

Abstract
Background:
Glucose homeostasis derangement is a common pathophysiology of malaria whose aetiology is still controversial. The Plasmodium parasite, immunological and inflammatory responses, as well as chemotherapeutics currently used cause hypoglycaemia in malaria. Anti-parasitic and anti-disease drugs are required to combat malaria while ameliorating the pathophysiology of the infection. Asiatic acid has anti-hyperglycaemic, antioxidant, pro-oxidant properties useful in glucose homeostasis but its influence in malaria is yet to be reported. Here we present findings on the influence of asiatic acid on glucose metabolism in vivo using P. berghei-infected Sprague Dawley rats.
Materials and Methods:
Acute as well as sub-chronic studies were carried out in vivo where physicochemical properties and glucose homeostasis were monitored after administration of asiatic acid (10mg/kg) in both non-infected and infected animals. Glucose metabolism associated biochemical changes in malaria were also investigated.
Results:
In acute studies, asiatic acid improved oral glucose response while in the sub-chronic state it maintained food and water intake and suppressed parasitaemia. Normoglycaemic control was maintained in infected animals through insulin suppression and increasing glucagon secretion, in both acute and chronic studies. Asiatic acid administration curtailed lactate concentration towards normal.
Conclusion:
Per oral post-infection asiatic acid administration preserved drinking and eating habits, inhibited sickness behaviour while suppressing parasitaemia. Reciprocal relationship between insulin and glucagon concentrations was maintained influencing glucose homeostasis positively and inhibition of hyperlactaemia in malaria.
Abbreviations: ip -intraperitoneal, po -per oral, ig -intragastric, AA-Asciatic acid, OGTT-oral glucose tolerance test, OS-oxidative stress, ROS-reactive oxygen species, NO-nitric oxide, ONOO- - peroxynitrite, BRU-Biomedical Research Unit, SD-Sprague Dawley,
Keywords: Asiatic acid, malaria, Plasmodium berghei, glucose homeostasis, anti-disease, anti-parasitic
Introduction
Malaria is an immunological disease displaying systematic inflammatory aspects with marked cachexia (Goldring, 2004, Schofield, 2007). Glucose homeostasis in malaria is critical in the more vulnerable groups of pregnant women and children <5 years of age with highest disease associated morbidity and mortality rates (White N.J. et al., 1983, English et al., 1998). Either group has less immune competency, heighten or constant demand for energy supply, and tend to experience slight physiological deficits in a more exaggerated way (Miller et al., 2013). Hypoglycaemia, cognitive impairment, severe malaria anaemia (SMA) non-respiratory acidosis and renal insufficiency are recognised facets of a post malarial treatment syndrome (Mackintosh et al., 2004). Aetiology of malaria hypoglycaemia is multi-faceted. Anti-parasitic treatment therefore may be inadequate to avert or ameliorate malaria-induced glucose homeostasis derangements. Glycolysis has also been observed to have more homeostatic effects beyond being involved in nucleic acid, lipid and amino acid biosynthesis in what is called the “Warburg effect” shown by increased glycolysis in the presence of oxygen (Warburg, 1930, Warburg, 1956; Pedersen, 2007). Glycolysis is involved in signal transduction regulating immunometabolic processes (Ho, 2016), histone acetyl-CoA acetylation control of early differentiation of embryonic stem cells (Moussaieff, 2015) and immune system function modulation through interferon-γ and IL-2 biosynthesis (Zhao, 2016). This makes glucose metabolism controlling agents in malaria have possible influence on the disease outcomes.
Asiatic acid (AA), is a phytotherapeutic with antioxidant, antihyperglycaemic, antihyperlipidaemic (Ramachandran & Saravanan, 2014) as well as emerging antimalarial properties (Mavondo, 2016). However, there is a lack of information on AA’s influence on glucose homeostasis in malaria. Desirous to unravel further the metabolic effects of AA, we have investigated the influence of this phytomedicinal in murine malaria. Here we report on the influence of AA on glucose homeostatic changes in Plasmodium berghei malaria infection in young (90-120g) male Sprague Dawley rats.
Materials and Methods
Drugs, Chemical and Accessories
Asiatic acid (AA 97% purity), Giemsa stain, dimethyl sulphoxide (DMSO), chloroquine diphosphate were purchased from Sigma-Aldrich (St. Louis, Missouri, USA). All other chemicals and reagents were of analytical grade.
Animals
Male Sprague-Dawley (SD) rats weighing 90-120 g were obtained from the Biomedical Research Unit (BRU) of the University of KwaZulu-Natal where they were bred and housed for the entire experiment period. The animals were kept under maintained laboratory conditions of constant temperature (22±1 °C); C02 (<5000 ppm), humidity of 55±5% and illumination (12 h light/dark cycle). Food, standard rat chow (Meadows Feeds, Pietermaritzburg, South Africa) and water were supplied to the animals ad libitum. Animals were sacrificed at specific time point up to 12 days for the non-treated infected control (IC) and up to 21 days for the rest of the experimental animals. Lethal anaesthetic inhalation of isofor (Safeline Pharmaceuticals, Rooderport, South Africa) in a gas chamber (100mg/kg) was used to sacrifice the animals. All experiments and protocols used in this study were reviewed and approved by the animal ethics committee of the University of KwaZulu Natal (UKZN) with ethical clearance numbers 079/14/Animal and 013/15/Animal issued.
Malaria Parasite
Chloroquine-susceptible strain of P. berghei ANKA, was a kind donation from Professor Peter Smith (University of Cape Town, Division of Clinical Pharmacology, South Africa). P. berghei (105 parasitized red blood cells [pRBC’s] suspension in saline) was inoculated intraperitoneal (ip) into stock animals which were sacrificed after 12 days and the infected blood was harvested, washed and stored at -80°C in freezing media (30% glycerol in phosphate buffer) until used.
Experimental Design
The study was divided into two viz; an acute and a sub-chronic study (lasting 21 days). Experimental animals were divided into 5 groups (n = 6 per group) as follows:
Non infected absolute control [3mL/kg H2O] (AC)
Non-infected treated control [AC+AA 10mg/kg] (NIC)
Infected non-treated control (IC)
Infected treated with CHQ 30mg/kg (30CHQ)
Infected treated with AA 10mg/kg (10mg) (AA10)
Preliminary studies using 3 concentrations of AA (5, 10, 20mg/kg) indicated that the AA10 oral administration had the most antimalarial efficacy hence it is the dose of choice in this study.
Malaria Induction
P. berghei (105 parasitized red blood cells [pRBC’s] suspension in saline) was inoculated ip and control animals received equivalent amount of saline.
