

Abstract
Type 2 diabetes (T2D) is increasing worldwide, making identification of biomarkers for detection, staging, and effective prevention strategies an especially critical scientific and medical goal. Fortunately, advances in metabolomics techniques, together with improvements in bioinformatics and mathematical modeling approaches, have provided the scientific community with new tools to describe the T2D metabolome. Among the metabolomics signatures associated with T2D and obesity include increased levels of lactate, glycolytic intermediates, branched-chain and aromatic amino acids, and long-chain fatty acids. Conversely, tricarboxylic acid cycle intermediates, betaine and other metabolites decrease. Future studies will be required to fully integrate these and other findings into our understanding of the diabetes pathophysiology and to identify biomarkers of disease risk, stage, and responsiveness to specific treatments.
Keywords: Metabolomics, Type 2 diabetes, Insulin resistance, Insulin secretion
Introduction
While type 2 diabetes mellitus (T2D) is defined by elevations in plasma glucose levels, it is not simply a disorder of carbohydrate metabolism. Alterations in glucose, lipid, and protein metabolism accompany insulin resistance and disordered pancreatic β-cell function, and are present years before the disease is clinically diagnosed (1–3). Given the increasing prevalence of T2D worldwide (4), it is especially important to identify the earliest, potentially pathogenic disruptions which confer disease risk in order to improve approaches to treatment.
More than 40 years ago, Pauling and Robinson postulated that the functional status of a biological system was reflected in the pattern of metabolites in biological fluids (5), now termed the metabolome. With the recent advent of high-throughput and comprehensive approaches to quantify the plasma and tissue metabolome, this hypothesis has been repeatedly tested and verified. In this review, we will summarize studies of the metabolome related to the pathophysiology and identification of biomarkers of T2D and obesity.
Techniques for Analyzing the Metabolome
Analysis of the metabolome commonly utilizes mass spectrometry (MS) and/or nuclear magnetic resonance (NMR) techniques (reviewed in (6)). Samples are typically separated using gas (GC) or liquid chromatography (LC) prior to MS; and compounds are identified by mass and fragmentation patterns. By contrast, NMR spectroscopy measures all the metabolites at once, with no need for separation, but this method is less sensitive (7, 8). For targeted metabolomics, stable isotope-labeled standards are added to the sample prior to extraction in order to quantify absolute concentrations of a limited number of known metabolites (7–9). Conversely, untargeted metabolomics approaches can detect and determine relative concentrations of a large number of metabolites by measuring the differences between two experimental conditions (7, 9). Untargeted approaches are limited by incomplete spectral and chemical libraries, leading to incomplete identification of all spectral peaks (7).
Glycolysis and TCA Cycle Metabolites in T2D
Not surprisingly, plasma hexose sugars are positively associated with T2D, and largely accounted for by disease-defining elevations in glucose (10). Nevertheless, elevations in other carbohydrates, such as fructose, mannose, and sorbitol, and related compounds such as lactate and malate, have been consistently associated with T2D (10–16). Changes in plasma lactate during an oral glucose tolerance test (OGTT) are inversely correlated with fasting insulin, indicating that insulin resistance can be reflected through the metabolite’s response to a glucose challenge (17).
Intracellular metabolite profiles also indicate perturbations in glycolytic and tricarboxylic acid (TCA) cycle metabolism in multiple tissues. For example, glucose-stimulated insulin secretion (GSIS) requires glucose-stimulated increases in late glycolytic and early TCA cycle metabolites; these metabolites are unchanged in clonal lines lacking GSIS (18). 13C-NMR studies have also revealed a role for altered pyruvate cycling in lipid-induced impairments in GSIS (19, 20).
Similar defects in metabolism are characteristic of insulin resistant muscle. For example, lipid-induced insulin resistance results in reduced pyruvate dehydrogenase (PDH) activity (21) and incomplete fatty acid (FA) oxidation, and is accompanied by decreases in downstream TCA cycle and electron transport chain (ETC) metabolites (22, 23). TCA cycle intermediates are also decreased in liver and skeletal muscle of genetically obese mice, in parallel with increased glycolytic intermediates in muscle (24). Muscle from high-fat diet (HFD)-fed mice have decreased labelling of TCA intermediates during glucose tracing experiments (25). Distinct patterns have been observed in adipose from obese mice, with increases in TCA cycle metabolites in ob/ob and similar trends in db/db (24).
Metabolomics-based studies of urine samples have provided more contradictory data. Urinary α-ketoglutarate levels are decreased in obese vs. lean individuals (26). Additionally, decreases in urinary fumarate and succinate contribute to the differentiation of patients with T2D from healthy individuals in a principal component analysis (27). By contrast, urinary succinate and lactate are increased, and α-ketoglutarate decreased, in rodents with genetic obesity and T2D (db/db mice, fa/fa rats). Similarly, NMR metabolomics analysis reveals increased urinary glucose, lactate and several TCA intermediates in diabetic mice (28).
Differences between studies could arise from differences in platform, species, gender, urinary excretion patterns for specific metabolites, and/or the experimental model used. Nevertheless, alterations in TCA cycle intermediates provide support for the central role of this pathway as a marker, and perhaps mediator, of perturbed energetic metabolism in the insulin resistant state. In turn, such defects may be related to impaired anaplerotic flux (12, 29), potentially due to incomplete FA oxidation (23) and amino acid metabolism (26, 30), as described below.
Amino acids and derivative metabolites
Increases in plasma levels of amino acids, particularly branched-chain and aromatic amino acids (BCAA and AAA), have long been recognized as a marker of obesity-associated insulin resistance (31). Similar patterns have been confirmed in more recent metabolomics analyses (26, 32, 33). Additional amino acids are also elevated in obesity e.g. alanine, phenylalanine, tyrosine, glutamate/glutamine, aspartate/asparagine, and arginine (26). In a Japanese population (34), alanine, glutamate, tryptophan, tyrosine and BCAA were positively correlated with visceral adiposity, while glycine was inversely correlated.
Amino acid metabolites are also perturbed in insulin resistance and T2D. An untargeted study demonstrated that the branched-chain ketoacid 3-methyl-2-oxovalerate strongly predicted impaired fasting glucose (IFG) (16). In this same study, other branched-chain ketoacids (4-methyl-2-oxopentanoate and 3-methyl-2-oxobutyrate) and the downstream BCAA metabolic products propionylcarnitine, alpha-methylbutyrylcarnitine and isovalerylcarnitine were also elevated in IFG and T2D. Another product of amino acid catabolism, α-hydroxybutyrate, is also increased in insulin resistance, potentially reflecting oxidative stress (35).
Plasma levels of BCAA and AAA also predict risk for developing T2D in healthy individuals (32, 33). High levels of three amino acids (isoleucine, phenylalanine and tyrosine) were associated with a five- to seven-fold increased risk of developing diabetes in one cohort, and a similar two-fold risk in a second case-control cohort (33). Interestingly, fasting concentrations of these amino acids were already elevated as early as 12 years before the onset of T2D (33). Alanine has also been associated with hyperglycemia and T2D risk (36–38).
Conversely, other amino acids are reduced in insulin resistance. Low glycine is associated with insulin resistance (36) and predicts both IGT and T2D (39); these associations may be related to glycine’s role as an upstream precursor for glutathione synthesis and antioxidant responses (40). Glutamine levels are also reduced in insulin resistance (38); higher levels of glutamine or the glutamine-to-glutamate ratio are associated with lower risk of incident diabetes, even after adjustment for BMI and BCAA (32, 36). Indeed, supplementation of mice with glutamine improves glucose tolerance and reduces blood pressure (36). Similarly plasma levels of the methyl donor betaine are reduced in individuals with insulin resistance (41). Betaine may be particularly important for hepatic lipid metabolism, as betaine supplementation of mice with diet-induced obesity reduces hepatic lipid and improves whole-body metabolism via FGF21-dependent mechanisms (41).
Amino acids may also serve as biomarkers for response to medical therapies for established T2D (42) or preventive strategies (41, 43). BCAA and AAA decrease with sulfonylurea therapy, and after gastric bypass (44), but surprisingly increase in response to metformin (42). Plasma betaine levels not only increase in response to lifestyle and pharmacologic therapies, but also predict their success (43).
Taken together, multiple studies consistently report that amino acids are altered in insulin resistance and predict T2D. Indeed, amino acid metabolites add predictive value to standard clinical measures. For example, the odds of developing future diabetes are increased by 57% to 102% for each SD increment in a panel of 5 BCAA and AAA (33). Such increases in relative risk are comparable to or higher than risk associated with BMI, increased age, fasting glucose, or genetic polymorphisms linked to T2D risk (45).
