J., Cook J.L. Reporter genes: Application to the study of mammalian gene transcription. Anal. Biochem. 1990;188:245–254. doi: 10.1016/0003-2697(90)90601-5. [DOI] [PubMed] [Google Scholar]
- 70.Farrand S.K., Qin Y., Oger P. Quorum-sensing system of Agrobacterium plasmids: Analysis and utility. Methods Enzymol. 2002;358:452–484. doi: 10.1016/s0076-6879(02)58108-8. [DOI] [PubMed] [Google Scholar]
- 71.Shaw P.D., Ping G., Daly S.L., Cha C., Cronan J.E., Rinehart K.L., Farrand S.K. Detecting and characterizing N-acyl-homoserine lactone signal molecules by thin-layer chromatography. Proc. Natl. Acad. Sci. USA. 1997;94:6036–6041. doi: 10.1073/pnas.94.12.6036. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 72.Lu Y., Dong X., Liu S., Bie X. Characterization and Identification of a Novel Marine Streptomyces sp. Produced Antibacterial Substance. Mar. Biotechnol. 2009;11:717–724. doi: 10.1007/s10126-009-9186-1. [DOI] [PubMed] [Google Scholar]
- 73.Cragg G.M., Newman D.J. Natural products: A continuing source of novel drug leads. Biochim. Biophys. Acta. 2013;1830:3670–3695. doi: 10.1016/j.bbagen.2013.02.008. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 74.Liu X., Ashforth E., Ren B., Song F., Dai H., Liu M., Wang J., Xie Q., Zhang L. Bioprospecting microbial natural product libraries from the marine environment for drug discovery. J. Antibiot. 2010;63:415–422. doi: 10.1038/ja.2010.56. [DOI] [PubMed] [Google Scholar]
- 75.Blunt J.W., Copp B.R., Keyzers R.A., Munro M.H., Prinsep M.R. Marine natural products. Nat. Prod. Rep. 2016;33:382–431. doi: 10.1039/C5NP00156K. [DOI] [PubMed] [Google Scholar]
- 76.Saurav K., Kannabiran K. Diversity and Optimization of Process Parameters for the Growth of Streptomyces VITSVK9 spp İsoled From Bay of Bengal, İndia. J. Nat. Environ. Sci. 2010;1:56–65. [Google Scholar]
- 77.Li M., Huiru Z., Biting D., Yun J., Wei J., Kunming D. Study on the anti-quorum sensing activity of a marine bacterium Staphylococcus saprophyticus 108. BioTechnology Indian J. 2013;7:480–487. [Google Scholar]
- 78.Martínez-Matamoros D., Fonseca M.L., Duque C., Ramos F.A., Castellanos L. Screening of marine bacterial strains as source of quorum sensing inhibitors (QSI): First chemical study of Oceanobacillus profundus (RKHC-62B) VITAE. 2016;23:30–47. doi: 10.17533/udea.vitae.v23n1a04. [DOI] [Google Scholar]
- 79.Hasan S., Ansari M.I., Ahmad A., Mishra M. Major bioactive metabolites from marine fungi: A Review. Bioinformation. 2015;11:176–181. doi: 10.6026/97320630011176. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 80.Li X., Jeong J.H., Lee K.T., Rho J.R., Choi H.D., Kang J.S., Son B.W. Gamma-pyrone derivatives, kojic acid methyl ethers from a marine-derived fungus Alternaria [correction of Altenaria] sp. Arch. Pharm. Res. 2003;26:532–534. doi: 10.1007/BF02976876. [DOI] [PubMed] [Google Scholar]
- 81.Zheng C.J., Sohn M.-J., Lee S., Kim W.-G. Meleagrin, a New FabI Inhibitor from Penicillium chryosogenum with at Least One Additional Mode of Action. PLoS ONE. 2013;8:e78922. doi: 10.1371/journal.pone.0078922. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 82.Miller B., Friedman A.J., Choi H., Hogan J., McCammon J.A., Hook V., Gerwick W.H. The marine cyanobacterial metabolite gallinamide A is a potent and selective inhibitor of human cathepsin L. J. Nat. Prod. 2014;77:92–99. doi: 10.1021/np400727r. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 83.Kleigrewe K., Almaliti J., Tian I.Y., Kinnel R.B., Korobeynikov A., Monroe E.A., Duggan M., Di Marzo V., Sherman D.H., Dorrestein P.C., et al. Combining Mass Spectrometric Metabolic Profiling with Genomic Analysis: A Powerful Approach for Discovering Natural Products from Cyanobacteria. J. Nat. Prod. 2015;78:1671–1682. doi: 10.1021/acs.jnatprod.5b00301. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 84.Clark B.R., Engene N., Teasdale M.E., Rowley D.C., Matainaho T., Valeriote F.A., Gerwick W.H. Natural products chemistry and taxonomy of the marine cyanobacterium Blennothrix cantharidosmum. J. Nat. Prod. 2008;71:1530–1537. doi: 10.1021/np800088a. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 85.Kwan J.C., Teplitski M., Gunasekera S.P., Paul V.J., Luesch H. Isolation and biological evaluation of 8-epi-malyngamide C from the Floridian marine cyanobacterium Lyngbya majuscula. J. Nat. Prod. 2010;73:463–466. doi: 10.1021/np900614n. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 86.Choi H., Mascuch S.J., Villa F.A., Byrum T., Teasdale M.E., Smith J.E., Preskitt L.B., Rowley D.C., Gerwick L., Gerwick W.H. Honaucins A–C, Potent Inhibitors of Eukaryotic Inflammation and Bacterial Quorum Sensing: Synthetic Derivatives and Structure-Activity Relationships. Chem. Biol. 2012;19:589–598. doi: 10.1016/j.chembiol.2012.03.014. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 87.Montaser R., Paul V.J., Luesch H. Modular Strategies for Structure and Function Employed by Marine Cyanobacteria: Characterization and Synthesis of Pitinoic Acids. Org. Lett. 2013;15:4050–4053. doi: 10.1021/ol401396u. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 88.Mehbub M.F., Lei J., Franco C., Zhang W. Marine Sponge Derived Natural Products between 2001 and 2010: Trends and Opportunities for Discovery of Bioactives. Mar. Drugs. 2014;12:4539–4577. doi: 10.3390/md12084539. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 89.Costantino V., Fattorusso E., Mangoni A., Perinu C., Cirino G., De Gruttola L., Roviezzo F. Tedanol: A potent anti-inflammatory ent-pimarane diterpene from the Caribbean Sponge Tedania ignis. Bioorg. Med. Chem. 2009;17:7542–7547. doi: 10.1016/j.bmc.2009.09.010. [DOI] [PubMed] [Google Scholar]
- 90.Teta R., Irollo E., Della Sala G., Pirozzi G., Mangoni A., Costantino V. Smenamides A and B, chlorinated peptide/polyketide hybrids containing a dolapyrrolidinone unit from the Caribbean sponge Smenospongia aurea. Evaluation of their role as leads in antitumor drug research. Mar. Drugs. 2013;11:4451–4463. doi: 10.3390/md11114451. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 91.Esposito G., Teta R., Miceli R., Ceccarelli L.S., Della Sala G., Camerlingo R., Irollo E., Mangoni A., Pirozzi G., Costantino V. Isolation and Assessment of the in Vitro Anti-Tumor Activity of Smenothiazole A and B, Chlorinated Thiazole-Containing Peptide/Polyketides from the Caribbean Sponge, Smenospongia aurea. Mar. Drugs. 2015;13:444–459. doi: 10.3390/md13010444. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 92.Costantino V., Fattorusso E., Imperatore C., Mangoni A. Glycolipids from sponges. 20. J-Coupling analysis for stereochemical assignments in furanosides: Structure elucidation of vesparioside B, a glycosphingolipid from the marine sponge Spheciospongia vesparia. J. Org. Chem. 2008;73:6158–6165. doi: 10.1021/jo800837k. [DOI] [PubMed] [Google Scholar]