Monitoring of Parasitaemia
Peripheral blood was collected using the tail prick method (Rangaraj et al., 2014). Parasitaemia % monitored after 72 hours (pre-patent period), every third day up to day 7 [patent period] (Changa & Stevenson, 2004), every day during treatment period of five days and thereafter every other day (post-treatment period) until day 21. A 15-20% parasitaemia on day 7, confirmed by May-Grünwald-Giemsa staining under a microscope (Olympus Cooperation, Tokyo, Japan), was considered as stable state malaria.
Asiatic Acid Influence on Biophysical Changes
Body weights, food and water intake were monitored gravimetrically in all animals at 09h00 every third day during the pre-treatment, treatment and post treatment periods. The effects of AA on these parameters were examined.
Asiatic Acid Administration in Acute Studies (Oral Glucose Tolerance Response)
With parasitaemia at 15-20% in infected animals, all animals were fasted for 16 hours and a 240-minute oral glucose (0.86g/kg) tolerance response (OGTR) was carried out to distinguish effects of AA and malaria on glucose homeostasis. The time-glucose concentration course was determined to map out the influence of AA on the rate of glucose disappearance from plasma. Animals were sacrificed after the experiments by exposing to isofor (100 mg/kg) inhalation anaesthesia [Safeline Pharmaceuticals, Rooderport, South Africa]. Blood samples were collected by cardiac puncture into pre-cooled lithium heparin tubes onto melting ice, immediately centrifuged for 15 minutes at 959 x G in a 4°C centrifuge (Eppendorf International, Hamburg, Germany) and plasma separated. Plasma was stored in a Bio Ultra freezer (Snijers Scientific, Tilburg, Netherlands) at -80°C until AA influence on hormonal changes was assessed.
Asiatic Acid Influence on Insulin and Glucagon Concentrations Post Glucose Bolus Administration
Insulin concentration was determined quantitatively in plasma using the Mercodia Ultrasensitive Rat Insulin ELISA (Mercodia AB, Uppsala, Sweden) following the manufacturer’s instructions. The lower and upper limits of detection were <3.38pmol/L and 783pmol/L, respectively. Plasma glucagon concentration was determined using Elabscience Rat GC (Glucagon) ELISA kit Catalog No: E-EL-R0425 (Elabscience Biotechnology Co. Ltd, WuHan, P.R.C.) as per the manufacturer’s instructions. Same methods were used in the sub-chronic studies.
Sub-chronic Studies
Post-infection po AA administration’s influence on glucose metabolism and murine-malaria in SD rats was carried out in animals housed individually in Makrolon polycarbonate metabolic cages (Techniplast, Labotec, South Africa), with food and water availed ad libitum, at the Biomedical Resource Unit, University of KwaZulu Natal over a 3-week period. Chloroquine (CHQ) control (30mg/kg) was dissolved in distilled water and administered as for AA. AA was dissolved in DMSO (0.5mL) and diluted with distilled water to give a stock solution of 10mg/kg. AA (10mg/kg) was administered once daily po (at 09h00) for five days. CHQ (30mg/kg) was administered twice daily (09h00 hours and 17h00 hours) according to posology developed in our laboratory. A ball-tipped, 18-gauge gavage needle (Kyron Laboratories (Pty) LTD, Benrose, South Africa) attached to a 1 ml syringe was used. Animals were sacrificed at day 8, 12 and 21. The IC animals were sacrificed at day 12 for ethical reasons.
Plasma and Tissue Sample Harvesting
Influence of AA on glucose homeostasis, hormonal, immunology and inflammatory changes as well as other biochemical parameters were determined in either plasma or organ tissues. Animal sacrifice and plasma collection was as described in the acute studies (5.2.4). The liver, kidney, muscle and heart were removed, snap frozen in liquid nitrogen and stored together with the plasma in a Bio Ultra freezer (Snijers Scientific, Tilburg, Netherlands) at -80°C until use.
Influence of Asiatic Acid on Glucose Utilization in Sub-Chronic Conditions
The effects of AA on blood glucose homeostasis in malaria were evaluated by measuring blood glucose every other day during pre-patent, daily during patent/treatment and every three days post treatment periods in all the animal groups using blood glucose testing strips (Lifescan, Zug, Switzerland).
Influence of Asiatic Acid on Tissue Glycogen Storage
Influence of AA10 on glycogen storage capacity in liver, muscle and kidney were compared amongst AA treated infected animals and controls according to the method by Seifter (Seifter et al., 1949), with some modifications.
Influence of Asiatic Acid on Plasma Lactate Concentration
The influence of futile glucose homeostasis was indicated by levels of lactate in plasma which were estimated using CobasR Accutrend Plus using Accutrend BM-Lactate strips (Roche Diagnostics GmbH, Mannheim, Germany).
Statistical Analysis
Data are presented as the means ± standard error of mean (SEM). Overall statistical comparisons between the control means and experimental groups were performed with GraphPad Prism Software version 5.00, (GraphPad Prism Software, San Diego, California, USA), using one-way analysis of variance (ANOVA), followed by Tukey-Kramer post hoc multiple comparison test. A value of p < 0.05 was considered significant.
Results
Parasitaemia Monitoring
Compared to IC and CHQ, AA10 administration had significantly lower %parasitaemia (*, ≠p<0.05. Table 1) at relevant time points. Compared to both IC and CHQ, AA (10mg/kg) had a lower peak %parasitaemia (*, ≠ p<0.05. Table 1) AA10 suppressed parasitaemia to undetectable levels compared to CHQ (ψ p<0.05. Table 1) Compared to IC, CHQ had lower %parasitaemia during patent/ treatment period (ψ p<0.05. Table 1)
Table 1.
Comparison of %parasitaemia changes over time (21 days). NIC-non infected treated control, IC- infected control; CHQ-chloroquine treated infected control. Values are presented as means ± SEM, (n=6 per group). *, ≠ p<0.05 compared to IC and CHQ, respectively. ψ p<0.05 CHQ compared to IC
| Protocol | Experimental Groups | %Parasitaemia per time point | ||||||
|---|---|---|---|---|---|---|---|---|
| Pre-patent | Patent/ treatment period | Post-treatment | ||||||
| Day 7 | Day 9 | Day 12 | Day 15 | Day 21 | ||||
| Post-infectionAAadministration | Ic | 3.7±0.21 | 13.68 ±0.98 | 33.43 ±1.07 | 56.52 ±3.20 | N/A | N/A | |
| 30CHQ | 2.7±0.41 | 18.68 ±1.89ψ | 42.08 ±1.325ψ | 37.37 ±4.36ψ | 23.38 ±1.32 | 1.14 ±1.21 | ||
| AA10 | 3.2±0.90 | 13.47 ±0.95*≠ | 21.13 ±2.031*≠ | 26.67 ±2.91**≠ | 3.71 ±0.12≠ | 0.00 ±0.00≠ | ||
Food and Water Intake
A10 administration significantly preserved food and water intake as compared to the IC, and CHQ (*, ≠ p<0.05, respectively. Figure 1) Compared to the AC and NIC, CHQ treated animals had significantly lower food and water intake (ψ, γ p<0.05, respectively). IC had significantly lower intake compared to AC and NIC (δ, χ p<0.05, respectively. Figure 1) Compared to the NIC, AC had higher food and water intake (β p<0.05. Figure 1)
Figure 1.