While amino acids are consistently increased in T2D, it remains uncertain whether these patterns simply result from insulin resistance and reduced inhibition of proteolysis, or from increased dietary intake or absorption. Regardless, such perturbations in amino acids may also contribute to the onset of overt diabetes by reducing both insulin action and secretion (30, 46–49). The cellular and tissue mechanisms responsible for these mechanisms remain uncertain. BCAA can modulate cellular insulin action (50, 51), stimulating Tor-dependent pathways while inhibiting proximal steps in insulin signaling. Whether specific amino acids have unique roles remains uncertain, as rodent studies of BCAA supplementation have yielded discordant data. For example, leucine supplementation produced beneficial metabolic effects in one study (52), while supplementation with multiple BCAA typically induces insulin resistance in parallel with increases in acylcarnitines (26). These data formed the basis of the “BCAA overload” hypothesis, which posits that BCAA contribute to insulin resistance and T2D in the context of HFD-feeding (26, 53). Moreover, HFD-fed mice null for the BCAT2 gene, which encodes the first enzyme in BCAA catabolism, have improved glucose homeostasis and insulin sensitivity, indicating that products of BCAA oxidative metabolism may confer insulin resistance. Further studies will be required to clarify these possibilities.
The Lipid Metabolome
Given the importance of obesity as a risk factor for T2D, it is not surprising that adipose-derived metabolites are increased in insulin resistant individuals. Free fatty acids (FFAs, also known as non-esterified fatty acids or NEFAs) are released from adipose as a consequence of increased lipolysis when levels or action of insulin are reduced, such as with fasting, obesity, and insulin resistance. Elevations in plasma FFAs in turn promote increased tissue uptake of lipids and ectopic deposition of triglycerides (TG) in liver and muscle. Other lipids or metabolic byproducts such as diacylglycerides, ceramides, acylcarnitines, and reactive oxygen species (ROS) are proposed to mediate adverse effects on both insulin signaling and metabolism (54–61). Collectively, these intermediates may ultimately lead to reduced insulin-stimulated glucose uptake in muscle, increased hepatic glucose production in liver, and β-cell dysfunction.
Not all lipid species have detrimental effects on whole-body metabolism. For example; omega-3 FA, palmitoleate, and other unsaturated FA have beneficial effects (62–66). Specific FA constituents of circulating TG also confer differential risk for T2D. For example, LC/MS analysis of fasting plasma from individuals at risk for T2D indicates that TG of lower carbon number and double bond content (i.e. saturated and monounsaturated species) are associated with increased T2DM risk, whereas TG of increased carbon number and double bond content are associated with decreased risk (65). Moreover, insulin resistant patients have a blunted decrease in TG with lower carbon number during OGTT.
Novel lipids have recently been identified as both plasma biomarkers and contributors to T2D pathophysiology. An early GC/MS-based study by Newgard and colleagues revealed that five FFA were increased in obesity (C14:0, C16:1, C16:0, C18:1, C20:4), while the polyunsaturated linoleic and linolenic acids (C18:3 and C18:2) were unchanged (26). Lipids with even longer chain length (adrenate and arachidonate, C22:4 and C20:4) are elevated in IFG (16). Conversely, some lipid species, such as heptanoate, pelargonate and 5-dodecenoate (C7:0, C9:0 and 12:1) are decreased in T2D (16). Moreover, both FAs and by-products of FA oxidation (C14:0, C16:1, C16:0, alpha-C18:3. C18:2, C18:1, C18:0, C22:6, hydroxybutyrate and other ketones) are inversely correlated with acute insulin secretion (30).
New lipid candidates have recently been identified. Yore and colleagues (67) reported novel lipids with a branched ester linkage between a FA and a hydroxyl-FA, termed fatty-acid-hydroxy FA (FAHFA). Sixteen FAHFAs family members were identified, with palmitic-acid-9-hydroxy-stearic acid (9-PAHSA) being the most upregulated in adipose from GLUT4-null mice (67). However, PAHSA was decreased in serum and adipose from insulin resistant humans and mice. Such reductions in PAHSA could impact on key facets of metabolism, as this lipid can increase GSIS in human islets, increase insulin-stimulated glucose transport via GPR120 and GLUT4 translocation, and exert anti-inflammatory effects in vivo. Thus, reduced PAHSA may contribute to T2D risk (67).
Acylcarnitines have also been studied as T2D predictive biomarkers given their key roles in both lipid and amino acid metabolism, including transport of long-chain FAs into the mitochondria for oxidation. Newgard and colleagues reported that 4 of 37 acylcarnitines were increased in obese humans (C3, C5, C6, C8:1)(26). Moreover, the combination of C3 and C5 acylcarnitines, together with BCAAs and AAAs, methionine, and glutamate/glutamine, was particularly most robust for differentiating lean from obese patients. Since C3 and C5 acylcarnitines accumulate to a greater extent with BCAA supplementation, these may reflect BCAA catabolism. Huffman and colleagues also found elevations in some medium-chain acylcarnitines (C8:1, C10:3), which were inversely associated with disposition index (30). By contrast, decanoylcarnitine (C10) and the lysophospholipid 1-lineoylglycerophosphocholine were decreased in and predictive of insulin resistance in another study (35).
Genome links with the metabolome
Complex interactions between many factors, both genetic and environmental, converge to influence the metabolite profile of a given individual. Recent studies have utilized genome-wide association studies (GWAS) to examine the impact of genetic variation on plasma metabolites. For example, one GWAS study of 163 metabolites identified genetic variants linked to metabolite levels, or to ratios of enzymatic product to substrates (68). The majority of validated loci were located within or near genes encoding enzymes or carriers, including many related to β-oxidation, FA and phospholipid biosynthesis, and amino acid metabolism. Interestingly, these loci explained up to 36% of the variance in metabolites, pointing to an important role for genetics. Similarly, genetic variants associated with glycine, serine, and betaine levels have been identified (69); but these variants were not clearly linked to diabetes-related traits.
Environmental factors affecting the metabolome
Beyond disease state and genetic background, additional environmental factors such as diet, activity, medication, age, developmental history, and the microbiome influence the metabolome (70). Metabolomics profiles can be influenced by dietary macronutrients, and thus are highly valuable for studies of nutritional epidemiology (71); an estimated one-third of all compounds associated with a given nutritional pattern can be linked to environmental factors. Plasma metabolites are also highly influenced by acute or prolonged exercise, with increased levels of glucose-6-phosphate (indicator of glycogenolysis); succinate, malate and fumarate (TCA-related); glycerol (lipolysis product); panthothenate (related to FA oxidation); and niacinamide. Moreover, β-aminoisobutyric acid (BAIBA) concentrations are increased with exercise and inversely related to markers of metabolic risk (72). Metabolomic patterns can also be influenced by gender (30) and age (73). Collectively, these studies highlight the importance of considering differences in subjects’ diet, physical activity, gender, and age as potential confounding factors for metabolomics study design and data interpretation.
The composition of the gut microbiome is increasingly recognized as an important contributor to both host systemic metabolism and metabolomics profiles (74–76). In turn, microbiota composition is influenced by the host genome, dietary macronutrients, activity, and antibiotic use (especially during early life). For example, obesity and T2D are characterized not only by altered microbial profiles but also reduced microbial diversity (77–79), although differences in diversity are still controversial (80, 81). Microbiome profiles can be profoundly modified in response to bariatric surgery-induced weight loss, paralleling changes in bile acid profiles (82–84). Whether these altered microbe populations actually contribute to, or are a consequence of disease, is difficult to address in humans. Differences in microbial populations can modulate host intestinal development, secretion of incretin peptides regulating insulin secretion and whole-body metabolism, and intestinal inflammation (85), thus contributing to impaired glucose homeostasis, insulin resistance, and non-alcoholic fatty liver disease (NAFLD) (86). Perhaps the most convincing are microbiota transplantation studies; germ-free mice transplanted with obese microbiota gain more weight than recipients of microbiota from lean mice (87, 88). Moreover, transplantation of microbiota from post-bariatric surgical patients results in reduced fat mass in germ-free recipient mice (89). Human data are more limited, but insulin sensitivity improved in patients with metabolic syndrome after transplantation of healthy lean microbiota (90).
Identification of the specific microbes or microbial metabolites responsible for host metabolic regulation is an important goal. The microbiome of Western diet-fed mice produces more end-products of bacterial fermentation, such as lactate, acetate and butyrate (88). Bacteria which produce butyrate and those which alter bile acid composition may be particularly important modulators of insulin sensitivity and diabetes risk. The relative primacy of host genotype vs. environmental effects on the microbiome and related metabolite profiles remains uncertain at this time.
