

Abstract
The adaptation of the brain and cerebral circulation to pregnancy are unique compared with other organs and circulatory systems, ultimately functioning to maintain brain homeostasis. In this review, the effect of pregnancy on critical functions of the cerebral circulation is discussed, including changes occurring at the endothelium and blood-brain barrier, and changes in the structure and function of cerebral arteries and arterioles, hemodynamics, and cerebral blood flow autoregulation.
The brain is an organ of high metabolic demand that consumes ∼20% of the body's oxygen at rest, despite comprising only 2% of body weight (77). Importantly, the brain has a relatively narrow capacity to tolerate changes in ion and water balance, and blood flow (77). The brain is also unique in that it is enclosed in a rigid skull, and therefore increased vascular permeability or volume could result in detrimentally increased intracranial pressure that can cause serious neurological symptoms, brain herniation, and even death. Thus there is a need to maintain tight control of cerebral blood flow (CBF) and water flux in the face of a 40–50% increase in plasma volume and cardiac output during pregnancy and a decline in systemic vascular resistance necessary for the maintenance of a healthy blood pressure (28). In contrast to other organs outside the central nervous system that undergo substantial increases in both perfusion and transvascular filtration during pregnancy, including the uterus, kidney, and heart, the cerebral circulation must resist these adaptations to counterbalance global hemodynamic changes to maintain the delicate microenvironment of the brain. Thus the adaptation of the brain and cerebral circulation to pregnancy appears to be to maintain normalcy despite substantial hormonal and cardiovascular changes in almost every other organ. Throughout this review, the effect of pregnancy on several aspects of the cerebral circulation will be discussed, including the cerebral endothelium and blood-brain barrier (BBB), the structure and function of the cerebrovasculature, hemodynamics, and CBF autoregulation. Furthermore, in the context of these cerebrovascular adaptations, the risk for neurological complications that arises in women with preeclampsia, a dangerous hypertensive complication of pregnancy, will be considered. Preeclampsia affects 3–8% of all pregnancies and is defined as the appearance of hypertension and proteinuria after week 20 of gestation. Many organs are affected by preeclampsia, including the brain, putting women at risk of seizure and hemorrhage. In fact, the cerebral circulation has a central role in neurological complications of preeclampsia, and thus understanding how pregnancy and preeclampsia affect the cerebrovasculature is of interest.
Vasomotor Responses to Circulating Factors
One of the most important adaptations of the cerebral circulation during pregnancy is to counteract the effects of circulating vasoactive factors. During pregnancy, large amounts of hormones are secreted from the placenta, ovaries, and brain into the maternal circulation, including pro- and anti-inflammatory cytokines, chemokines, steroids, and growth factors (1, 78). These factors are critical for the development and survival of the fetus and adaptation of other organ systems needed for a successful pregnancy. Cerebral arteries uniquely adapt during pregnancy to oppose an increase in circulating vasoconstrictors present late in gestation. Exposure of plasma from pregnant women causes vasoconstriction of posterior cerebral arteries from nonpregnant rats; however, this vasoconstrictive effect is absent in arteries from pregnant rats (3). This suggests the cerebral circulation adapts to combat vasoconstrictors present late in gestation. This may be due to either development of resistance to vasoconstrictors circulating during pregnancy or increased sensitivity to vasodilators also circulating in pregnancy. Interestingly, this finding was specific to the cerebral vasculature, since the effect of pregnant plasma was not seen in mesenteric arteries (3). Thus the adaptation of the cerebral circulation during pregnancy is unique compared with the adaptation of other organ systems. The exact mechanism by which the cerebral vasculature resists the vasoconstrictive effect of circulating factors in pregnancy remains unclear but may involve receptor downregulation or changes in the influence of the endothelium on vascular tone in response to plasma (16, 33). Regardless, this adaptation of the cerebral circulation likely occurs to prevent the cerebrovasculature from constricting in response to circulating factors and may help maintain physiological levels of cerebrovascular resistance and blood flow to the brain during pregnancy.