Comparison of food and water intake as influenced by AA10. NIC-non infected treated control, IC- infected control; CHQ-chloroquine treated infected control. Values are represented as means ± SEM. α, *, ≠ p<0.05 compared to the NIC, IC and CHQ, respectively. ψ p<0.05 CHQ compared to IC. γ p<0.05 CHQ compared to the NIC. χ p<0.05 IC compared to NIC. δ p<0.05 IC compared to AC. β p<0.05 AC compared to NIC.
Influence of AA on Glucose under Acute Conditions (OGTR)
OGTR as influenced by AA10 administration was compared to the NIC, IC and 30 CHQ groups in Figure 2. AA10 administration significantly improved OGTR compared to IC and CHQ (*, ≠ p<0.05, respectively. Figure 2) At the 15 minute time point, AA10 ablated glucose spike compared to both IC and CHQ (*, ≠ p<0.05, respectively. Figure 2) Compared to NIC, the IC and CHQ groups had significantly poor OGTR (γ, ψ p<0.05, respectively).
Figure 2.
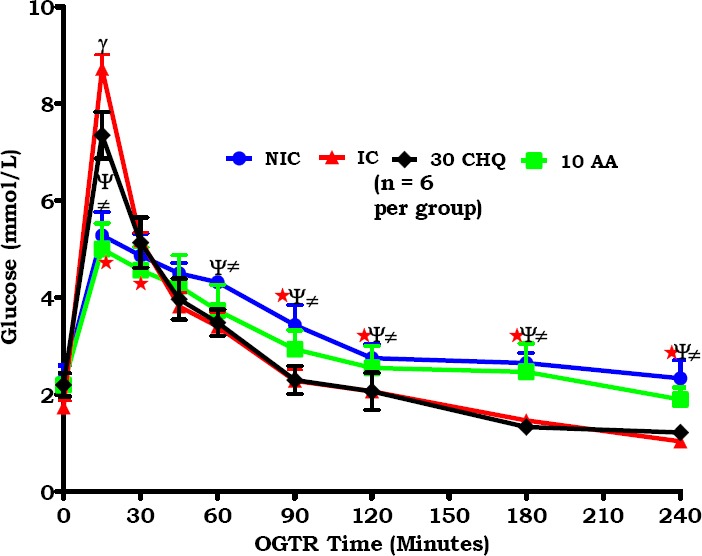
Oral glucose test response curve for AA10 compared to controls. NIC-non infected treated control, IC- infected control; CHQ-chloroquine treated infected control. Values are presented as means ± SEM, (n=6 per group). (*, ≠ p<0.05 compared to IC and CHQ, respectively. ψ p<0.05 CHQ compared to IC. γ p<0.05 IC compared to NIC.
Asiatic Acid Influence on Insulin and Glucagon Concentrations Post Bolus Administration
Insulin and glucagon concentration changes in OGTR as influenced by AA10 administration was compared to controls in Table 2. AA10 significantly increased glucagon and lowered insulin compared to IC and CHQ (*, ≠ p<0.05, respectively). NIC had significantly lower insulin and higher glucagon compared to IC and CHQ (γ, χ p<0.05, respectively).
Table 2.
Influence of AA (10mg/kg) on acute insulin and glucagon secretion compared to controls. NIC-non infected treated control, IC- infected control; CHQ-chloroquine treated infected control. Values are presented as means ± SEM, (n=6 per group). (*, ≠ p<0.05 compared to IC and CHQ, respectively. γ, χ p<0.05 NIC compared to CHQ and IC.
| Hormone | Parameter | Concertation after 20 hour fast |
|---|---|---|
| Insulin (pmol/L) | NIC | 8.12±10.76 |
| IC | 225.38±5.72 χ | |
| 30 CHQ | 258.53±11.78γ | |
| AA10 | 8.93±3.87*≠ | |
| Glucagon (pmol/L) | NIC | 628±32.66 |
| IC | 98±6.98χ | |
| 30CHQ | 76±3.87γ | |
| AA10 | 498.76±7.12*≠ |
Influence of AA on Glucose Utilization in Sub-Chronic Conditions
Administration of AA10 significantly preserved blood glucose compared to IC and CHQ (*, p<0.05, respectively. Figure 3) at days 7-12 and CHQ (≠ p<0.05 Figure 3) at days 7-18). IC displayed lower glucose compared to NIC (χ p<0.05 Figure 3). CHQ administration lowered glucose levels compared to NIC (γ p<0.05 Figure 3) but increased it compared to IC (ψ p<0.05 Figure 3) at days 7-12. CHQ treatment glucose lowering continued post treatment compared to NIC at day 18
Figure 3.

A comparison of the influence of AA on blood glucose concentration in sub-chronic studies over time. NIC-non infected treated control, IC- infected control; CHQ-chloroquine treated infected control. Values are represented as means ± SEM. α,*, ≠ p<0.05 compared to the NIC, IC and CHQ, respectively. ψ p<0.05 CHQ compared to IC. γ p<0.05 CHQ compared to the NIC. χ p<0.05 IC compared to NIC.
Influence of AA Administration on Liver Glycogen Storage
Glycogen concentration, as influenced by AA10, was compared amongst animal groups in Figure 6. Administration of AA significantly increased glycogen in liver as compared to the IC, and CHQ (*, ≠ p<0.05, respectively). Compared to the AC and NIC, CHQ treated animals had lower glycogen levels (ψ, γ p<o.o5, respectively). IC had significantly lower glycogen compared to AC and NIC (δ, χ p<0.05, respectively).
Figure 4.

A comparison of liver glycogen concentration as influenced by AA 10mg/kg administration. NIC-non infected treated control, IC- infected control; CHQ-chloroquine treated infected control. Values are represented as means ± SEM. α,*, ≠ p<0.05 compared to the NIC, IC and CHQ, respectively. ψ p<0.05 CHQ compared to IC. γ p<0.05 CHQ compared to the NIC. χ p<0.05 IC compared to NIC. δ p<0.05 IC compared to AC.