Another metabolite which has drawn attention to the role of the microbiota in the systemic metabolome is trimethylamine N-oxide (TMAO), an oxidized derivative of trimethylamine (TMA) produced by gut microbial metabolism. Plasma levels of choline, betaine and TMAO are strongly correlated with cardiovascular disease (CVD) (91). Moreover, increased TMAO has also been linked to increased cardiovascular risk (92), accelerated atherosclerosis (93), and heart failure in rodents (94).
Modelling approaches to integrating metabolomics data
Pathway mapping and visualization tools have become integral to the analysis of metabolomics experiments (reviewed in (95)). Systems biology tools that model metabolic networks have made in silico studies of metabolic disease feasible. Gene expression and metabolomics data can be incorporated to generate causal networks that map to genetic regions correlating with metabolite levels (96). Tissue-specific models have been developed to predict metabolic responses to hormones and nutrients; the predictions have correlated with known biomarkers and experimentally measured metabolites (97–101).
Flux balance analysis (FBA) has been used to model muscle metabolism and the impact of increased substrate availability (as observed in insulin resistance) in order to identify enzymatic reactions that may contribute to disease risk (100). Using this approach, Nogiec and colleagues found that perturbations in a single enzyme were insufficient to mimic insulin resistant metabolism e.g. decreased ATP synthesis, TCA flux, and metabolic inflexibility. However, combined inhibition of two enzymes, such as PDH and electron-transferring-flavoprotein dehydrogenase (ETFDH), was able to replicate insulin resistant phenotypes, indicating that defects in multiple pathways may be required to prevent compensatory effects and impact net metabolic flux (100). An integrated multi-tissue model, which was developed to simulate the metabolic interdependence of organs in diabetes, found differentially expressed reactions which could yield observed increases in FFA and lactate (99). Further technical improvements in computational models are needed to extend these findings and fully integrate metabolomics with transcriptomics and proteomics data sets.
Metabolomic studies in pregnancy and childhood
Metabolomic analysis of maternal or infant biofluids (e.g. plasma, urine, amniotic fluid, breast milk) provides valuable insights into the metabolic status of the mother and the developing child, and thus may be a promising strategy for identifying women at risk for adverse pregnancy outcomes, and infants at increased risk for future metabolic disease. Metabolomic analysis has shown promise for the early detection of pregnancy complications (e.g. preterm delivery, pre-eclampsia, intrauterine growth restriction (IUGR), fetal macrosomia, gestational diabetes (GDM), etc.) (102–107), but is not yet widely used beyond the research setting.
Altered maternal lipid metabolism is a potential biomarker for at-risk pregnancies. For example, increased maternal glycerol in the first trimester predicts subsequent pre-eclampsia (107), while lower levels of phospholipids, lysolipids, monoacylglyerols and FA binding protein predict fetal macrosomia (105). Small for gestational age (SGA) can be predicted by a panel of maternal metabolites assayed at 15 weeks of pregnancy, including sphingolipids, phospholipids, and carnitines (108). Other groups have described associations between lower maternal urinary acetate, tyrosine, and TMA levels during the first trimester and subsequent risk of SGA (103). The mechanisms responsible for these associations remain uncertain.
Gestational diabetes (GDM) is a strong risk factor for the subsequent development of overt diabetes in mothers, and for childhood obesity and diabetes in offspring. Not surprisingly, metabolomic analyses in GDM have identified signatures similar to other insulin resistant states, including elevations in NEFA and ketones (109–114). Elevations in FA, cholesterol, and lipoproteins may precede and predict GDM (115). Increased BCAA are identified in some GDM studies (110, 111), but not all, perhaps due to differences in timing of sample collection (116, 117). The fatty acid metabolite 3-carboxy-4-methyl-5-propyl-2-furanpropanoic acid (CMPF) (118) is also increased in GDM and may potentially contribute to β-cell dysfunction, via impaired mitochondrial function and oxidative stress. Infants of mothers with GDM have reduced cord blood glucose (consistent with neonatal hyperinsulinemia), together with increases in pyruvate, amino acids, ornithine, and the BCAA metabolite alpha-ketoisovaleric acid (116, 119).
Low birth weight resulting from either prematurity or intrauterine growth restriction is associated with increased risk of insulin resistance and diabetes in later life (120, 121); identification of metabolic patterns associated with risk would be helpful to target preventive strategies. Studies of cord blood metabolites have revealed lower levels of glucose, glutamine, alanine, and proline, and higher phenylalanine and citrulline in low birth weight infants (122–124) than controls. Accelerated postnatal weight gain, or “catch-up growth”, which often follows intrauterine growth restriction, independently increases adult risk of diabetes and obesity (125, 126). In a nested case-control study of cord blood from neonates with accelerated postnatal weight gain and early-childhood obesity vs. controls with normal postnatal growth, Isganaitis et al. reported reduced tryptophan and one-carbon metabolism-related metabolites, and reduced glutamine:glutamate ratio (127). These findings parallel results from insulin resistant adults (36). Other groups have observed positive associations between accelerated postnatal weight gain and plasma LPC14:0 and glutamine, and inverse associations with urine myoinositol (128, 129).
Childhood metabolome and disease risk
Several groups have reported that the plasma metabolome is altered in obese vs. lean children (130–132), but patterns are distinct from adults. For example, increased levels of acylcarnitines are found in adult obesity, yet variable in children (131, 133, 134). Some, but not all, reports indicate increased BCAA in obese and/or insulin resistant children and adolescents (130, 131, 133, 135). Such differences may reflect the impact of age, growth, pubertal stage, or the longitudinal changes in insulin secretion that precede T2D onset.
SUMMARY and CONCLUSIONS
In summary, advances in technical and computational methods to assess the metabolome have been invaluable for the investigation of the pathogenesis of T2D and related risk factors. Results from these studies (Figure 1) have consistently highlighted perturbations in both amino acid and lipid metabolism, as evidenced by increased BCAA and AAA, long-chain FAs, and short and odd-chain acylcarnitines. In parallel, glycolytic intermediates are increased, and TCA cycle intermediates are decreased. Collectively, these data support the hypothesis that impaired amino acid and FA metabolism could contribute to reduced anaplerotic flux and complete oxidative metabolism, thus contributing to reduced cellular energetics in insulin resistance and T2D. Emerging data also indicate that the gut microbiome is an important contributor to host metabolism. However, additional studies will be required to define the contribution of host vs. microbiome metabolism to specific metabolomics patterns, and identify the species and concrete mechanisms responsible for these important interactions. Development of expanded computational techniques to integrate transcriptomic and proteomic data with metabolomics profiles, and to model cellular and interorgan flux, will be essential for improving our understanding of metabolic pathophysiology associated with T2D risk. From a clinical perspective, understanding mechanisms responsible for altered metabolomics profiles will be essential for identifying specific biomarkers to guide improved prediction, prevention, and treatment of T2D.
Figure 1.
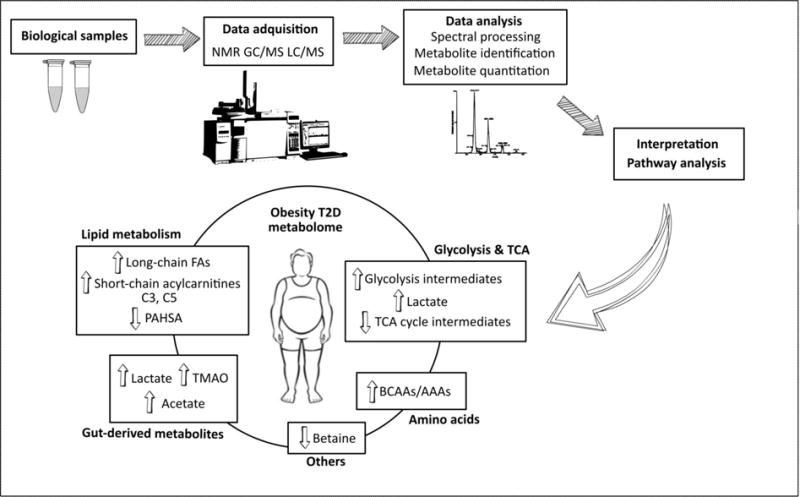
Obesity and T2D-related metabolome. Metabolomics experimental workflow from biological samples to interpretation of the results. NMR: nuclear magnetic resonance; GC: gas chromatography; LC: liquid chromatography; MS: mass spectrometry; T2D: type 2 diabetes; FA: fatty acid; TCA: tricarboxylic acid; TMAO: trimethylamine N-oxide; BCAA: branched-chain amino acid; AAA: aromatic amino acid.