The Cerebral Endothelium and BBB
Vasomotor responses are not the only feature of the cerebral circulation that adapt to pregnancy. The cerebral endothelium that forms the BBB is a complex interface between systemically circulating factors and the delicate microenvironment of the brain. The endothelial cells of the BBB contain specialized high electrical resistance tight junctions and lack fenestrations (79, 84). BBB tight junctions limit the passage of blood constituents into the brain parenchyma by preventing paracellular transport and are highly protective of the brain milieu (73, 81). Cerebral endothelial cells also have a low rate of pinocytosis, which limits the amount of transcellular transport, reinforcing the overall function of the BBB (13, 42, 69). Pregnancy does not affect mRNA expression of the primary tight junction proteins of the BBB, including claudin-1, claudin-5, occludin, and zona occludens-1, since these are similar to the nonpregnant state (26). In addition, paracellular and transcellular transport at the BBB remain unchanged during normal pregnancy, since BBB permeability to solutes does not increase (26). In addition, hydraulic conductivity, an important parameter that relates water movement through the vessel wall in response to hydrostatic pressure, is normally very low in cerebral endothelial cells due to the high electrical resistance tight junctions and low pinocytotic activity (72). Similar to paracellular and transcellular permeability, hydraulic conductivity is not changed during pregnancy (24) but is increased in response to preeclamptic plasma (2). Increased hydraulic conductivity during preeclampsia could promote neurological symptoms since increased BBB permeability has been linked to several pathological states, including preeclampsia and eclampsia, as well as epilepsy (55, 56, 63).
Pregnancy is a state marked by increased circulating permeability factors, including several that are known to promote BBB permeability, yet it is remarkable that no such changes in permeability have been measured. For example, vascular endothelial growth factor (VEGF), a cytokine originally named vascular permeability factor, is secreted by the placenta and elevated during pregnancy in the uteroplacental unit and the maternal circulation (2, 14, 19, 37). VEGF interacts with its receptors VEGFR1 [or FMS-like tyrosine kinase receptor 1 (Flt-1)] and VEGFR2 [or fetal liver kinase 1 (Flk-1)] located on vascular endothelium to initiate several critical physiological processes involved in angiogenesis, vascular growth, and endothelial cell survival (34, 76). In addition to VEGF, placental growth factor (PlGF), a member of the VEGF family, is also elevated during pregnancy and contributes to angiogenesis in the uterus and placenta (51). Most notably, VEGF and PlGF are potent vasodilators and increase peripheral microvascular permeability to serum proteins and macromolecules, considered a primary step in preparation for angiogenesis (32, 34, 41, 65), and increase BBB permeability (74). Interestingly, despite elevated circulating VEGF and PlGF during pregnancy, exposure of cerebral vessels to pregnant plasma or serum does not increase BBB permeability (24, 74). Furthermore, VEGF receptor expression in cerebral arteries does not appear to change during pregnancy, suggesting the lack of effect of VEGF and PlGF is not due to downregulation of VEGFRI/II or neuropilin (74). In fact, plasma from late-pregnant rats prevents VEGF-induced increases in BBB permeability (74), likely due to increased levels of soluble Flt-1 (sFlt-1). The selective binding of VEGF and PlGF to sFlt-1 is important for regulating their bioavailability, thus limiting the permeability-promoting effects at the BBB during pregnancy (74). The prevention of circulating permeability factors from increasing BBB permeability is an important adaptation during pregnancy to help maintain brain homeostasis.
Although paracellular and transcellular permeability of the BBB appear to remain intact during pregnancy, efflux transporters present at the BBB are an important regulatory mechanism controlling passage of serum factors into the brain that appear to be gestationally regulated. Specifically, p-glycoprotein (Pgp) is a main efflux transporter at the BBB that extricates steroids, cytokines, and chemokines, as well as many pharmacologic agents that can pass through the BBB, essentially acting as a gatekeeper to the central nervous system (10, 79). Pgp is an obstacle in administration of therapeutics to the brain, making it difficult for pharmacological interventions to be delivered in patients with epilepsy, brain tumors, HIV, etc. (10). The role of Pgp in restricting drug delivery to the brain during pregnancy has been investigated in pregnant mice and in nonhuman primates. Pgp protein expression is elevated at the BBB mid-gestation but returns to pre-pregnancy levels by late-gestation in mice (29). In the same study, the protein expression of another efflux transporter, multi-drug resistance-associated protein 1 (Mrp1), was also elevated at the BBB midpregnancy and remained higher late in gestation (29). A study using positron emission tomography scanning to investigate Pgp activity at the BBB across gestation in nonhuman primates reported that Pgp activity increases with gestational age (20). Thus it appears that efflux transporters are gestationally regulated, potentially in response to the increase in circulating factors occurring during pregnancy (7, 30). Although it appears that Pgp expression increases at the BBB only in mid-gestation, its activity may increase late in pregnancy. This potential adaptation may play a key role in maintaining barrier function despite increases in circulating factors, some of which are hormones and steroids that can pass through the BBB due to their lipophilic nature. Overall, increases in efflux transporter expression and/or activity across gestation are likely a critical adaptation of the BBB to prevent passage of circulating factors into the brain during pregnancy.