Figure 5.

AA (10mg/kg) influence on muscle glycogen compared to other animal groups. NIC-non infected treated control, IC-infected control; CHQ-chloroquine treated infected control. Values are represented as means ± SEM. α,*, ≠ p<0.05 compared to the NIC, IC and CHQ, respectively. ψ p<0.05 CHQ compared to IC. γ p<0.05 CHQ compared to the NIC. χ p<0.05 IC compared to NIC. δ p<0.05 IC compared to AC.
Figure 6.
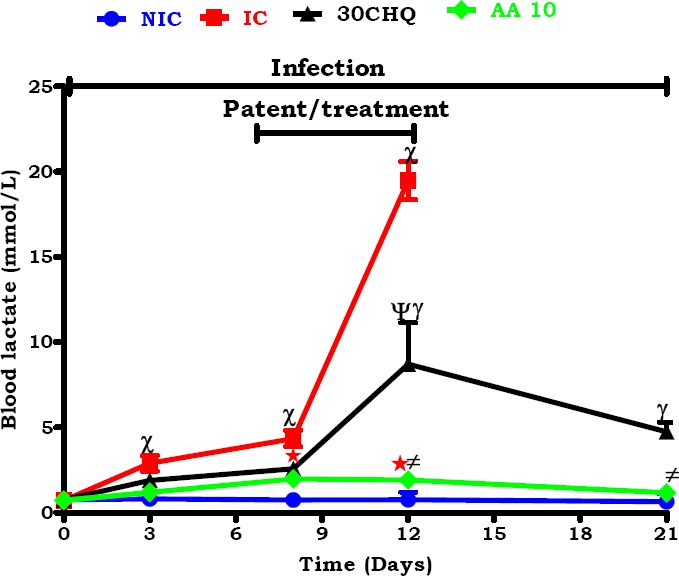
Influence of AA10 administration on lactate concentration changes compared to controls. NIC-non infected treated control, IC- infected control; CHQ-chloroquine treated infected control. Concentrations are presented as means ±SEM (n = per group). *, ≠ p<0.05 compared to the IC and CHQ, respectively. ψ p<0.05 CHQ compared to IC. γ p<0.05 CHQ compared to the NIC. χ p<0.05 IC compared to NIC.
Influence of AA Administration on Muscle Glycogen Storage
Glycogen concentration, as influenced by AA10, was compared to controls in figure 5. Administration of AA10 significantly increased glycogen when compared to the IC, and CHQ (*, ≠ p<0.05, respectively). Compared to the AC and NIC, CHQ treated animals had lower glycogen levels (ψ, γ p<0.05, respectively). IC had significantly lower glycogen compared to AC and NIC (δ, χ p<0.05, respectively).
Plasma Lactate Production
A comparison of the influence AA10 had on lactate production to controls was made in Figure 6. AA10 administration significantly lowered blood lactate concentration compared to the IC and 30CHQ (*, ≠ p<0.05, respectively). IC had higher blood lactate concentrations than NIC and CHQ (χ, ψ p<0.05, respectively). CHQ displayed higher blood lactate concentrations than NIC (γ p<0.05).
Plasma Insulin Levels
Relationships of AA10 administration to insulin secretion were compared to various animals groups in Figure 7, displaying significantly lowered values compared to AC, NIC, IC and CHQ (μ, α,*, ≠ p<0.05, respectively). AC had significantly higher basal insulin levels compared to NIC (β p<0.05), although below reference interval (RR). NIC insulin was significantly lower than IC (χ p<0.05). Compared to the NIC and IC, CHQ treatment had significantly higher (γ p<0.05) and lower (ψ p<0.05) values, respectively.
Figure 7.
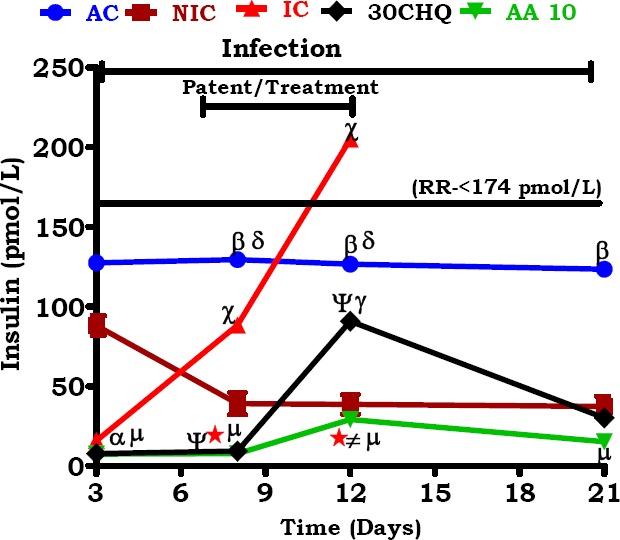
AA10 influence on insulin levels in comparison to the AC (absolute control), NIC, IC and 30 CHQ. NIC-non infected treated control, IC- infected control; CHQ-chloroquine treated infected control. Concentrations are presented as means ±SEM (n = per group). μ, α,*, ≠ p<0.05 compared to the AC, NIC, IC and CHQ, respectively. ψ p<0.05 CHQ compared to IC. γ p<0.05 CHQ compared to the NIC. χ p<0.05 NIC compared to IC. β p<0.05 NIC compared to AC. δ p<0.05 IC compared to AC.
Glucagon Concentrations
Relationships of changes of glucagon concentrations over time as influenced by AA10 were compared to controls in Figure 8. AA10 administration had significantly higher concentrations compared to AC, NIC, IC and CHQ treatment (μ, α, *, ≠ p<0.05, respectively). Glucagon concentrations were higher in animals treated with CHQ compared to NIC and IC groups (γ, ψ p<0.05, respectively). IC group glucagon concentrations were significantly higher than NIC (χ p<0.05).
Figure 8.

Influence of AA10 on glucagon (GLN) concentration changes over time in malaria. AC-absolute control, NIC-non infected treated control, IC- infected control; CHQ-chloroquine treated infected control. Concentrations are presented as means ±SEM (n = per group). μ, α, *, ≠ p<0.05 compared to the AC, NIC, IC and CHQ, respectively. ψ p<0.05 CHQ compared to IC. γ p<0.05 CHQ compared to the NIC. χ p<0.05 NIC compared to IC.