Acknowledgments
AG-F gratefully acknowledges support from the American Diabetes Association (mentored research fellowship to MEP). AB acknowledges research support from the NIH and the Harold Whitworth Pierce Charitable Trust Postdoctoral Fellowship (T32DK007260). EI acknowledges grant support from K99R00 HD064793 and Boston Nutrition and Obesity Research Center. MEP acknowledges research support from the NIH, American Diabetes Association, Bristol-Myers Squibb, Janssen, Nuclea, Medimmune, and Joslin DRC grant P30 DK036836.
Footnotes
Conflict of Interest:
AG-F, AB, EI, and MEP declare no conflict of interest related to this manuscript.
References
- 1.DeFronzo RA. Banting Lecture. From the triumvirate to the ominous octet: a new paradigm for the treatment of type 2 diabetes mellitus. Diabetes. 2009;58(4):773–95. doi: 10.2337/db09-9028. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 2.Tabak AG, Jokela M, Akbaraly TN, Brunner EJ, Kivimaki M, Witte DR. Trajectories of glycaemia, insulin sensitivity, and insulin secretion before diagnosis of type 2 diabetes: an analysis from the Whitehall II study. Lancet. 2009;373(9682):2215–21. doi: 10.1016/S0140-6736(09)60619-X. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 3.Martin BC, Warram JH, Rosner B, Rich SS, Soeldner JS, Krolewski AS. Familial clustering of insulin sensitivity. Diabetes. 1992;41:850–4. doi: 10.2337/diab.41.7.850. [DOI] [PubMed] [Google Scholar]
- 4.IDF. IDF Diabetes Atlas. Seventh. Brussels, Belgium: International Diabetes Federation; 2015. p. 144. [Google Scholar]
- 5.Pauling L, Robinson AB, Teranishi R, Cary P. Quantitative analysis of urine vapor and breath by gas-liquid partition chromatography. Proc Natl Acad Sci U S A. 1971;68(10):2374–6. doi: 10.1073/pnas.68.10.2374. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 6.Rhee EP, Gerszten RE. Metabolomics and cardiovascular biomarker discovery. Clin Chem. 2012;58(1):139–47. doi: 10.1373/clinchem.2011.169573. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 7.Bain JR, Stevens RD, Wenner BR, Ilkayeva O, Muoio DM, Newgard CB. Metabolomics applied to diabetes research. Moving from information to knowledge. Diabetes. 2009;58:2429–43. doi: 10.2337/db09-0580. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 8.Patti GJ, Yanes O, Siuzdak G. Innovation: Metabolomics: the apogee of the omics trilogy. Nat Rev Mol Cell Biol. 2012;13(4):263–9. doi: 10.1038/nrm3314. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 9.Roberts LD, Gerszten RE. Toward new biomarkers of cardiometabolic diseases. Cell Metab. 2013;18(1):43–50. doi: 10.1016/j.cmet.2013.05.009. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 10.Drogan D, Dunn WB, Lin W, Buijsse B, Schulze MB, Langenberg C, et al. Untargeted metabolic profiling identifies altered serum metabolites of type 2 diabetes mellitus in a prospective, nested case control study. Clin Chem. 2015;61(3):487–97. doi: 10.1373/clinchem.2014.228965. [DOI] [PubMed] [Google Scholar]
- 11.Park S, Sadanala KC, Kim EK. A Metabolomic Approach to Understanding the Metabolic Link between Obesity and Diabetes. Mol Cells. 2015;38(7):587–96. doi: 10.14348/molcells.2015.0126. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 12.Fiehn O, Garvey WT, Newman JW, Lok KH, Hoppel CL, Adams SH. Plasma metabolomic profiles reflective of glucose homeostasis in non-diabetic and type 2 diabetic obese African-American women. PLoS One. 2010;5(12):e15234. doi: 10.1371/journal.pone.0015234. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 13.Gogna N, Krishna M, Oommen AM, Dorai K. Investigating correlations in the altered metabolic profiles of obese and diabetic subjects in a South Indian Asian population using an NMR-based metabolomic approach. Mol Biosyst. 2015;11(2):595–606. doi: 10.1039/c4mb00507d. [DOI] [PubMed] [Google Scholar]
- 14.Xu F, Tavintharan S, Sum CF, Woon K, Lim SC, Ong CN. Metabolic signature shift in type 2 diabetes mellitus revealed by mass spectrometry-based metabolomics. JClinEndocrinolMetab. 2013;98(6):E1060–E5. doi: 10.1210/jc.2012-4132. [DOI] [PubMed] [Google Scholar]
- 15.Suhre K, Meisinger C, Doring A, Altmaier E, Belcredi P, Gieger C, et al. Metabolic footprint of diabetes: a multiplatform metabolomics study in an epidemiological setting. PLoS One. 2010;5(11):e13953. doi: 10.1371/journal.pone.0013953. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 16.Menni C, Fauman E, Erte I, Perry JR, Kastenmuller G, Shin SY, et al. Biomarkers for type 2 diabetes and impaired fasting glucose using a nontargeted metabolomics approach. Diabetes. 2013;62(12):4270–6. doi: 10.2337/db13-0570. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 17.Shaham O, Wei R, Wang TJ, Ricciardi C, Lewis GD, Vasan RS, et al. Metabolic profiling of the human response to a glucose challenge reveals distinct axes of insulin sensitivity. Mol Syst Biol. 2008;4:214. doi: 10.1038/msb.2008.50. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 18.Spegel P, Malmgren S, Sharoyko VV, Newsholme P, Koeck T, Mulder H. Metabolomic analyses reveal profound differences in glycolytic and tricarboxylic acid cycle metabolism in glucose-responsive and -unresponsive clonal beta-cell lines. Biochem J. 2011;435(1):277–84. doi: 10.1042/BJ20100655. [DOI] [PubMed] [Google Scholar]
- 19.Boucher A, Lu D, Burgess SC, Telemaque-Potts S, Jensen MV, Mulder H, et al. Biochemical mechanism of lipid-induced impairment of glucose-stimulated insulin secretion and reversal with a malate analogue. J Biol Chem. 2004;279(26):27263–71. doi: 10.1074/jbc.M401167200. [DOI] [PubMed] [Google Scholar]
- 20.Lu D, Mulder H, Zhao P, Burgess SC, Jensen MV, Kamzolova S, et al. 13C NMR isotopomer analysis reveals a connection between pyruvate cycling and glucose-stimulated insulin secretion (GSIS) Proc Natl Acad Sci U S A. 2002;99(5):2708–13. doi: 10.1073/pnas.052005699. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 21.Kelley DE, Mokan M, Simoneau JA, Mandarino LJ. Interaction between glucose and free fatty acid metabolism in human skeletal muscle. JClinInvest. 1993;92(1):91–8. doi: 10.1172/JCI116603. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 22.Koves TR, Li P, An J, Akimoto T, Slentz D, Ilkayeva O, et al. Peroxisome proliferator-activated receptor-gamma co-activator 1alpha-mediated metabolic remodeling of skeletal myocytes mimics exercise training and reverses lipid-induced mitochondrial inefficiency. J Biol Chem. 2005;280(39):33588–98. doi: 10.1074/jbc.M507621200. [DOI] [PubMed] [Google Scholar]
- 23.Koves TR, Ussher JR, Noland RC, Slentz D, Mosedale M, Ilkayeva O, et al. Mitochondrial overload and incomplete Fatty Acid oxidation contribute to skeletal muscle insulin resistance. Cell Metab. 2008;7(1):45–56. doi: 10.1016/j.cmet.2007.10.013. [DOI] [PubMed] [Google Scholar]
- 24.Giesbertz P, Padberg I, Rein D, Ecker J, Hofle AS, Spanier B, et al. Metabolite profiling in plasma and tissues of ob/ob and db/db mice identifies novel markers of obesity and type 2 diabetes. Diabetologia. 2015;58(9):2133–43. doi: 10.1007/s00125-015-3656-y. [DOI] [PubMed] [Google Scholar]
- 25.Kowalski GM, De Souza DP, Burch ML, Hamley S, Kloehn J, Selathurai A, et al. Application of dynamic metabolomics to examine in vivo skeletal muscle glucose metabolism in the chronically high-fat fed mouse. Biochem Biophys Res Commun. 2015;462(1):27–32. doi: 10.1016/j.bbrc.2015.04.096. [DOI] [PubMed] [Google Scholar]