Preservation of BBB properties and adaptation of efflux transporters in the face of elevated circulating permeability factors during pregnancy appears to be highly protective and may be central to seizure prevention. Seizure-provoking serum constituents are also present late in gestation. Serum from late-pregnant but not nonpregnant rats causes hyperexcitability of hippocampal neuronal networks in cultured slices, measured by evoked field potentials (24). The increase in excitability is due to serum factors causing neuroinflammation via activation of microglia and secretion of tumor necrosis factor-α (TNF-α) (24, 70). However, under normal conditions, microglia are not active in the brain during pregnancy (48). In addition, the brain is not likely to come into contact with circulating serum factors due to the protective nature of the BBB (48, 79).
An important finding is that the brain also appears to be in a state of increased seizure susceptibility during pregnancy (48). Although not completely understood, seizure susceptibility during pregnancy may be due to high levels of neurosteroids that decrease gamma-aminobutyric acid A (GABAA) receptor subunits in the brain, the main inhibitory receptor on neurons, resulting in increased excitability of neuronal networks and lower seizure threshold (48, 54). In addition to direct modulation of neuronal excitability, aquaporin 4 (AQP4) located in astrocytic endfeet is involved in potassium siphoning and maintaining neuronal excitability as well. AQP4 is significantly increased in mid- and late-gestation, and may also be responsible for decreased seizure threshold (12, 68, 80). Despite mechanisms that produce neuronal hyperexcitability and decrease seizure threshold during normal pregnancy, the vast majority of women do not have seizure activity during pregnancy. Thus it appears that the BBB is critical in limiting the passage of harmful seizure-provoking serum constituents into the hyperexcitable maternal brain. Whether BBB properties are sufficient to maintain homeostasis or there is an adaptation of the efflux transporters during pregnancy that prevents passage of seizure-provoking factors is currently unknown. However, it is possible that failure of the efflux transporters to adapt and expel high levels of circulating factors during pregnancy may be one mechanism by which de novo seizure occurs during pregnancy and preeclampsia. FIGURE 1 summarizes our current understanding of the adaptation of the BBB during normal pregnancy.
FIGURE 1.
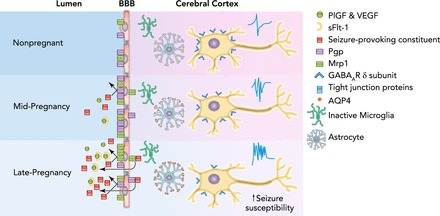
Adaptive response to pregnancy at the blood-brain barrier
Throughout gestation, the maternal circulation is flooded with many serum factors, including permeability factors and seizure-provoking constituents (24). Simultaneously, expression and/or activity of main efflux transporters Pgp and Mrp1 increase at the BBB, potentially in response to increased circulating factors that serves to protect the brain from these factors (20, 29). Black arrows indicate expulsion of circulating factors by Pgp and Mrp1 during pregnancy and protection of the brain from such seizure-provoking factors. Circulating permeability factors VEGF and PlGF increase during pregnancy but are unable to exert an effect on BBB permeability due to concurrent increases in sFlt-1 that limit their bioavailability (14, 76). Pregnancy does not affect tight junction proteins or paracellular permeability of the BBB (26). Centrally, microglia remain in an inactive state; however, AQP4 expression on astrocytes is increased over the course of gestation (48, 68). Furthermore, GABAA receptor subunits are downregulated in the cerebral cortex and hippocampus, resulting in increased neuronal excitability (48, 54).