- 93.Costantino V., Fattorusso E., Imperatore C., Mangoni A. Ectyoceramide, the First Natural Hexofuranosylceramide from the Marine Sponge Ectyoplasia ferox. Eur. J. Org. Chem. 2003;2003:1433–1437. doi: 10.1002/ejoc.200390202. [DOI] [Google Scholar]
- 94.Martinez A., Duque C., Hara N., Fujimoto Y. Variabilin 11-Methyloctadecanoate, A Branched-Chain Fatty Acid Ester of Furanosesterterpene Tetronic Acid, from the Sponge Ircinia Felix. Nat. Prod. Lett. 1995;6:281–284. doi: 10.1080/10575639508043171. [DOI] [Google Scholar]
- 95.Martinez A., Duque C., Sato N., Fujimoto Y. (8Z,13Z,20Z)-Strobilinin and (7Z,13Z,20Z)-Felixinin: New Furanosesterterpene Tetronic Acids from Marine Sponges of the Genus Ircinia. Chem. Pharm. Bull. 1997;45:181–184. doi: 10.1248/cpb.45.181. [DOI] [Google Scholar]
- 96.Saurav K., Bar-Shalom R., Haber M., Burgsdorf I., Oliviero G., Costantino V., Morgenstern D., Steindler L. In Search of Alternative Antibiotic Drugs: Quorum-Quenching Activity in Sponges and their Bacterial Isolates. Front. Microbiol. 2016;7:416. doi: 10.3389/fmicb.2016.00416. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 97.Hardoim C.C., Costa R. Microbial Communities and Bioactive Compounds in Marine Sponges of the Family Irciniidae—A Review. Mar. Drugs. 2014;12:5089–5122. doi: 10.3390/md12105089. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 98.Supriyono A., Schwarz B., Wray V., Witte L., Muller W.E., van Soest R., Sumaryono W., Proksch P. Bioactive alkaloids from the tropical marine sponge Axinella carteri. Z. Naturforsch. C. 1995;50:669–674. doi: 10.1515/znc-1995-9-1012. [DOI] [PubMed] [Google Scholar]
- 99.Hertiani T., Edrada-Ebel R., Ortlepp S., van Soest R.W., de Voogd N.J., Wray V., Hentschel U., Kozytska S., Muller W.E., Proksch P. From anti-fouling to biofilm inhibition: New cytotoxic secondary metabolites from two Indonesian Agelas sponges. Bioorg. Med. Chem. 2010;18:1297–1311. doi: 10.1016/j.bmc.2009.12.028. [DOI] [PubMed] [Google Scholar]
- 100.Hertiani T. New hope on drug leads development from deep ocean: Halogenated alkaloids of Agelas sponge. Indones. J. Pharm. 2014;25:199–222. doi: 10.14499/indonesianjpharm25iss4pp199. [DOI] [Google Scholar]
- 101.Ebel R., Brenzinger M., Kunze A., Gross H.J., Proksch P. Wound Activation of Protoxins in Marine Sponge Aplysina aerophoba. J. Chem. Ecol. 1997;23:1451–1462. doi: 10.1023/B:JOEC.0000006475.10310.3a. [DOI] [Google Scholar]
- 102.Loïc G.C., Leonel P. Marine Algae. CRC Press; Boca Raton, FL, USA: 2014. Review of Marine Algae as Source of Bioactive Metabolites: A Marine Biotechnology Approach; pp. 195–227. [Google Scholar]
- 103.El Gamal A.A. Biological importance of marine algae. Saudi Pharm. J. 2010;18:1–25. doi: 10.1016/j.jsps.2009.12.001. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 104.Pérez M.J., Falqué E., Domínguez H. Antimicrobial Action of Compounds from Marine Seaweed. Mar. Drugs. 2016;14:52. doi: 10.3390/md14030052. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 105.Michalak I., Chojnacka K. Algae as production systems of bioactive compounds. Eng. Life Sci. 2015;15:160–176. doi: 10.1002/elsc.201400191. [DOI] [Google Scholar]
