

Abstract
Abstract
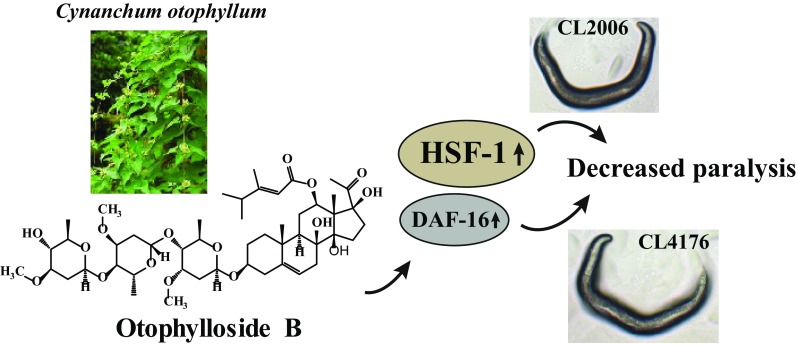
Alzheimer’s disease (AD) is a major public health concern worldwide and the few drugs currently available only treat the symptoms. Hence, there is a strong need to find more effective anti-AD agents. Cynanchum otophyllum is a traditional Chinese medicine for treating epilepsy, and otophylloside B (Ot B), isolated from C. otophyllum, is the essential active component. Having previously identified anti-aging effects of Ot B, we evaluated Ot B for AD prevention in C. elegans models of AD and found that Ot B extended lifespan, increased heat stress-resistance, delayed body paralysis, and increased the chemotaxis response. Collectively, these results indicated that Ot B protects against Aβ toxicity. Further mechanistic studies revealed that Ot B decreased Aβ deposition by decreasing the expression of Aβ at the mRNA level. Genetic analyses showed that Ot B mediated its effects by increasing the activity of heat shock transcription factor (HSF) by upregulating the expression of hsf-1 and its target genes, hsp-12.6, hsp-16.2 and hsp-70. Ot B also increased the expression of sod-3 by partially activating DAF-16, while SKN-1 was not essential in Ot B-mediated protection against Aβ toxicity.
Graphical Abstract
Electronic supplementary material
The online version of this article (doi:10.1007/s13659-017-0122-1) contains supplementary material, which is available to authorized users.
Keywords: Alzheimer’s disease, Caenorhabditis elegans, Otophylloside B, β-Amyloid peptide, HSF-1
Introduction
Alzheimer’s disease (AD) is a neurodegenerative disorder that is strongly related to aging. As the number one cause of senile dementia, it is now becoming a major public health concern around the world [1]. Paradoxically, there are only a few drugs approved for the AD treatment, and these drugs only treat the symptoms. To date, there are still no disease-modifying drugs available, and there is a strong need to find more effective anti-AD drugs [2].
Although the etiology of AD remains unclear, a growing body of evidence implicates that the extracellular senile plaques that result from accumulation of β-amyloid (Aβ) and intracellular tau protein tangles are key histopathological hallmarks of Alzheimer’s disease (AD), and that the neurotoxicity of Aβ may play a central role in the pathogenesis of AD [3, 4].
Because of its short lifespan and amenability to genetic manipulation, Caenorhabditis elegans models that mimic human disease have been extensively used to study the disease mechanism and to screen potential drugs [5]. To study the neurotoxicity of Aβ, multiple transgenic C. elegans strains expressing the human Aβ1–42 peptides in either neurons or muscle cells have been constructed. For example, CL2006 has muscle-specific expression of Aβ, leading to a progressive paralysis that starts in adulthood. CL4176 expresses Aβ in muscle cells in temperature-sensitive manner [6], while CL2355 expresses Aβ in the neurons, which may more accurately represent the amyloid induced toxicity seen in AD [7].