- 26.Newgard CB, An J, Bain JR, Muehlbauer MJ, Stevens RD, Lien LF, et al. A branched-chain amino acid-related metabolic signature that differentiates obese and lean humans and contributes to insulin resistance. Cell Metab. 2009;9(4):311–26. doi: 10.1016/j.cmet.2009.02.002. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 27.Salek RM, Maguire ML, Bentley E, Rubtsov DV, Hough T, Cheeseman M, et al. A metabolomic comparison of urinary changes in type 2 diabetes in mouse, rat, and human. PhysiolGenomics. 2007;29(2):99–108. doi: 10.1152/physiolgenomics.00194.2006. [DOI] [PubMed] [Google Scholar]
- 28.Connor SC, Hansen MK, Corner A, Smith RF, Ryan TE. Integration of metabolomics and transcriptomics data to aid biomarker discovery in type 2 diabetes. Mol Biosyst. 2010;6(5):909–21. doi: 10.1039/b914182k. [DOI] [PubMed] [Google Scholar]
- 29.Adams SH, Hoppel CL, Lok KH, Zhao L, Wong SW, Minkler PE, et al. Plasma acylcarnitine profiles suggest incomplete long-chain fatty acid beta-oxidation and altered tricarboxylic acid cycle activity in type 2 diabetic African-American women. J Nutr. 2009;139(6):1073–81. doi: 10.3945/jn.108.103754. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 30.Huffman KM, Shah SH, Stevens RD, Bain JR, Muehlbauer M, Slentz CA, et al. Relationships between circulating metabolic intermediates and insulin action in overweight to obese, inactive men and women. Diabetes Care. 2009;32(9):1678–83. doi: 10.2337/dc08-2075. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 31.Felig P, Marliss E, Cahill GF., Jr Plasma amino acid levels and insulin secretion in obesity. N Engl J Med. 1969;281(15):811–6. doi: 10.1056/NEJM196910092811503. [DOI] [PubMed] [Google Scholar]
- 32.Wurtz P, Soininen P, Kangas AJ, Ronnemaa T, Lehtimaki T, Kahonen M, et al. Branched-chain and aromatic amino acids are predictors of insulin resistance in young adults. Diabetes Care. 2013;36(3):648–55. doi: 10.2337/dc12-0895. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 33.Wang TJ, Larson MG, Vasan RS, Cheng S, Rhee EP, McCabe E, et al. Metabolite profiles and the risk of developing diabetes. NatMed. 2011;17(4):448–53. doi: 10.1038/nm.2307. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 34.Yamakado M, Tanaka T, Nagao K, Ishizaka Y, Mitushima T, Tani M, et al. Plasma amino acid profile is associated with visceral fat accumulation in obese Japanese subjects. Clin Obes. 2012;2(1–2):29–40. doi: 10.1111/j.1758-8111.2012.00039.x. [DOI] [PubMed] [Google Scholar]
- 35.Gall WE, Beebe K, Lawton KA, Adam KP, Mitchell MW, Nakhle PJ, et al. alpha-hydroxybutyrate is an early biomarker of insulin resistance and glucose intolerance in a nondiabetic population. PLoSONE. 2010;5(5):e10883. doi: 10.1371/journal.pone.0010883. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 36.Cheng S, Rhee EP, Larson MG, Lewis GD, McCabe EL, Shen D, et al. Metabolite profiling identifies pathways associated with metabolic risk in humans. Circulation. 2012;125(18):2222–31. doi: 10.1161/CIRCULATIONAHA.111.067827. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 37.Wurtz P, Tiainen M, Makinen VP, Kangas AJ, Soininen P, Saltevo J, et al. Circulating metabolite predictors of glycemia in middle-aged men and women. Diabetes Care. 2012;35(8):1749–56. doi: 10.2337/dc11-1838. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 38.Stancakova A, Civelek M, Saleem NK, Soininen P, Kangas AJ, Cederberg H, et al. Hyperglycemia and a common variant of GCKR are associated with the levels of eight amino acids in 9,369 Finnish men. Diabetes. 2012;61(7):1895–902. doi: 10.2337/db11-1378. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 39.Wang-Sattler R, Yu Z, Herder C, Messias AC, Floegel A, He Y, et al. Novel biomarkers for pre-diabetes identified by metabolomics. MolSystBiol. 2012;8:615. doi: 10.1038/msb.2012.43. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 40.Sekhar RV, McKay SV, Patel SG, Guthikonda AP, Reddy VT, Balasubramanyam A, et al. Glutathione synthesis is diminished in patients with uncontrolled diabetes and restored by dietary supplementation with cysteine and glycine. Diabetes Care. 2011;34(1):162–7. doi: 10.2337/dc10-1006. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 41.Ejaz A, Martinez-Guino L, Goldfine AB, Ribas-Aulinas F, De Nigris V, Ribo S, et al. Dietary Betaine Supplementation Increases Fgf21 Levels to Improve Glucose Homeostasis and Reduce Hepatic Lipid Accumulation in Mice. Diabetes. 2016 doi: 10.2337/db15-1094. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 42.Walford GA, Davis J, Warner AS, Ackerman RJ, Billings LK, Chamarthi B, et al. Branched chain and aromatic amino acids change acutely following two medical therapies for type 2 diabetes mellitus. Metabolism. 2013 doi: 10.1016/j.metabol.2013.07.003. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 43.Walford GA, Ma Y, Clish C, Florez JC, Wang TJ, Gerszten RE, et al. Metabolite profiles of diabetes incidence and intervention response in the Diabetes Prevention Program. Diabetes. 2016 doi: 10.2337/db15-1063. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 44.Laferrere B, Reilly D, Arias S, Swerdlow N, Gorroochurn P, Bawa B, et al. Differential metabolic impact of gastric bypass surgery versus dietary intervention in obese diabetic subjects despite identical weight loss. SciTranslMed. 2011;3(80):80re2. doi: 10.1126/scitranslmed.3002043. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 45.Lyssenko V, Jonsson A, Almgren P, Pulizzi N, Isomaa B, Tuomi T, et al. Clinical risk factors, DNA variants, and the development of type 2 diabetes. N Engl J Med. 2008;359(21):2220–32. doi: 10.1056/NEJMoa0801869. [DOI] [PubMed] [Google Scholar]
- 46.Floyd JC, Jr, Fajans SS, Conn JW, Knopf RF, Rull J. Stimulation of insulin secretion by amino acids. J Clin Invest. 1966;45(9):1487–502. doi: 10.1172/JCI105456. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 47.van Loon LJ, Saris WH, Verhagen H, Wagenmakers AJ. Plasma insulin responses after ingestion of different amino acid or protein mixtures with carbohydrate. Am J Clin Nutr. 2000;72(1):96–105. doi: 10.1093/ajcn/72.1.96. [DOI] [PubMed] [Google Scholar]
- 48.Newgard CB, Matschinsky FM. Substrate control of insulin resistance. In: Jefferson L, Cherrington A, editors. Handbook in Physiology. Vol. 2. Oxford University Press; 2001. pp. 125–51. [Google Scholar]
- 49.Nilsson M, Holst JJ, Bjorck IM. Metabolic effects of amino acid mixtures and whey protein in healthy subjects: studies using glucose-equivalent drinks. Am J Clin Nutr. 2007;85(4):996–1004. doi: 10.1093/ajcn/85.4.996. [DOI] [PubMed] [Google Scholar]
- 50.Patti ME, Brambilla E, Luzi L, Landaker EJ, Kahn CR. Bidirectional modulation of insulin action by amino acids. JClinInvest. 1998;101:1519–29. doi: 10.1172/JCI1326. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 51.Lynch CJ, Adams SH. Branched-chain amino acids in metabolic signalling and insulin resistance. Nat Rev Endocrinol. 2014;10(12):723–36. doi: 10.1038/nrendo.2014.171. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 52.Macotela Y, Emanuelli B, Bang AM, Espinoza DO, Boucher J, Beebe K, et al. Dietary leucine–an environmental modifier of insulin resistance acting on multiple levels of metabolism. PLoSONE. 2011;6(6):e21187. doi: 10.1371/journal.pone.0021187. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 53.She P, Reid TM, Bronson SK, Vary TC, Hajnal A, Lynch CJ, et al. Disruption of BCATm in mice leads to increased energy expenditure associated with the activation of a futile protein turnover cycle. Cell Metab. 2007;6(3):181–94. doi: 10.1016/j.cmet.2007.08.003. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 54.Muoio DM, Newgard CB. Mechanisms of disease:Molecular and metabolic mechanisms of insulin resistance and beta-cell failure in type 2 diabetes. Nat Rev Mol Cell Biol. 2008;9(3):193–205. doi: 10.1038/nrm2327. [DOI] [PubMed] [Google Scholar]
