
Fig. 1.
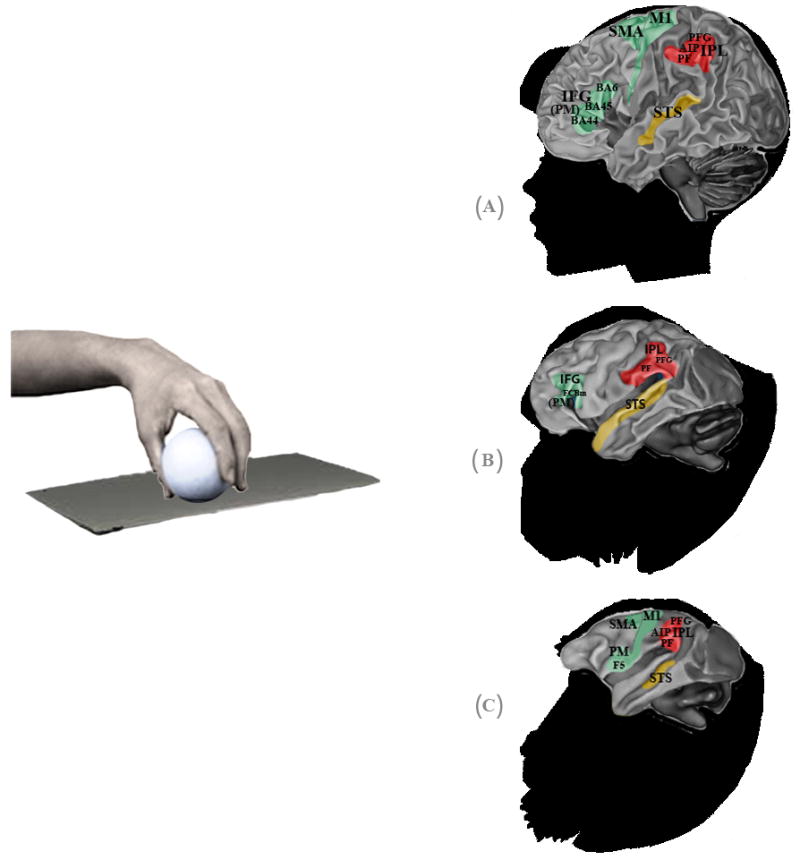
Hand mirror neurons (MNs) in humans, chimpanzees and macaques. Specific anatomical regions of human (A), chimpanzee (B) and macaque (C) brains activated during observation of transitive grasping that overlap with regions activated during the execution of the same grasping action. Premotor, parietal and sensory areas are shown in green, red and yellow, respectively. In humans, the premotor cortex (PM) and inferior frontal gyrus (IFG) approximate Brodman areas BA44 (known as Broca’s area), BA45 and BA6, while the rostral part of the inferior parietal lobule (IPL) is thought to correspond to cytoarchitectonic areas PF and PFG. In the parietal area, mirror activity has also been found in the anterior sector of the intraparietal sulcus (AIP). In chimpanzees, the IFG occupies the anatomical region FCBm, following Bailey et al. (1950); while in the IPL, mirror responses occupy areas PF and PFG. In macaques, the PM contains area F5, where mostly hand MNs are located, and in the IPL MNs have been found in areas PF, PFG and in the AIP region. In humans and macaques, hand mirror responses have also been found in the motor cortex (M1) and supplementary motor area (SMA). The superior temporal sulcus (STS) is a sensorial region which lacks motor properties but is considered part of the hand mirror neuron system, because it is thought to be the main source of visual information.