Qingyangshen (Cynanchum otophyllum) is a traditional Chinese medicine, whose root is used for the treatment of epilepsy, rheumatic pain, kidney weakness and muscle injuries [8, 9]. Otophylloside B (Ot B) is a C-21 steroidal glycoside, the essential active ingredient of Qingyangshen. Ot B has been shown to inhibit the seizure-like locomotor activity of zebrafish [10] and extends the lifespan of wild type worms [11]. Since nutraceuticals with pro-longevity properties often have the potential to delay the onset of AD [12–16], we are wondering if Ot B could delay the Aβ-induced pathological behavior in C. elegans.
Here, we used several AD transgenic C. elegans models to evaluate the potential of Ot B for the prevention of AD and to determine its molecular mechanism of action. Our results showed that Ot B extended the lifespan, improved the heat resistance, dramatically improved Aβ-induced pathological behavior, for example, delayed the progression of body paralysis, and improved chemotaxis response. These results indicated that Ot B played a protective role against Aβ toxicity. Further results showed that Ot B reduced the accumulation of Aβ, probably by increasing the gene expression of several heat shock proteins (HSP).
Results and Discussion
Ot B Extends Lifespan and Increases Heat Stress Resistance in C. elegans with Muscle-Specific Expression of Aβ
Treatment of CL2006 worms having muscle-specific expression of Aβ with 50 μM of Ot B caused a significant increase in their lifespan compared with controls (p < 0.005; Fig. 1b, Supplementary Table 1). We also measured the effect of Ot B on heat stress resistance and heat recovery in CL2006 worms. Ot B treatment suppressed the lethality of heat stress in heat resistance experiments and heat resistance recovery experiments (p < 0.005; Fig. 1c, d, Supplementary Table 2). Together, these results indicated that Ot B slows aging and delays age-related degeneration in C. elegans with muscle-specific expression of Aβ.
Fig. 1.
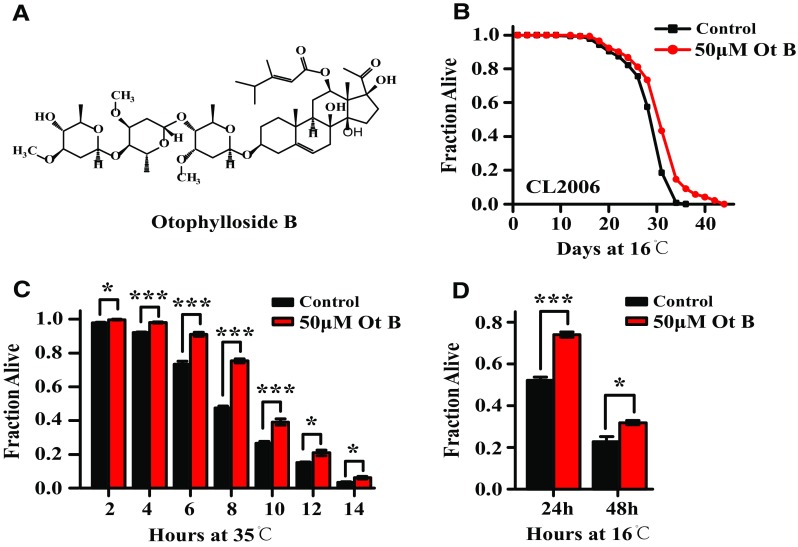
Otophylloside (Ot B) extended lifespan and improved thermo-tolerance in a C. elegans model of AD. a Chemical structure of Ot B. b Survival curves of CL2006 animals treated with vehicle or 50 μM of Ot B. c Survival percentage of CL2006 animals in heat resistance experiments. Heat resistance experiments were carried out at 35 °C and calculated by 2 h. d Survival percentage of CL2006 animals in heat resistance recovery experiment. The experiments were carried out at 35 °C for 7 h, then transferred to 16 °C and the number of dead worms was measured after 24 and 48 h. All the assays were carried out in triplicate, and at least three independent trials were performed. The columns showed the mean value of three independent experiments with error bars representing SEM. *** represents p < 0.001, * represents p < 0.05, calculated using two-tailed t test. Statistical details and repetition of this experiment are summarized in Table S1, S2 (Supplementary information)
Ot B Delays the Progression of Body Paralysis, and Improves Chemotaxis Response
Paralysis is an apparent symptom of AD, and in C. elegans models of AD, it is a measurable phenotype that is considered as a result of Aβ toxicity [17]. Our paralysis assay with CL2006 showed that Ot B delayed paralysis by 21.4%, significantly increasing the PT50 from 8.0 to 10.1 days, which is comparable to 10.1 days in the curcumin-treated positive control group (p < 0.005; Fig. 2a, b, c, Supplementary Table 3).