Discussion
Glucose metabolic disturbance is an intricate balance between supply, transportation, utilization or storage which the malaria parasite exploit successfully to systematically weaken the body defence systems while optimising its own survival (Olszewski et al., 2009), with poor prognosis in children under five years (English et al., 1998, Agbenya et al., 2000) and in pregnant women (Croft, 2000). Hypoglycaemia has a prevalence of approximately 10% in both adults and paediatric patients (White N.J. et al., 1983) and up to 25°% in children (Osier et al., 2003). There are a number of systems that deplete plasma glucose in malaria which can be targets for AA intervention. We report here the influence of AA10 on glucose utilization in an acute study and over a period of 21 days in P. berghei-infected Sprague Dawley rats. All animal groups which were inoculated with P. berghei murine malaria parasite developed severe malaria (SM) by Day 7 (patent malaria period) at which period treatment was effectuated. Parasitaemia percentages were extreme in non-treated animals with a perceptible difference in suppression of the parasite in the AA10 orally administered groups which may be attributable to the influence of AA. Moreover, AA10 surpassed the efficacy of 30CHQ oral treatment (Table 1) although a single dose of AA10 was administered compared to the double dose of CHQ, daily.
Concomitant with % parasitaemia changes was a significant disparity in food and water intake between AA10 administered animals and IC groups. Animals that were administered with oral AA10 displayed a positive preservation of food and water intake at all relevant time periods during the study which correlated well with AC and contrasted with the IC and 30CHQ treated groups (Figure 1). There seems to be a causal relationship between parasite infection and food and water intake together with %weight gain (results not shown) seeing that, compared to the AA10, the IC group posted more averse outcomes in the experiments.
Malaria, like other chronic diseases, is associated with reduced food and water intake in what is termed the sickness behaviour (Hart, 1988). The natural response to infection or injury is for the animal to down-regulate normal activities (including eating and drinking), reorganize its resources to overcome the infection and re-establish homeostatic functions (Clark et al., 2008). However, when the insult perpetuates, the associated somnolence becomes a liability to the animal with energy depletion and anorexia leading to cachexia (Clark & Vissel, 2014). The resulting malnutrition and inanition accelerated a wasting diathesis as we have observed in the IC group despite ad libitum provision of food and water. The loss of appetite in sickness behaviour is driven by tumour necrosis factor-α (TNF-α). TNF-α secretion is an intrinsic mechanism to create a hostile environment for the pathogen (Grimble, 1992) which becomes aberrant in malaria due to unregulated immune response. However, AA10 administration preserved the social habit of eating and drinking showing possible inhibition of TNF-α and inflammation or the nuclear factor-kp (NF-Κβ) suppression which directs inflammatory responses (Huang et al., 2011).
In the acute study, AA10 improved oral glucose tolerance response [OGTR] (Figure 2). Administration of AA ablated the rise in glucose at the 15 minute time point while the there was a sharp rise in glucose in both the IC and CHQ treated groups. This could be the result of the inflammatory cytokines (TNF^-induced insulin resistance that occurs in diabetes mellitus (Fen^ndez-Real & Ricart, 1999) as well as in malaria (Acquah et al., 2014). The anti-inflammatory effect of AA has been reported as well as its capacity to potentiate insulin (Ramachandran & Saravanan, 2013, Ramachandran & Saravanan, 2014) resulting in a faster uptake of glucose without inducing hypoglycaemia. Insulin levels were observed to be lower in AA10 administered animals as well as in the NIC as compared to either the IC or the 30CHQ treated groups showing the suppressive effect of low glucose levels (Table 2). Terminal glucagon concentration, on the other hand, was elevated in the AA10 group as well as the NIC showing how the animals did not experience severe hypoglycaemia even after a 20 hour fast. The same hormone levels were, however, suppressed in the IC and the 30CHQ treated animals during the acute studies. This scenario seemed to occur in acute circumstance of low glucose concentration during fasting than in the sub-chronic studies.
In the sub-chronic study there was an inflection in the glucose-time course which corresponded with the rise in parasitaemia in the AA10 administered animals. But there was a steep decline in glucose concentration showing a terminal hypoglycaemic trend that is typical of the malaria parasite effect on glucose homeostasis. 30CHQ treatment showed a contraction of glucose-time curve during the patent/treatment phase of the study. Chloroquine (Cansu & Korkmaz, 2008) and quinine (Elbadawi et al., 2011) have been shown to have hypoglycaemic effects due to their insulin memetic effect (English et al., 1998) which persisted well after treatment had ceased and parasitaemia resolved (English et al., 1998). We have also shown in our laboratory that CHQ had a glucose lowering effect in Sprague Dawley rats in malaria with a concomitant rise in insulin (Musabayane et al., 2010) which was ameliorated by transdermal delivery of CHQ (Murambiwa et al., 2013). The hyperinsulinemia of malaria was clearly enunciated by our finding in the IC group which displayed a steep rise in insulin (Figure 7) correlating well with the changes in %parasitaemia (Table 1). Indeed, this collateral rise in insulin predetermined suppression of glucagon (Figure 8) which invariably decreased plasma glucose in the group.
Malaria causes hyperinsulinemia as a result of increasing insulin resistance created by inflammation which disturbs glucose homeostasis. However, we observed correction of this trend when AA10 was administered. To maintain glucose concentration in malaria, glucagon was elevated in the AA10 administered animals in comparison with both the AC and NIC. The presence of AA and absence of malaria potentiated insulin activity such that a low concentration of glucose was maintained in the NIC compared to the AC during the fed state of the experiments. Indeed, AA has been reported to increase glycolysis enzymes in streptozotocin-induced diabetes while inhibiting gluconeogenesis and glycogenolysis (Ramachandran & Saravanan, 2013) facets that will have preserved food and water intake until the animal recovered from malaria. Rationally, in an increased in vivo glucose oxidation that is accompanied by either inaccessible stored energy sources (AA effect) or depleted stores (malaria effect), the animal will seek to replenish the energy deficit by foraging. The food and water intake which was preserved in our AA10 administered animals but not observed in the IC may be attributed to AA influencing these biophysical activities fostered by parasitaemia eradication. The antimalarial, anti-inflammatory, antioxidant and anti-hyperglycaemia influence of AA may have given the animals time to recover from toxic effects of parasitaemia and re-establish homeostasis.