- 55.Morino K, Petersen KF, Shulman GI. Molecular mechanisms of insulin resistance in humans and their potential links with mitochondrial dysfunction. Diabetes. 2006;55(Suppl 2):S9–S15. doi: 10.2337/db06-S002. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 56.Boden G, Shulman GI. Free fatty acids in obesity and type 2 diabetes: defining their role in the development of insulin resistance and beta-cell dysfunction. EurJClinInvest. 2002;32(Suppl 3):14–23. doi: 10.1046/j.1365-2362.32.s3.3.x. [DOI] [PubMed] [Google Scholar]
- 57.Lowell BB, Shulman GI. Mitochondrial dysfunction and type 2 diabetes. Science. 2005;307(5708):384–7. doi: 10.1126/science.1104343. [DOI] [PubMed] [Google Scholar]
- 58.Samuel VT, Petersen KF, Shulman GI. Lipid-induced insulin resistance: unravelling the mechanism. Lancet. 2010;375(9733):2267–77. doi: 10.1016/S0140-6736(10)60408-4. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 59.Dube JJ, Amati F, Toledo FG, Stefanovic-Racic M, Rossi A, Coen P, et al. Effects of weight loss and exercise on insulin resistance, and intramyocellular triacylglycerol, diacylglycerol and ceramide. Diabetologia. 2011;54(5):1147–56. doi: 10.1007/s00125-011-2065-0. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 60.Itani SI, Ruderman NB, Schmieder F, Boden G. Lipid-induced insulin resistance in human muscle is associated with changes in diacylglycerol, protein kinase C, and IkappaB-alpha. Diabetes. 2002;51(7):2005–11. doi: 10.2337/diabetes.51.7.2005. [DOI] [PubMed] [Google Scholar]
- 61.Adams JM, Pratipanawatr T, Berria R, Wang E, DeFronzo RA, Sullards MC, et al. Ceramide content is increased in skeletal muscle from obese insulin-resistant humans. Diabetes. 2004;53(1):25–31. doi: 10.2337/diabetes.53.1.25. [DOI] [PubMed] [Google Scholar]
- 62.Oh DY, Talukdar S, Bae EJ, Imamura T, Morinaga H, Fan W, et al. GPR120 is an omega-3 fatty acid receptor mediating potent anti-inflammatory and insulin-sensitizing effects. Cell. 2010;142(5):687–98. doi: 10.1016/j.cell.2010.07.041. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 63.Virtanen JK, Mursu J, Voutilainen S, Uusitupa M, Tuomainen TP. Serum omega-3 polyunsaturated fatty acids and risk of incident type 2 diabetes in men: the Kuopio Ischemic Heart Disease Risk Factor study. Diabetes Care. 2014;37(1):189–96. doi: 10.2337/dc13-1504. [DOI] [PubMed] [Google Scholar]
- 64.Cao H, Gerhold K, Mayers JR, Wiest MM, Watkins SM, Hotamisligil GS. Identification of a lipokine, a lipid hormone linking adipose tissue to systemic metabolism. Cell. 2008;134(6):933–44. doi: 10.1016/j.cell.2008.07.048. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 65.Rhee EP, Cheng S, Larson MG, Walford GA, Lewis GD, McCabe E, et al. Lipid profiling identifies a triacylglycerol signature of insulin resistance and improves diabetes prediction in humans. JClinInvest. 2011;121(4):1402–11. doi: 10.1172/JCI44442. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 66.Riserus U, Willett WC, Hu FB. Dietary fats and prevention of type 2 diabetes. Prog Lipid Res. 2009;48(1):44–51. doi: 10.1016/j.plipres.2008.10.002. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 67.Yore MM, Syed I, Moraes-Vieira PM, Zhang T, Herman MA, Homan EA, et al. Discovery of a class of endogenous mammalian lipids with anti-diabetic and anti-inflammatory effects. Cell. 2014;159(2):318–32. doi: 10.1016/j.cell.2014.09.035. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 68.Illig T, Gieger C, Zhai G, Romisch-Margl W, Wang-Sattler R, Prehn C, et al. A genome-wide perspective of genetic variation in human metabolism. NatGenet. 2010;42(2):137–41. doi: 10.1038/ng.507. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 69.Xie W, Wood AR, Lyssenko V, Weedon MN, Knowles JW, Alkayyali S, et al. Genetic variants associated with glycine metabolism and their role in insulin sensitivity and type 2 diabetes. Diabetes. 2013;62(6):2141–50. doi: 10.2337/db12-0876. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 70.Tannock GW. The normal microflora: new concepts in health promotion. Microbiol Sci. 1988;5(1):4–8. [PubMed] [Google Scholar]
- 71.Menni C, Zhai G, MacGregor A, Prehn C, Romisch-Margl W, Suhre K, et al. Targeted metabolomics profiles are strongly correlated with nutritional patterns in women. Metabolomics. 2013;9(2):506–14. doi: 10.1007/s11306-012-0469-6. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 72.Roberts LD, Bostrom P, O’Sullivan JF, Schinzel RT, Lewis GD, Dejam A, et al. beta-Aminoisobutyric acid induces browning of white fat and hepatic beta-oxidation and is inversely correlated with cardiometabolic risk factors. Cell Metab. 2014;19(1):96–108. doi: 10.1016/j.cmet.2013.12.003. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 73.Lawton KA, Berger A, Mitchell M, Milgram KE, Evans AM, Guo L, et al. Analysis of the adult human plasma metabolome. Pharmacogenomics. 2008;9(4):383–97. doi: 10.2217/14622416.9.4.383. [DOI] [PubMed] [Google Scholar]
- 74.Nicholson JK, Wilson ID. Opinion: understanding ‘global’ systems biology: metabonomics and the continuum of metabolism. Nat Rev Drug Discov. 2003;2(8):668–76. doi: 10.1038/nrd1157. [DOI] [PubMed] [Google Scholar]
- 75.Nicholson JK, Holmes E, Lindon JC, Wilson ID. The challenges of modeling mammalian biocomplexity. Nat Biotechnol. 2004;22(10):1268–74. doi: 10.1038/nbt1015. [DOI] [PubMed] [Google Scholar]
- 76.Wu H, Tremaroli V, Backhed F. Linking Microbiota to Human Diseases: A Systems Biology Perspective. Trends Endocrinol Metab. 2015;26(12):758–70. doi: 10.1016/j.tem.2015.09.011. [DOI] [PubMed] [Google Scholar]
- 77.Turnbaugh PJ, Hamady M, Yatsunenko T, Cantarel BL, Duncan A, Ley RE, et al. A core gut microbiome in obese and lean twins. Nature. 2009;457(7228):480–4. doi: 10.1038/nature07540. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 78.Ley RE, Backhed F, Turnbaugh P, Lozupone CA, Knight RD, Gordon JI. Obesity alters gut microbial ecology. Proc Natl Acad Sci U S A. 2005;102(31):11070–5. doi: 10.1073/pnas.0504978102. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 79.Remely M, Dworzak S, Hippe B, Zwielehner J, Aumüller E, Brath H, et al. Abundance and Diversity of Microbiota in Type 2 Diabetes and Obesity. Diabetes and Metabolism. 2013;4(3) [Google Scholar]
- 80.Larsen N, Vogensen FK, van den Berg FW, Nielsen DS, Andreasen AS, Pedersen BK, et al. Gut microbiota in human adults with type 2 diabetes differs from non-diabetic adults. PLoS One. 2010;5(2):e9085. doi: 10.1371/journal.pone.0009085. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 81.Wu X, Ma C, Han L, Nawaz M, Gao F, Zhang X, et al. Molecular characterisation of the faecal microbiota in patients with type II diabetes. Curr Microbiol. 2010;61(1):69–78. doi: 10.1007/s00284-010-9582-9. [DOI] [PubMed] [Google Scholar]
- 82.Zhang H, DiBaise JK, Zuccolo A, Kundrna D, Braidotti M, Yu Y, et al. Human gut microbiota in obesity and after gastric bypass. PNAS. 2008:1–6. doi: 10.1073/pnas.0812600106. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 83.Holmes E, Li JV, Athanasiou T, Ashrafian H, Nicholson JK. Understanding the role of gut microbiome-host metabolic signal disruption in health and disease. Trends Microbiol. 2011;19(7):349–59. doi: 10.1016/j.tim.2011.05.006. [DOI] [PubMed] [Google Scholar]