Fig. 2.
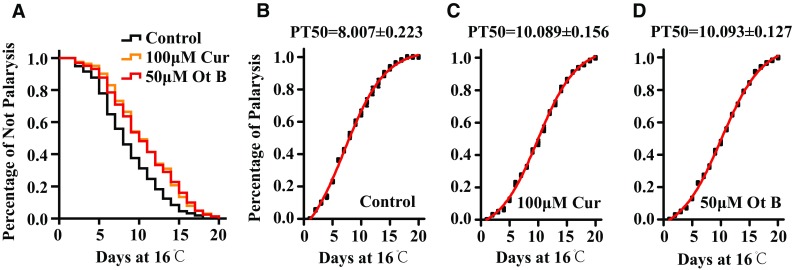
Ot B delayed progression of body paralysis in CL2006 worms. a Survival curves of control, Ot B treated and Cur treated animals. b Percentage of paralysis of control animals. c Percentage of paralysis of Cur treated animals. d Percentage of paralysis of Ot B treated animals. Each bar represents the mean of four independent experiments with error bars representing SEM. Each paralysis assay was conducted in triplicate and four trials were performed. Statistical details and repetition of this experiment are summarized in Table S3 (Supplementary information)
Our previous study showed that Ot B delayed aging and aging-related disorders [11], we were wondering if the delayed onset of paralysis was the result of an anti-aging effect, rather than a reduction in toxicity. To distinguish between these two hypotheses, we performed the paralysis assay in CL4176 worms, which express Aβ in muscle cells in a temperature-sensitive manner, leading to paralysis in larval animals (Fig. 3a) [18, 19]. At 30 h post temperature up-shift, Ot B decreased the paralysis from 73.3% to 51.1%, and 36 h later, 92.1% of the untreated worms became paralyzed, while only 79.4% of Ot B-treated and 78.7% of curcumin-treated positive control worms were paralyzed (p < 0.05; Fig. 3b, Supplementary Table 4). Together, these results indicated that Ot B delayed paralysis in young adults, and that this beneficial effect did not result from anti-aging.
Fig. 3.
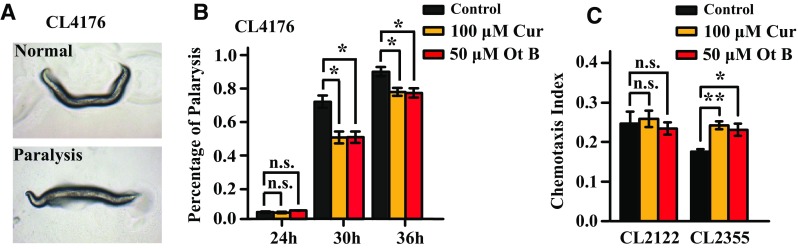
Ot B delayed progression of body paralysis in CL4176 and increased the chemotaxis response in CL2355 worms. a Photomicrograph showing CL4176 worms were paralyzed in larval animals. b Percentage of paralysis in CL4176 animals treated or non-treated with Ot B. 100 µM Curcumin was used as a positive control. Worms were grown at 16 °C for 48 h, and then transferred to 25°C. Paralysis experiments were carried out at 25 °C and calculated the paralysis of worms in 24, 30 and 36 h. c Chemotaxis index of CL2355 and its vector control, CL2122. Each bar represents the mean of three independent experiments with error bars representing SEM. ** represents p < 0.01,* represents p < 0.05, calculated using two-tailed t test. Statistical details and repetition of this experiment are summarized in Table S4, S5 (Supplementary information)
The CL2355 strain expresses Aβ in the neurons, which may more accurately represent toxicity of Aβ seen in AD [20]. The chemotaxis response in C. elegans is mediated by interneurons to stimulate the motor neurons [21]. To determine if Ot B could protect against Aβ toxicity to neurons, we performed the chemotaxis response assay in CL2355 worms, using the CL2122 strain as a vector control. The response is reported as a chemotaxis index. The results for the vector control (CL2122) showed no difference between Ot B treated, curcumin positive controls, and untreated worms. In CL2355 worms, Ot B significantly improved the chemotaxis response (p < 0.05; Fig. 3c, Supplementary Table 5). These results indicated that Ot B protected against Aβ toxicity to neurological functions.