Glucagon has a catabolic role on glycogen adjacent to the anabolic insulin action that tightly regulates blood glucose concentration (Schulman et al., 1957). We observed that liver glycogen stores were significantly higher in in the AA10, the NIC and the AC groups compared to either the IC and the CHQ controls (Figure 4). The glycogen levels correlated well with the glucagon activities in the relevant groups, being high where glycogen was low and being low where glycogen was high. This reciprocity was also evident in muscle glycogen concentrations showing that glucose homeostasis critically depended on the presence of AA10. However, it is now known that glucagon receptors are expressed in many tissues other than the liver showing extra influence of the hormone outside of glucose metabolism. One of these mechanisms is the involvement of glucagon in satiety initiation (Salter, 1960). We observed that animals in the well-fed disease-free groups (NIC, AC) had suppressed glucagon concentration and increased glycogen levels in either the liver or the muscles. The opposite was seen in the IC group where anorexia was apparent by reduced feeds and cachexic tendencies as shown by low muscle and liver glycogen concentrations.
Glucagon-induced hypophagia is regulated in the hypothalamus (Heppner et al., 2010) but only at low doses such that the high levels we saw in the acute studies (Table 1) and the sub-chronic studies may have initiated glucose mobilization regulating energy balance through thermogenesis and oxygen consumption (Davidson et al., 1957) by stimulation of brown adipose tissue (BAT) (Billington et al., 1991). In other normal conditions, this will have had a favourable effect on the animals’ wellbeing, but in malaria the disease process was promoted instead. Low insulin concentration is necessary for the pharmacological effects of glucagon as the hyperinsulinemia we saw in the IC group would invariably blunt glucagon thermogenic effects (Calles-Escandon, 1994). Administration of AA resulted in increased glucose utilization and may, therefore, utilize the glucagon-induced glucose mobilization consequently quenching thermogenesis. Thermogenesis and febrile response are, on the other hand, cardinal features of malaria which could be orchestrated by either glucagon or malaria glycosylphosphatidylinositol (GPI)-induced TNF-α secretion and other cytokines by acting on the hypothalamus in severe malaria (Schofield & Hackett, 1993). GPI moieties have pleiotropic effects in malaria one of which is insulin memetic action which regulates glucose utilization by adipocytes and causing profound hypoglycaemia (Schofield & Hackett, 1993, Krishnegowda et al., 2005).
Overall, the administration of AA10 may have had an influence on the insulin-glucagon axis through amelioration of the debilitating effects of malaria by suppression parasitaemia and inhibiting the subsequent sequelae of malaria or may have abated malaria inflammatory effects of malaria and emasculating the disease in the process. Either way, AA10 administration positively influenced glucose homeostasis as compared to infected non-treated or CHQ-treated animals.
Glucose utilization in malaria readily becomes dysregulated early in the disease course driven by over expression of glucose transporter-1 (GLUT-1) in virtually all the cells, which depletes plasma glucose autonomous of insulin activity. The over expression of GLUT-1 in the muscle is instituted through the influence of GPI on TNF-α increased secretion and subsequent over expression of inducible nitric oxide synthase (iNOS) and nitric oxide (NO) synthesis (Balon & Nadler, 1996, Bedard et al., 1997). While we did not investigate GPI interaction with AA or AA with GLUT-1 receptors, this may formulate gaps for future studies. Furthermore, the high concentrations of glycogen in the liver and the muscle may also point to an upregulations of GLUT 2 and GLUT 4 influenced by AA administration which may need to be investigated.
Plasma hyperlactataemia (Figure 6) observed in the IC group is evidence of increased energy demands of stress in the host or in the parasite or both and not necessarily hypoxia as lungs have been shown to produce increased amounts on lactate in disease (Opdam & Bellomo, 2000). Higher %parasitaemia was associated with higher levels of lactate production. Infection with malaria results in increased glucose production by approximately 50% (Davis et al., 1993) but also an expedited non-insulin mediated glucose disposal in glycolysis has been reported (Binh et al., 1997). This may mean possibly gluconeogenesis upregulation at the same time when glycolysis is increased. Therefore, the possibility that the intermediary molecule feeding both systems could be lactate is plausible. In sepsis, a condition that has many similarities with malaria infection, hyperlactaemia has been reported as an independent predictor of mortality (Garcia-Alvarez et al., 2014). We have also observed that administration of AA10 preserved the concentration of lactate at lower concentration close to those of the NIC showing that possibly the phytochemical is also able to inhibit futile glycolysis either through its antioxidant or anti-inflammatory activity and also gluconeogenesis. The preservation of glycogen in the AA10 administered animals may have been as a result of lactate conversion to the glucose storage unit or the high glycogen stores may have inhibited hyperlactaemia development, both facets may be attributable to the influence of AA administration on glucose homeostasis.
Conclusion
We have shown the novelty of AA administration in malaria influencing both acute and sub-chronic glucose homeostasis, the preservation of food and water intake and modulating hormonal glucose handling. Insulin and glucagon preserved their reciprocal relationship which is usually dysregulated in malaria and other forms of malaria treatments showing the anti-disease aspect of AA. The preservation of glucose in malaria is a novel finding for a phytochemical that has be known to be anti-hyperglycaemic in diabetes mellitus.
The link between AA and glucose homeostasis may be in that the phytochemical potentiates the activity of glycolytic enzymes creating a demand for glucose. This energy source deficit cannot be satisfied by glycogenolysis which was suppressed as shown by liver and muscle stores leaving exogenous food source as the only alternative for energy metabolism, initiating foraging habits. Lactate metabolism was normalised showing efficient lactate utilization through oxidation or gluconeogenesis. The satiation drive by glucagon was suppressed in the treated animals requiring higher concentrations for the same effect. The findings suggest that AA may be an antimalarial which preserves glucose homeostasis and therefore, may be an anti-disease phytotherapeutic with both anti-parasitic and anti-infection properties.
Acknowledgements
Our gratitude knows bounds to the veritable influence and knowledge Professor Cephas Tagumirwa Musabayane continues to have, even in death, on this work. We are highly indebted to Professor Fanie R Van Heerden for her insightful contributions to the conception and embankment of this project. Our thanks go to the Discipline of Physiology Endocrinology Group. Mr. M. Luvuno is highly appreciated for the tremendous contributions in animal and laboratory experiments.
Footnotes
Conflict of interest
The authors declare no conflict of in interest in this work.