- 84.Patti ME, Houten SM, Bianco AC, Bernier R, Larsen PR, Holst JJ, et al. Serum Bile Acids Are Higher in Humans With Prior Gastric Bypass: Potential Contribution to Improved Glucose and Lipid Metabolism. Obesity (SilverSpring) 2009 doi: 10.1038/oby.2009.102. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 85.Mutch DM, Fuhrmann JC, Rein D, Wiemer JC, Bouillot JL, Poitou C, et al. Metabolite profiling identifies candidate markers reflecting the clinical adaptations associated with Roux-en-Y gastric bypass surgery. PLoSONE. 2009;4(11):e7905. doi: 10.1371/journal.pone.0007905. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 86.Dumas ME, Barton RH, Toye A, Cloarec O, Blancher C, Rothwell A, et al. Metabolic profiling reveals a contribution of gut microbiota to fatty liver phenotype in insulin-resistant mice. Proc NatlAcad Sci U SA. 2006;103(33):12511–6. doi: 10.1073/pnas.0601056103. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 87.Turnbaugh PJ, Ley RE, Mahowald MA, Magrini V, Mardis ER, Gordon JI. An obesity-associated gut microbiome with increased capacity for energy harvest. Nature. 2006;444(7122):1027–31. doi: 10.1038/nature05414. [DOI] [PubMed] [Google Scholar]
- 88.Turnbaugh PJ, Backhed F, Fulton L, Gordon JI. Diet-induced obesity is linked to marked but reversible alterations in the mouse distal gut microbiome. Cell Host Microbe. 2008;3(4):213–23. doi: 10.1016/j.chom.2008.02.015. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 89.Tremaroli V, Karlsson F, Werling M, Stahlman M, Kovatcheva-Datchary P, Olbers T, et al. Roux-en-Y Gastric Bypass and Vertical Banded Gastroplasty Induce Long-Term Changes on the Human Gut Microbiome Contributing to Fat Mass Regulation. Cell Metab. 2015;22(2):228–38. doi: 10.1016/j.cmet.2015.07.009. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 90.Vrieze A, Van Nood E, Holleman F, Salojarvi J, Kootte RS, Bartelsman JF, et al. Transfer of intestinal microbiota from lean donors increases insulin sensitivity in individuals with metabolic syndrome. Gastroenterology. 2012;143(4):913–6 e7. doi: 10.1053/j.gastro.2012.06.031. [DOI] [PubMed] [Google Scholar]
- 91.Wang Z, Klipfell E, Bennett BJ, Koeth R, Levison BS, Dugar B, et al. Gut flora metabolism of phosphatidylcholine promotes cardiovascular disease. Nature. 2011;472(7341):57–63. doi: 10.1038/nature09922. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 92.Tang WH, Wang Z, Levison BS, Koeth RA, Britt EB, Fu X, et al. Intestinal microbial metabolism of phosphatidylcholine and cardiovascular risk. N Engl J Med. 2013;368(17):1575–84. doi: 10.1056/NEJMoa1109400. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 93.Koeth RA, Wang Z, Levison BS, Buffa JA, Org E, Sheehy BT, et al. Intestinal microbiota metabolism of L-carnitine, a nutrient in red meat, promotes atherosclerosis. Nat Med. 2013;19(5):576–85. doi: 10.1038/nm.3145. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 94.Organ CL, Otsuka H, Bhushan S, Wang Z, Bradley J, Trivedi R, et al. Choline Diet and Its Gut Microbe-Derived Metabolite, Trimethylamine N-Oxide, Exacerbate Pressure Overload-Induced Heart Failure. Circ Heart Fail. 2016;9(1):e002314. doi: 10.1161/CIRCHEARTFAILURE.115.002314. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 95.Sas KM, Karnovsky A, Michailidis G, Pennathur S. Metabolomics and diabetes: analytical and computational approaches. Diabetes. 2015;64(3):718–32. doi: 10.2337/db14-0509. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 96.Ferrara CT, Wang P, Neto EC, Stevens RD, Bain JR, Wenner BR, et al. Genetic networks of liver metabolism revealed by integration of metabolic and transcriptional profiling. PLoSGenet. 2008;4(3):e1000034. doi: 10.1371/journal.pgen.1000034. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 97.Jerby L, Shlomi T, Ruppin E. Computational reconstruction of tissue-specific metabolic models: application to human liver metabolism. MolSystBiol. 2010;6:401. doi: 10.1038/msb.2010.56. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 98.Gille C, Bolling C, Hoppe A, Bulik S, Hoffmann S, Hubner K, et al. HepatoNet1: a comprehensive metabolic reconstruction of the human hepatocyte for the analysis of liver physiology. Mol Syst Biol. 2010;6:411. doi: 10.1038/msb.2010.62. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 99.Bordbar A, Feist AM, Usaite-Black R, Woodcock J, Palsson BO, Famili I. A multi-tissue type genome-scale metabolic network for analysis of whole-body systems physiology. BMC Syst Biol. 2011;5:180. doi: 10.1186/1752-0509-5-180. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 100.Nogiec C, Burkart A, Dreyfuss JM, Lerin C, Kasif S, Patti ME. Metabolic modeling of muscle metabolism identifies key reactions linked to insulin resistance phenotypes. Mol Metab. 2015;4(3):151–63. doi: 10.1016/j.molmet.2014.12.012. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 101.Shlomi T, Cabili MN, Ruppin E. Predicting metabolic biomarkers of human inborn errors of metabolism. Mol SystBiol. 2009;5:263. doi: 10.1038/msb.2009.22. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 102.Kuc S, Koster MP, Pennings JL, Hankemeier T, Berger R, Harms AC, et al. Metabolomics profiling for identification of novel potential markers in early prediction of preeclampsia. PLoS One. 2014;9(5):e98540. doi: 10.1371/journal.pone.0098540. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 103.Maitre L, Fthenou E, Athersuch T, Coen M, Toledano MB, Holmes E, et al. Urinary metabolic profiles in early pregnancy are associated with preterm birth and fetal growth restriction in the Rhea mother-child cohort study. BMC Med. 2014;12:110. doi: 10.1186/1741-7015-12-110. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 104.de Seymour JV, Conlon CA, Sulek K, Villas Boas SG, McCowan LM, Kenny LC, et al. Early pregnancy metabolite profiling discovers a potential biomarker for the subsequent development of gestational diabetes mellitus. Acta Diabetol. 2014;51(5):887–90. doi: 10.1007/s00592-014-0626-7. [DOI] [PubMed] [Google Scholar]
- 105.Ciborowski M, Zbucka-Kretowska M, Bomba-Opon D, Wielgos M, Brawura-Biskupski-Samaha R, Pierzynski P, et al. Potential first trimester metabolomic biomarkers of abnormal birth weight in healthy pregnancies. Prenat Diagn. 2014;34(9):870–7. doi: 10.1002/pd.4386. [DOI] [PubMed] [Google Scholar]
- 106.Heazell AE, Bernatavicius G, Warrander L, Brown MC, Dunn WB. A metabolomic approach identifies differences in maternal serum in third trimester pregnancies that end in poor perinatal outcome. Reprod Sci. 2012;19(8):863–75. doi: 10.1177/1933719112438446. [DOI] [PubMed] [Google Scholar]
- 107.Bahado-Singh RO, Akolekar R, Mandal R, Dong E, Xia J, Kruger M, et al. First-trimester metabolomic detection of late-onset preeclampsia. Am J Obstet Gynecol. 2013;208(1):58 e1–7. doi: 10.1016/j.ajog.2012.11.003. [DOI] [PubMed] [Google Scholar]
- 108.Horgan RP, Broadhurst DI, Walsh SK, Dunn WB, Brown M, Roberts CT, et al. Metabolic profiling uncovers a phenotypic signature of small for gestational age in early pregnancy. J Proteome Res. 2011;10(8):3660–73. doi: 10.1021/pr2002897. [DOI] [PubMed] [Google Scholar]