Ot B Decreases Aβ Deposition in C. elegans Model of AD by Downregulating the Expression of Aβ
In CL2006 worms with muscle-specific Aβ expression, Aβ deposits are immunoreactive to anti-Aβ antibodies, and then these deposits bind to thioflavin S to produce deposits that can be viewed by confocal microscopy (Fig. 4a) [13]. To investigate if Ot B has a direct impact on the formation of Aβ, we conducted a thioflavin S staining experiment. The number of Aβ deposits was scored in the worm head region. The results showed that the mean number of Aβ deposits per nematode was significantly reduced in CL2006 worms treated with Ot B, compared with untreated worms at both day 3 and day 5 (p < 0.05; Fig. 4b, Supplementary Table 6). We next performed qRT-PCR to examine the effect of Ot B on the expression of Aβ. Ot B significantly reduced Aβ expression compared to untreated controls (p < 0.05; Fig. 4c, Supplementary Table 7). Collectively, these results showed the protection of Ot B against Aβ toxicity may result from the ability to decrease Aβ deposition by downregulating the expression of Aβ at the mRNA level.
Fig. 4.
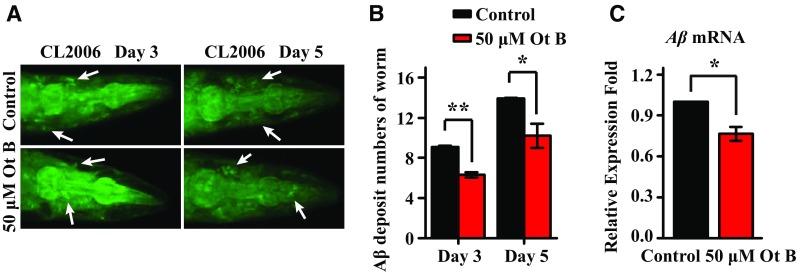
Ot B decreased Aβ deposition by depressing expression of Aβ. a Thioflavin S staining of CL2006. Ot B treated CL2006 worms were stained with thioflavin S on day 3 and day 5. White arrows indicate Aβ deposits. b Number of Aβ deposits in the worm head region. 50 μM Ot B significantly reduced the number of Aβ deposits in CL2006 both at day 3 and day 5. c The transcript level of Aβ, measured by qRT-PCR. The transcript level of Aβ was significantly downregulated by Ot B. The data was normalized to the expression of cdc-42. Each bar represents the mean value of three independent experiments with error bars representing SEM. ** represents p < 0.01,* represents p < 0.05, calculated using two-tailed t test. Statistical details and repetition of this experiment are summarized in Table S6, S7 (Supplementary information)
Ot B Alleviates Aβ Toxicity Mainly Through the HSF-1 Transcription Factor
Previous studies in C. elegans models of AD showed that the transcription factors DAF-16, SKN-1, and HSF-1 were involved in Aβ deposition [22, 23]. We performed qRT-PCR to test whether these regulators were involved in Ot B protection against Aβ toxicity.