References
- 1.Acquah S, Boampong J. N, Eghan Jnr BA, Eriksson M. Evidence of Insulin Resistance in Adult Uncomplicated Malaria: Result of a Two-Year Prospective Study. Malaria. Res. Treat. 20142014:8. doi: 10.1155/2014/136148. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 2.Agbenya T, Angus B. J, Bedu A. G, Baffoe B. B, Guyton T, Stacpoole P. W, Krishna S. Glucose and lactate kinetics in children with severe malaria. J. Clin. Endo. Metab. 2000;85:1569–1576. doi: 10.1210/jcem.85.4.6529. [DOI] [PubMed] [Google Scholar]
- 3.Balon T. W, Nadler J. L. Nitric oxide mediates skeletal glucose transport. Am. J. Physiol. 1996;33:E1058–E1064. doi: 10.1152/ajpendo.1996.270.6.E1058. [DOI] [PubMed] [Google Scholar]
- 4.Bedard S, Marcotte B, Marette A. Cytokines modulate glucose transport in skeletal muscle by inducing the expression of inducible nitric oxide synthase. Biochem. J. 1997;325:487–493. doi: 10.1042/bj3250487. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 5.Billington C. J, Briggs J. E, Link J. G, Levine A. S. Glucagon in physiological concentrations stimulates brown fat thermogenesis in vivo. Am. J. Physiol. 1991;261:R501–R507. doi: 10.1152/ajpregu.1991.261.2.R501. [DOI] [PubMed] [Google Scholar]
- 6.Binh T. Q, Davis T. M. E, Johnston W, Thu L. T. A, Boston R, Danh P. T, Anh T. K. Glucose metabolism in severe malaria: Minimal model analysis of the intravenous glucose tolerance test incorporating a stable glucose label. Metab. Clin. Exp. 1997;46:1435–1440. doi: 10.1016/s0026-0495(97)90144-x. [DOI] [PubMed] [Google Scholar]
- 7.Calles-Escandon J. Insulin dissociates hepatic glucose cycling and glucagoninduced thermogenesis in man. Metabolism. 1994;43:1000–1005. doi: 10.1016/0026-0495(94)90180-5. [DOI] [PubMed] [Google Scholar]
- 8.Cansu D, Korkmaz C. Hypoglycaemia induced by hydroxychloroquine in a non-diabetic patient treated for RA. Rheumatology (Oxford) 2008;47:378–379. doi: 10.1093/rheumatology/kem378. [DOI] [PubMed] [Google Scholar]
- 9.Changa K.-H, Stevenson M. M. Malarial anaemia: mechanisms and implications of insufficient erythropoiesis during blood-stage malaria. Internat. J. Parasitol. 2004;34:1501–1516. doi: 10.1016/j.ijpara.2004.10.008. [DOI] [PubMed] [Google Scholar]
- 10.Clark I. A, Budd A. C, Alleva L. M. Sickness behaviour pushed too far-the basis of the syndrome seen in severe protozoal, bacterial and viral diseases and post-trauma. Malar. J. 2008;7:208. doi: 10.1186/1475-2875-7-208. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 11.Clark I. A, Vissel B. Inflammation-sleep interface in brain disease: TNF, insulin, orexin. JNI. 2014;11:51. doi: 10.1186/1742-2094-11-51. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 12.Croft A. Malaria affects children and pregnant women most. BMJ. 2000;321:1288. [PMC free article] [PubMed] [Google Scholar]
- 13.Davidson I. W, Salter J. M, Best C. H. Calorigenic action of glucagon. Nature. 1957;180:1124. doi: 10.1038/1801124a0. [DOI] [PubMed] [Google Scholar]
- 14.Davis T. M, Looareesuwan S, Pukritayakamee S, Levy J. C, Nagachinta B.N.J. W. Glucose turnover in severe falciparum malaria. Metabolism. 1993;42:334–340. doi: 10.1016/0026-0495(93)90083-z. [DOI] [PubMed] [Google Scholar]
- 15.Elbadawi N. E. E, Mohamed M. I, Dawod O. Y, Ali K. E, Daoud O. H, Ali E. M, Ahmed E. G. E, Mohamed A. E. Effect of quinine therapy on plasma glucose and plasma insulin levels in pregnant women infected with Plasmodium falciparum malaria in Gezira state. East Mediterrenean Health J. 2011;17:697–700. [PubMed] [Google Scholar]
- 16.English M, Wale S, Binns G, Mwangi I, Sauerwein H, Marsh K. Hypoglycaemia on and after admissionin Kenyan children with severe malaria. Q. J. Med. 1998;91:191–197. doi: 10.1093/qjmed/91.3.191. [DOI] [PubMed] [Google Scholar]
- 17.Fen^ndez-Real J. M, Ricart W. Insulin resistance and inflammation in an evolutionary perspective: the contribution of cytokine genotype/phenotype to thriftiness. Diabetologia. 1999;42:1367–1374. doi: 10.1007/s001250051451. [DOI] [PubMed] [Google Scholar]
- 18.Garcia-Alvarez M, Marik P, Bellomo R. Sepsis-associated hyperlactatemia. Critical Care. 2014;18:503. doi: 10.1186/s13054-014-0503-3. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 19.Goldring J. P. Evaluation of immunotherapy to reverse sequestration in the treatment of severe Plasmodium falciparum malaria. Immunol. Cell Biol. 2004;82:447–452. doi: 10.1111/j.0818-9641.2004.01265.x. [DOI] [PubMed] [Google Scholar]
- 20.Grimble R. Dietary manipulation of the inflammatory response. Proc. Nutri. Soc. 1992;51:285–294. doi: 10.1079/pns19920039. [DOI] [PubMed] [Google Scholar]
- 21.Hart B. L. Biological basis of the behavior of sick animals. Neurosci. Biobehav. Rev. 1988;12:23–137. doi: 10.1016/s0149-7634(88)80004-6. [DOI] [PubMed] [Google Scholar]
- 22.Heppner K. M, Habegger K. M, Day J, Pfluger P. T, Perez-Tilve D, Ward B, Gelfanov V, Woods S. C, DiMarchi R, Tschöp M. Glucagon regulation of energy metabolism. Physiology Behavior. 2010;100:545–548. doi: 10.1016/j.physbeh.2010.03.019. [DOI] [PubMed] [Google Scholar]
- 23.Huang S.-S, Chiu C.-S, Chen H.-J, Hou W.-C, Sheu M.-J, Lin Y.-C, Shie P.-H, Huang G.-J. Antinociceptive Activities and the Mechanisms of Anti-Inflammation of Asiatic Acid in Mice. Evid-Based Complemen. Altern. Med 2011. 2011:10. doi: 10.1155/2011/895857. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 24.Krishnegowda G, Hajjar A. M, Zhu J, Douglass E. J, Uematsu S, Akira S, Woods A. S, Gowda D. C. Induction of proinflammatory responses in macrophages by the glycosylphosphatidylinositols of Plasmodium falciparum: cell signaling receptors, glycosylphosphatidylinositol (GPI) structural requirement, and regulation of GPI activity. J. Biol. Chem. 2005;280:8606–8616. doi: 10.1074/jbc.M413541200. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 25.Mackintosh C. L, Beeson J. G, Marsh K. Clinical features and pathogenesis of severemalaria. Trends in Parasitol. 2004;20:597–603. doi: 10.1016/j.pt.2004.09.006. [DOI] [PubMed] [Google Scholar]