- 109.Huynh J, Xiong G, Bentley-Lewis R. A systematic review of metabolite profiling in gestational diabetes mellitus. Diabetologia. 2014;57(12):2453–64. doi: 10.1007/s00125-014-3371-0. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 110.Metzger BE, Phelps RL, Freinkel N, Navickas IA. Effects of gestational diabetes on diurnal profiles of plasma glucose, lipids, and individual amino acids. Diabetes Care. 1980;3(3):402–9. doi: 10.2337/diacare.3.3.402. [DOI] [PubMed] [Google Scholar]
- 111.Scholtens DM, Muehlbauer MJ, Daya NR, Stevens RD, Dyer AR, Lowe LP, et al. Metabolomics reveals broad-scale metabolic perturbations in hyperglycemic mothers during pregnancy. Diabetes Care. 2014;37(1):158–66. doi: 10.2337/dc13-0989. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 112.Pappa KI, Vlachos G, Theodora M, Roubelaki M, Angelidou K, Antsaklis A. Intermediate metabolism in association with the amino acid profile during the third trimester of normal pregnancy and diet-controlled gestational diabetes. Am J Obstet Gynecol. 2007;196(1):65 e1, 5. doi: 10.1016/j.ajog.2006.06.094. [DOI] [PubMed] [Google Scholar]
- 113.Diaz SO, Pinto J, Graca G, Duarte IF, Barros AS, Galhano E, et al. Metabolic biomarkers of prenatal disorders: an exploratory NMR metabonomics study of second trimester maternal urine and blood plasma. J Proteome Res. 2011;10(8):3732–42. doi: 10.1021/pr200352m. [DOI] [PubMed] [Google Scholar]
- 114.Dudzik D, Zorawski M, Skotnicki M, Zarzycki W, Kozlowska G, Bibik-Malinowska K, et al. Metabolic fingerprint of Gestational Diabetes Mellitus. Journal of proteomics. 2014;103:57–71. doi: 10.1016/j.jprot.2014.03.025. [DOI] [PubMed] [Google Scholar]
- 115.Pinto J, Almeida LM, Martins AS, Duarte D, Barros AS, Galhano E, et al. Prediction of Gestational Diabetes through NMR Metabolomics of Maternal Blood. J Proteome Res. 2015;14(6):2696–706. doi: 10.1021/acs.jproteome.5b00260. [DOI] [PubMed] [Google Scholar]
- 116.Cetin I, de Santis MS, Taricco E, Radaelli T, Teng C, Ronzoni S, et al. Maternal and fetal amino acid concentrations in normal pregnancies and in pregnancies with gestational diabetes mellitus. Am J Obstet Gynecol. 2005;192(2):610–7. doi: 10.1016/j.ajog.2004.08.011. [DOI] [PubMed] [Google Scholar]
- 117.Lehmann R, Friedrich T, Krebiehl G, Sonntag D, Haring HU, Fritsche A, et al. Metabolic profiles during an oral glucose tolerance test in pregnant women with and without gestational diabetes. Exp Clin Endocrinol Diabetes. 2015;123(7):483–38. doi: 10.1055/s-0035-1549887. [DOI] [PubMed] [Google Scholar]
- 118.Prentice KJ, Luu L, Allister EM, Liu Y, Jun LS, Sloop KW, et al. The Furan Fatty Acid Metabolite CMPF Is Elevated in Diabetes and Induces beta Cell Dysfunction. Cell Metab. 2014;19(4):653–66. doi: 10.1016/j.cmet.2014.03.008. [DOI] [PubMed] [Google Scholar]
- 119.Dani C, Bresci C, Berti E, Ottanelli S, Mello G, Mecacci F, et al. Metabolomic profile of term infants of gestational diabetic mothers. J Matern Fetal Neonatal Med. 2014;27(6):537–42. doi: 10.3109/14767058.2013.823941. [DOI] [PubMed] [Google Scholar]
- 120.Gluckman PD, Hanson MA, Cooper C, Thornburg KL. Effect of in utero and early-life conditions on adult health and disease. N Engl J Med. 2008;359(1):61–73. doi: 10.1056/NEJMra0708473. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 121.Wang G, Divall S, Radovick S, Paige D, Ning Y, Chen Z, et al. Preterm birth and random plasma insulin levels at birth and in early childhood. JAMA. 2014;311(6):587–96. doi: 10.1001/jama.2014.1. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 122.Dessi A, Atzori L, Noto A, Visser GH, Gazzolo D, Zanardo V, et al. Metabolomics in newborns with intrauterine growth retardation (IUGR): urine reveals markers of metabolic syndrome. J Matern Fetal Neonatal Med. 2011;24(Suppl 2):35–9. doi: 10.3109/14767058.2011.605868. [DOI] [PubMed] [Google Scholar]
- 123.Ivorra C, Garcia-Vicent C, Chaves FJ, Monleon D, Morales JM, Lurbe E. Metabolomic profiling in blood from umbilical cords of low birth weight newborns. J Transl Med. 2012;10:142. doi: 10.1186/1479-5876-10-142. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 124.Alexandre-Gouabau MC, Courant F, Moyon T, Kuster A, Le GG, Tea I, et al. Maternal and cord blood LC-HRMS metabolomics reveal alterations in energy and polyamine metabolism, and oxidative stress in very-low birth weight infants. J ProteomeRes. 2013;12(6):2764–78. doi: 10.1021/pr400122v. [DOI] [PubMed] [Google Scholar]
- 125.Leunissen RW, Kerkhof GF, Stijnen T, Hokken-Koelega A. Timing and tempo of first-year rapid growth in relation to cardiovascular and metabolic risk profile in early adulthood. JAMA. 2009;301(21):2234–42. doi: 10.1001/jama.2009.761. [DOI] [PubMed] [Google Scholar]
- 126.Barker DJ, Osmond C, Forsen TJ, Kajantie E, Eriksson JG. Trajectories of growth among children who have coronary events as adults. New England Journal of Medicine. 2005;353(17):1802–9. doi: 10.1056/NEJMoa044160. [DOI] [PubMed] [Google Scholar]
- 127.Isganaitis E, Rifas-Shiman SL, Oken E, Dreyfuss JM, Gall W, Gillman MW, et al. Associations of cord blood metabolites with early childhood obesity risk. Int J Obes (Lond) 2015;39(7):1041–8. doi: 10.1038/ijo.2015.39. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 128.Rzehak P, Hellmuth C, Uhl O, Kirchberg FF, Peissner W, Harder U, et al. Rapid growth and childhood obesity are strongly associated with lysoPC(14:0) Ann Nutr Metab. 2014;64(3–4):294–303. doi: 10.1159/000365037. [DOI] [PubMed] [Google Scholar]
- 129.Stevens A, Bonshek C, Whatmore A, Butcher I, Hanson D, De Leonibus C, et al. Insights into the pathophysiology of catch-up compared with non-catch-up growth in children born small for gestational age: an integrated analysis of metabolic and transcriptomic data. Pharmacogenomics J. 2014;14(4):376–84. doi: 10.1038/tpj.2014.4. [DOI] [PubMed] [Google Scholar]
- 130.Perng W, Gillman MW, Fleisch AF, Michalek RD, Watkins SM, Isganaitis E, et al. Metabolomic profiles and childhood obesity. Obesity (Silver Spring) 2014;22(12):2570–8. doi: 10.1002/oby.20901. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 131.Mihalik SJ, Michaliszyn SF, de las HJ, Bacha F, Lee S, Chace DH, et al. Metabolomic profiling of fatty acid and amino acid metabolism in youth with obesity and type 2 diabetes: evidence for enhanced mitochondrial oxidation. Diabetes Care. 2012;35(3):605–11. doi: 10.2337/DC11-1577. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 132.Butte NF, Liu Y, Zakeri IF, Mohney RP, Mehta N, Voruganti VS, et al. Global metabolomic profiling targeting childhood obesity in the Hispanic population. Am J Clin Nutr. 2015;102(2):256–67. doi: 10.3945/ajcn.115.111872. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 133.McCormack SE, Shaham O, McCarthy MA, Deik AA, Wang TJ, Gerszten RE, et al. Circulating branched-chain amino acid concentrations are associated with obesity and future insulin resistance in children and adolescents. Pediatr Obes. 2013;8(1):52–61. doi: 10.1111/j.2047-6310.2012.00087.x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 134.Wahl S, Yu Z, Kleber M, Singmann P, Holzapfel C, He Y, et al. Childhood obesity is associated with changes in the serum metabolite profile. Obes Facts. 2012;5(5):660–70. doi: 10.1159/000343204. [DOI] [PubMed] [Google Scholar]
- 135.Michaliszyn SF, Sjaarda LA, Mihalik SJ, Lee S, Bacha F, Chace DH, et al. Metabolomic profiling of amino acids and beta-cell function relative to insulin sensitivity in youth. J Clin Endocrinol Metab. 2012;97(11):E2119–E24. doi: 10.1210/jc.2012-2170. [DOI] [PMC free article] [PubMed] [Google Scholar]