We found no difference in the expression of daf-16 and its target genes, dod-3 and sip-1 between non-treated and treated worms, while the expression of another target gene, sod-3 was significantly upregulated (Fig. 5a, Supplementary Table 7). We speculated that DAF-16 may play a partial role in Ot B -mediated protection against Aβ toxicity. Meanwhile, there was no difference observed in the expression of skn-1 and its target genes, gst-4, gcs-1 and nit-1. This may indicate that SKN-1 is not essential in Ot B -mediated protection against Aβ toxicity (Fig. 5b, Supplementary Table 7).
Fig. 5.
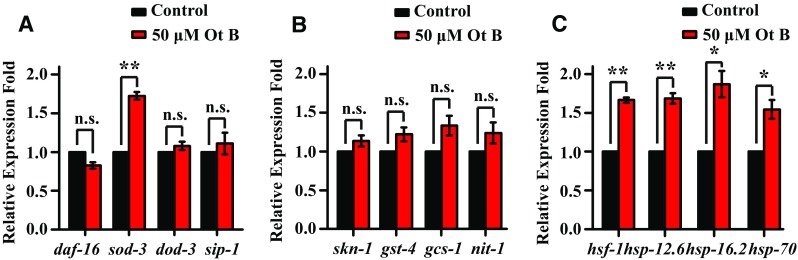
Gene expressions of transcription factors involved in Aβ deposition in controls and Ot B-treated C. elegans. a qRT-PCR measurement of mRNA transcripts for daf-16 and its targeted genes dod-3, sip-1, sod-3. b qRT-PCR measurement of mRNA transcripts for skn-1 and its targeted genes gst-4, gcs-1 and nit-1. c qRT-PCR measurement of mRNA transcripts for hsf-1 and its major target genes hsp-12.6, hsp-16.2 and hsp-70. qRT-PCR was carried out using method and normalized to the expression of gene cdc-42. In each experiment, control and experimental worms were conducted in parallel and repeated in three independent trials. Bars represent the mean value of three independent experiments with error bars representing SEM. ** represents p < 0.01,* represents p < 0.05, calculated using two-tailed t test. Statistical details and repetition of this experiment are summarized in Table S7 (Supplementary information)
HSF-1 was reported to alleviate Aβ toxicity by disaggregating and degrading large Aβ aggregates into peptides or amino acids [24]. Our results showed that the treatment of Ot B significantly upregulated the expression of hsf-1 and its targeted genes hsp-12.6, hsp-16.2 and hsp-70 (p < 0.05; Fig. 5c, Supplementary Table 7). Thus, heat shock protein (HSP) may be essential in Ot B-mediated protection against Aβ toxicity.
Conclusion
Otophylloside B (Ot B), a C-21 steroidal glycoside, is an essential active ingredient of Qingyangshen (C. otophyllum). We recently demonstrated anti-aging effects of Ot B in C. elegans, and it has been reported that Ot B is neuroprotective in epilepsy [9, 11]. Our present study showed that Ot B extended lifespan in a C. elegans model of AD, increased heat stress-resistance, delayed the process of paralysis, and increased the chemotaxis response. Collectively, these results indicated that Ot B protected against Aβ toxicity. The Aβ deposition assay and gene expression experiment showed this may result from the ability of Ot B to decrease Aβ deposition by down-regulating the expression of Aβ. The molecular mechanism study revealed that Ot B up-regulated the expression of several heat shock proteins (HSP), including hsf-1 and its target genes of hsp-12.6, hsp-16.2 and hsp-70. It may also increase the expression of sod-3 by partially activating the DAF-16, while SKN-1 was not essential in Ot B -mediated protection against Aβ toxicity. Taken together, these results indicate that Ot B has strong potential for development as a drug for AD prevention.