- 106.Lee J.-C., Hou M.-F., Huang H.-W., Chang F.-R., Yeh C.-C., Tang J.-Y., Chang H.-W. Marine algal natural products with anti-oxidative, anti-inflammatory, and anti-cancer properties. Cancer Cell Int. 2013;13:55. doi: 10.1186/1475-2867-13-55. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 107.Pereira H., Barreira L., Figueiredo F., Custódio L., Vizetto-Duarte C., Polo C., Rešek E., Engelen A., Varela J. Polyunsaturated Fatty Acids of Marine Macroalgae: Potential for Nutritional and Pharmaceutical Applications. Mar. Drugs. 2012;10:1920–1935. doi: 10.3390/md10091920. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 108.Janczyk W., Socha P., Lebensztejn D., Wierzbicka A., Mazur A., Neuhoff-Murawska J., Matusik P. Omega-3 fatty acids for treatment of non-alcoholic fatty liver disease: Design and rationale of randomized controlled trial. BMC Pediatr. 2013;13:85. doi: 10.1186/1471-2431-13-85. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 109.Dworjanyn A.S., De Nys R., Steinberg D.P. Localisation and surface quantification of secondary metabolites in the red alga Delisea pulchra. Mar. Biol. 1999;133:727–736. doi: 10.1007/s002270050514. [DOI] [Google Scholar]
- 110.Delago A., Mandabi A., Meijler M.M. Natural Quorum Sensing Inhibitors—Small Molecules, Big Messages. Isr. J. Chem. 2016;56:310–320. doi: 10.1002/ijch.201500052. [DOI] [Google Scholar]
- 111.Koch B., Liljefors T., Persson T., Nielsen J., Kjelleberg S., Givskov M. The LuxR receptor: The sites of interaction with quorum-sensing signals and inhibitors. Microbiology. 2005;151:3589–3602. doi: 10.1099/mic.0.27954-0. [DOI] [PubMed] [Google Scholar]
- 112.Rocha J., Peixe L., Gomes N.C.M., Calado R. Cnidarians as a Source of New Marine Bioactive Compounds—An Overview of the Last Decade and Future Steps for Bioprospecting. Mar. Drugs. 2011;9:1860–1886. doi: 10.3390/md9101860. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 113.Daly M., Brugler M.R., Cartwright P., Collins A.G., Dawson M.N., Fautin D.G., France S.C., McFadden C.S., Opresko D.M., Rodriguez E., et al. The phylum Cnidaria: A review of phylogenetic patterns and diversity 300 years after Linnaeus. Zootaxa. 2007;1668:127–182. [Google Scholar]
- 114.Turk T., Kem W.R. The phylum Cnidaria and investigations of its toxins and venoms until 1990. Toxicon. 2009;54:1031–1037. doi: 10.1016/j.toxicon.2009.06.031. [DOI] [PubMed] [Google Scholar]
- 115.Sharp J.H., Winson M.K., Porter J.S. Bryozoan metabolites: An ecological perspective. Nat. Prod. Rep. 2007;24:659–673. doi: 10.1039/b617546e. [DOI] [PubMed] [Google Scholar]
- 116.Nakao Y., Shiroiwa T., Murayama S., Matsunaga S., Goto Y., Matsumoto Y., Fusetani N. Identification of Renieramycin A as an Antileishmanial Substance in a Marine Sponge Neopetrosia sp. Mar. Drugs. 2004;2:55–62. doi: 10.3390/md202055. [DOI] [Google Scholar]
- 117.De Almeida Leone P., Carroll A.R., Towerzey L., King G., McArdle B.M., Kern G., Fisher S., Hooper J.N.A., Quinn R.J. Exiguaquinol: A Novel Pentacyclic Hydroquinone from Neopetrosia exigua that Inhibits Helicobacter pylori MurI. Org. Lett. 2008;10:2585–2588. doi: 10.1021/ol800898z. [DOI] [PubMed] [Google Scholar]
- 118.Thale Z., Johnson T., Tenney K., Wenzel P.J., Lobkovsky E., Clardy J., Media J., Pietraszkiewicz H., Valeriote F.A., Crews P. Structures and Cytotoxic Properties of Sponge-Derived Bisannulated Acridines. J. Org. Chem. 2002;67:9384–9391. doi: 10.1021/jo026459o. [DOI] [PubMed] [Google Scholar]