- 26.Mavondo G.A, Mkhwanazi B.N, Mabandla M.V. Pre-infection administration of asiatic acid retards parasitaemia induction in Plasmodium berghei murine malaria infected Sprague?Dawley rats. Malar. J. 2016;15:226. doi: 10.1186/s12936-016-1278-6. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 27.Miller L. H, Ackerman H. C, Su X.-z, Wellems T. E. Malaria biology and disease pathogenesis: insights for new treatments. Nature Med. 2013;19:156–167. doi: 10.1038/nm.3073. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 28.Moussaieff A, Rouleau M, Kitsberg D, Cohen M, Levy G, Barasch D, Nemirovski A, Shen-Orr S, Laevsky I, Amit M, Bomze D, Elena-Herrmann B, Scherf T, Nissim-Rafinia M, Kempa S, Itskovitz-Eldor J, Meshorer E, Aberdam D, Nahmias Y. Glycolysis-mediated changes in acetyl-CoA and histone acetylation control the early differentiation of embryonic stem cells. Cell Metab. 2015;21:392–402. doi: 10.1016/j.cmet.2015.02.002. [DOI] [PubMed] [Google Scholar]
- 29.Murambiwa P, Tufts M, Mukaratirwa S, van Heerden F R, Musabayane C. T. Evaluation of efficacy of transdermal delivery of chloroquine on Plasmodium berghei-infected male Sprague-Dawley rats and effects on blood glucose and renal electrolyte handling. Endocrine Abstracts. 2013;13:P203. [Google Scholar]
- 30.Musabayane C. T, Murambirwa P, Joosab N, Masola B, Mukaratirwa S. The effects of chloroquine on blood glucose and plasma insulin concentrations in male Sprague Dawley rats. Soc. Endocrinol. 2010;21:139. [Google Scholar]
- 31.Olszewski K. L, Morrisey J. M, Wilinski D, Burns J. M, Vaidya A. B, Rabinowitz J. D, Llinas M. Host-Parasite Interactions Revealed by Plasmodium falciparum Metabolomics. Cell Host Microbe. 2009;5:191–199. doi: 10.1016/j.chom.2009.01.004. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 32.Opdam H, Bellomo R. Oxygen consumprion and lactate release by the lung after cardiopulmonary bypass and during septic shock. Crit. Care Resusc. 2000;2:181–187. [PubMed] [Google Scholar]
- 33.Osier F. H, Berkley J. A, Ross A, Sanderson F, Mohammed S, Newton C. R. Abnormal blood glucose concentrations on admission to a rural Kenya district hospital: prevalence and outcome. Arch. Dis. Child. 2003;88:621–625. doi: 10.1136/adc.88.7.621. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 34.Pedersen P.L. Warburg, me and Hexokinase 2: Multiple discovoeris of key molecular evevents underlying of cancers’ most common phenotypes, the “Warbug Effect”, i.e, elevated gylcolysis in the presence of oxygen. J. Bioenerg. Biomembr. 2007;39:211–222. doi: 10.1007/s10863-007-9094-x. [DOI] [PubMed] [Google Scholar]
- 35.Ramachandran V, Saravanan R. Efficacy of asiatic acid, a pentacyclic triterpene on attenuating the key enzymes activities of carbohydrate metabolism in streptozotocininduced diabetic rats. Phytomedicine. 2013;20:230–236. doi: 10.1016/j.phymed.2012.09.023. [DOI] [PubMed] [Google Scholar]
- 36.Ramachandran V, Saravanan R. Antidiabetic and antihyperlipidemic activity of asiatic acid in diabetic rats, role of HMG CoA: in vivo and in silico approaches. Phytomedicine. 2014;21:225–232. doi: 10.1016/j.phymed.2013.08.027. [DOI] [PubMed] [Google Scholar]
- 37.Rangaraj N, Vaghasiya K, Jaiswal S, Sharma A, Shukla M, Lalb J. Do Blood Sampling Sites Affect Pharmacokinetics? Chemist Biol. Interface. 2014;4:176–191. [Google Scholar]
- 38.Salter J. M. Metabolic effects of glucagon in the Wistar rat. Am. J. Clin. Nutr. 1960;8:535–539. [Google Scholar]
- 39.Schofield L. Rational approaches to developing an anti-disease vaccine against malaria. Microbes Infect. 2007;9:784–791. doi: 10.1016/j.micinf.2007.02.010. [DOI] [PubMed] [Google Scholar]
- 40.Schofield L, Hackett F. Signal Transduction in Host Cells by a Glycosylphosphatidyllnositol Toxin of Malaria Parasites. J. Exp. Med. 1993;177:145–153. doi: 10.1084/jem.177.1.145. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 41.Schulman J. L, Carleton J. L, Whitney G, Whitehorn J. C. Effect of glucagon on food intake and body weight in man. J. Appl. Physiol. 1957;11:419–421. doi: 10.1152/jappl.1957.11.3.419. [DOI] [PubMed] [Google Scholar]
- 42.Seifter S, Dayton S, Novic B, Muntwyler E. The estimationof glycogen with the athrone reagent. Federation Proc. 1949;8:249. [Google Scholar]
- 43.Warburg O. The metabolism of tumours. London: Constable Co. LTD 1930; 1930. [Google Scholar]
- 44.Warburg O. Science. 1956;124:269–270. [PubMed] [Google Scholar]
- 45.White N.J, Warrell D. A, Chanthanavich P, Looareesuwan S, Warrell M. J, Krishna S. Severe hypoglycaemia and hyperinsulinaemia in falciparum malaria. N. Engl. J. Med. 1983;309:61–62. doi: 10.1056/NEJM198307143090201. [DOI] [PubMed] [Google Scholar]
- 46.Ende Zhao Tomasz, Maj Ilona, Kryczek Wei, Li Ke, Wu Lili, Zhao Shuang, Wei Joel, Crespo Shanshan, Wan Linda, Vatan Wojciech, Szeliga Irene, Shao Yin, Wang Yan, Liu Sooryanarayana, Varambally Arul M, Chinnaiyan Theodore H, Welling Victor, Marquez Jan, Kotarski Hongbo, Wang Zehua, Wang Yi, Zhang Rebecca, Liu Guobin Wang, Weiping Zou. Cancer mediates effector T cell dysfunction by targeting microRNAs and EZH2 via. glycolysis restriction. Nature Immunol. 2016;17:95–103. doi: 10.1038/ni.3313. [DOI] [PMC free article] [PubMed] [Google Scholar]