General Experimental Procedures
Chemicals and Strains
All strains were obtained from the Caenorhabditis Genetics Center (CGC) and maintained on NGM plates seeded with Escherichia coli OP50 at 16 °C. The following strains were used in this study: CL4176 dvIs27[myo-3::Aβ 3–42 let 3’UTR(pAF29); pRF4 (rol-6(su1006))], CL2006 dvIs2[pCL12(unc-54::Aβ 1–42 :) + pRF4], CL2122 dvIs15[(pPD30.38) unc-54(vector) + (pCL26) mtl-2::GFP], and CL2355 dvIs50[pCL45(snb-1::Aβ 1–42 ::3’ UTR(long) + mtl-2::GFP]I.
Ot B was dissolved in DMSO for storage and diluted in PBS to a concentration of 50 µM while in use. Then the dilutions were overlaid onto the NGM plates. The final DMSO concentration was 0.1% after adding the drugs to the plates, and the negative control group had the same concentration of DMSO.
Lifespan Assay
The lifespan assays were carried out in CL2006 at 16 °C. The strain was cultured for 2–3 generations before using for lifespan analysis. Lifespan assay were conducted as described previously [25]. In brief, for each assay, at least 40 synchronous L4 larvae or young adults were transferred to NGM plates containing inactivated OP50 (65 °C for 30 min), treated with 40 µM of FUdR to inhibit the growth of progeny and scored every other day. Animals were transferred to fresh plates with or without drugs every 2–4 days. All assays were carried out in triplicate, and at least three independent trials were performed. Statistics were calculated by using an SPSS package. The mean lifespan values were calculated by a log-rank (Kaplan–Meier) statistical test, with p < 0.05 accepted as statistically significant.
Heat Resistance Assay
For heat resistance assays, synchronous strains of CL2006 at L4 stage or young adults were transferred to plates with or without Ot B and incubated at 35 °C. Dead animals were counted every 2 h. For heat resistance recovery assays, synchronous animals of CL2006 at L4 stage or young adults were transferred to plates with or without Ot B at 35 °C for 7 h, then transferred to 16 °C and dead animals were counted daily [26]. For each assay, at least 30 synchronous nematodes were studied, and three independent trials were performed. For statistical analysis, p values were calculated by a two-tailed t test, each consisting of control and experimental animals at the same time.
Worm Paralysis Assay
For the paralysis assay of the CL4176 strain,late L3 larvae were grown at 16 °C for 48 h, then transferred to 25 °C to induce the expression of Aβ. Paralysis experiments were carried out at 25 °C. Calculation of the numbers of paralyzed worms was done at 24, 30 and 36 h after the transfer to 25 °C. For CL2006, paralysis experiments were carried out at 16 °C. Worms were checked every day until all worms were paralyzed. Worms were scored as paralyzed if they exhibited “halos” of cleared bacteria around their heads or moved their head only or did not move at all when they were gently touched by platinum worm pick [19]. Curcumin (100 µM) was used as a positive control. For each assay, at least 30 synchronous L4 larvae or young adult nematodes were studied. All paralysis plots were done in triplicate and three independent trials were performed. Statistics were calculated by using an SPSS package. The mean paralysis was calculated by a log-rank (Kaplan–Meier) statistical test, with p < 0.05 accepted as statistically significant.
Chemotaxis Assay
CL2355 and its vector control CL2122 were used in chemotaxis assays. Synchronized L1 larvae were treated with Ot B or the vehicle. Chemotaxis experiments were carried out at 23 °C as described previously [27]. Briefly, worms were placed to the center of the plate, and 1 μL 0.1% benzaldehyde in 100% ethanol a with 1 μL of 1 M sodium azide was placed on one side of the plate, and 1 μL 100% ethanol along with 1 μL of 1 M sodium azide was placed on the opposite side. Curcumin (100 µM) was used as a positive control. The chemotaxis index was defined as follows: (number of worms at the attractant location – number of worms at the control location)/total number of worms on the plate). A two-tailed t test was used to calculate p values.
Aβ Deposition Assay
For the Aβ deposition assay, CL2006 transgenic nematodes were fixed in 4% paraformaldehyde/PBS, pH 7.4, for 24 h at 4 °C, and then permeabilized in 5% fresh β-mercaptoethanol, 1% Triton X-100, 125 mM Tris pH 7.4, in a 37 °C incubator for 24 h. The worms were stained with 0.125% thioflavin S (Sigma) in 50% ethanol for 2 min, destained for 2 min in 50% ethanol, washed with PBS and mounted on slides for microscopy. Fluorescence images were acquired using a 40× objective of a fluorescence microscope. The Thioflavin S-reactive deposits anterior of the pharyngeal bulb in individual animals were scored [22]. A two-tailed test was used to calculate p values.
Gene Expression Analysis by q-Real-Time PCR
Synchronized CL2006 L1 larvae were transferred to NGM plates cultured with or without Ot B and incubated at 16 °C. Worms in the young adult stage were collected with M9 buffer, then total RNA was extracted using RNAiso Plus (Takara) and converted to cDNA with a High Capacity cDNA Reverse Transcription Kit (Applied Biosystems). The cDNA of candidate genes were amplified and quantified in a Power SYBR Green PCR Master Mix (Applied Biosystems) on an ABI 7500 DNA analyzer (Applied Biosystems). Relative fold-changes for transcripts were calculated using method, and normalized to cdc-42. The experiments were conducted in triplicate. The data were analyzed using a two-tailed t test, and a p value <0.05 was accepted as statistically significant.
Electronic supplementary material
Below is the link to the electronic supplementary material.
Acknowledgements
We thank the Caenorhabditis Genetics Center (CGC) for providing all the Caenorhabditis elegans strains used in this work. We thank Professor Wei-Min Yang from Kunming Medical University for providing curcumin as a gift used in our work. This work was partially supported by the Natural Science Foundation of China (81671405 and 81370453), Natural Science Foundation of Yunnan province (2013FA045 and 2015FB172), and Open Funds of Guangdong Key Laboratory of Marine Materia Medica (LMM2016-1).
Compliance with Ethical Standards
Conflict of interest
The authors declare no conflict of interest.
References
- 1.Udeochu JC, Shea JM, Villeda SA. Clin. Exp. Neuroimmunol. 2016;7:114–125. doi: 10.1111/cen3.12307. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 2.Cummings J, Aisen PS, DuBois B, Frolich L, Jack CR, Jr, Jones RW, Morris JC, Raskin J, Dowsett SA, Scheltens P. Alzheimers Res. Ther. 2016;8:39. doi: 10.1186/s13195-016-0207-9. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 3.Ferreira ST, Klein WL. Neurobiol. Learn. Mem. 2011;96(4):529–543. doi: 10.1016/j.nlm.2011.08.003. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 4.Viola KL, Klein WL. Acta Neuropathol. 2015;129(2):183–206. doi: 10.1007/s00401-015-1386-3. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 5.Wang Q, Yang F, Guo W, Zhang J, Xiao L, Li H, Jia W, Huang Z. Rejuvenation Res. 2014;17(2):205–208. doi: 10.1089/rej.2013.1512. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 6.Dostal V, Link CD. J. Vis. Exp. 2010;44:e2252–e2252. doi: 10.3791/2252. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 7.Lublin AL, Link CD. Drug Discov. Today Technol. 2013;10(1):e115–e119. doi: 10.1016/j.ddtec.2012.02.002. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 8.Zhou JK, Lv J, Zhou P. J. Guizhou Norm. Coll. 2010;26(12):23–26. [Google Scholar]
- 9.Long X, Dai FC, Xiao XH, He J, He ZD. J. Jilin Med. 2014;19:4135–4137. [Google Scholar]
- 10.Li JL, Zhou J, Chen ZH, Guo SY, Li CQ, Zhao WM. J. Nat. Prod. 2015;78(7):1548–1555. doi: 10.1021/np501058b. [DOI] [PubMed] [Google Scholar]
- 11.Yang J, Wan QL, Mu QZ, Wu CF, Ding AJ, Yang ZL, Qiu MH, Luo HR. Nat. Prod. Bioprospect. 2015;5(4):177–183. doi: 10.1007/s13659-015-0064-4. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 12.Gutierrez-Zepeda A, Santell R, Wu Z, Brown M, Wu Y, Khan I, Link CD, Zhao B, Luo Y. BMC Neurosci. 2005;6:54. doi: 10.1186/1471-2202-6-54. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 13.Morita M, Osoda K, Yamazaki M, Shirai F, Matsuoka N, Arakawa H, Nishimura S. Brain Res. 2009;1295:186–191. doi: 10.1016/j.brainres.2009.08.002. [DOI] [PubMed] [Google Scholar]
- 14.Arya U, Dwivedi H, Subramaniam JR. Exp. Gerontol. 2009;44(6–7):462–466. doi: 10.1016/j.exger.2009.02.010. [DOI] [PubMed] [Google Scholar]
- 15.Dostal V, Roberts CM, Link CD. Genetics. 2010;186(3):857–866. doi: 10.1534/genetics.110.120436. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 16.Guo H, Cao M, Zou S, Ye B, Dong Y. J. Gerontol. A Biol. Sci. Med. Sci. 2015 doi: 10.1093/gerona/glv165. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 17.Regitz C, Dussling LM, Wenzel U. Mol. Nutr. Food Res. 2014;58(10):1931–1940. doi: 10.1002/mnfr.201400014. [DOI] [PubMed] [Google Scholar]
- 18.Xin L, Yamujala R, Wang Y, Wang H, Wu WH, Lawton MA, Long C, Di R. PLoS ONE. 2013;8(5):e63874. doi: 10.1371/journal.pone.0063874. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 19.Takahashi A, Watanabe T, Fujita T, Hasegawa T, Saito M, Suganuma M. Biosci. Biotechnol. Biochem. 2014;78(7):1206–1211. doi: 10.1080/09168451.2014.921553. [DOI] [PubMed] [Google Scholar]
- 20.Wu Y, Wu Z, Butko P, Christen Y, Lambert MP, Klein WL, Link CD, Luo Y. J. Neurosci. 2006;26(50):13102–13113. doi: 10.1523/JNEUROSCI.3448-06.2006. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 21.Hobert O. J. Neurobiol. 2003;54(1):203–223. doi: 10.1002/neu.10168. [DOI] [PubMed] [Google Scholar]
- 22.Hsu AL, Murphy CT, Kenyon C. Science. 2003;300(5622):1142–1145. doi: 10.1126/science.1083701. [DOI] [PubMed] [Google Scholar]
- 23.Li X, Matilainen O, Jin C, Glover-Cutter KM, Holmberg CI, Blackwell TK. PLoS Genet. 2011;7(6):e1002119. doi: 10.1371/journal.pgen.1002119. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 24.Cohen E, Bieschke J, Perciavalle RM, Kelly JW, Dillin A. Science. 2006;313(5793):1604–1610. doi: 10.1126/science.1124646. [DOI] [PubMed] [Google Scholar]
- 25.Zheng SQ, Huang XB, Xing TK, Ding AJ, Wu GS, Luo HR. J. Gerontol. A Biol. Sci. Med. Sci. 2016 doi: 10.1093/gerona/glw105. [DOI] [PubMed] [Google Scholar]
- 26.Ogawa T, Kodera Y, Hirata D, Blackwell TK, Mizunuma M. Sci. Rep. 2016;6:21611. doi: 10.1038/srep21611. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 27.Zhang XG, Wang X, Zhou TT, Wu XF, Peng Y, Zhang WQ, Li S, Zhao J. Front. Pharmacol. 2016;7:227. doi: 10.3389/fphar.2016.00227. [DOI] [PMC free article] [PubMed] [Google Scholar]
Associated Data
This section collects any data citations, data availability statements, or supplementary materials included in this article.