CBF Autoregulation and Hemodynamics
The cerebral circulation ultimately functions to deliver oxygen, glucose, and nutrient-rich blood to, and remove metabolic waste from, the central nervous system that is crucial to ensure proper brain function. It is therefore not surprising that blood flow autoregulation is well developed in the brain. CBF autoregulation is the intrinsic property of the brain to maintain relatively constant blood flow in the face of changes in blood pressure (45, 46). In normal healthy adults, CBF autoregulation operates between ∼60 and 160 mmHg (52, 60). Pregnancy appears to extend both the upper and the lower limits of the CBF autoregulatory curve. In normal pregnant rats, the limits of the CBF autoregulatory curve were investigated either by inducing controlled hemorrhage to lower blood pressure or by phenylephrine infusion to acutely raise blood pressure together with continuous CBF measurements using laser Doppler flowmetry (18, 22). These studies found that, compared with the nonpregnant state, both the lower and upper limits of CBF autoregulation were shifted (57, 58). FIGURE 2 shows the extended autoregulatory curve in the pregnant state compared with the autoregulatory range in the nonpregnant state (52, 66). Taken together, the effect of pregnancy on CBF autoregulation appears to be protective, making the maternal brain better prepared to maintain blood flow in the face of both acute hypotension and hypertension. However, these studies used animal models of pregnancy since such measurements in pregnant women are challenging and potentially dangerous. Studies in humans using non-invasive techniques to measure dynamic autoregulation during normal pregnancy have found improved or no change in CBF autoregulation in pregnant compared with nonpregnant women (47). To our knowledge, no study has determined the limits of CBF autoregulation during pregnancy, but it is important to understand because of the potential for acute hypotensive and hypertensive episodes that exist, especially during parturition. For example, hemorrhage can occur during parturition, causing hypotension, or parturition can also lead to increased sympathetic discharge, resulting in an acute elevation in blood pressure (67).
FIGURE 2.
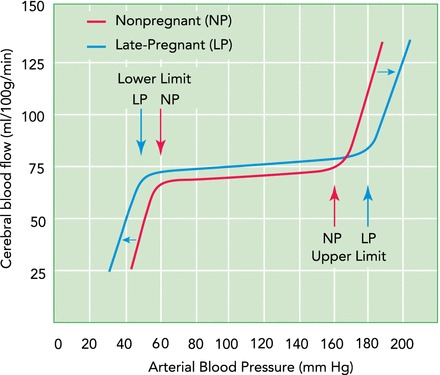
Cerebral blood flow autoregulatory curves in the pregnant and nonpregnant state
Within the arterial pressure range of ∼60–160 mmHg, cerebral blood flow is relatively constant due to cerebral arteries and arterioles that constrict in response to increased pressure and dilate in response to decreased pressure (52, 60). At pressures in excess of the upper limit of autoregulation, forced dilatation of cerebral arteries and arterioles occurs, causing hyperperfusion and a linear relationship between flow and pressure (52). At pressures below the lower limit of autoregulation, flow drops in response to decreased pressure when vasodilation of cerebral vessels cannot compensate (52). The drop of cerebral blood flow during hypotension can cause ischemic brain injury. Impaired autoregulation of cerebral blood flow occurs at both pressure extremes, and flow becomes linearly related to pressure. Pregnancy extends the cerebral blood flow autoregulatory curve at both ends, shifting both the lower and upper limits modestly (18, 22). The leftward shift of the lower limit may be due to the enhanced vasodilation of cerebral arteries that occurs during pregnancy in response to decreased intravascular pressure, whereas the outward remodeling of penetrating arterioles may contribute to the rightward shift in the upper limit. Overall, the extension of autoregulation to higher and lower pressures may be protective of the brain, especially during parturition. There is no significant change in basal cerebral blood flow in the pregnant compared with the nonpregnant state, which is only ∼10% increased globally (15, 26).
Numerous recent studies have also measured changes in basal CBF during pregnancy, employing several techniques including ultrasonography and magnetic resonance (MR) studies. Early human studies using inhalation of a gaseous mixture of nitrous oxide, oxygen, and nitrogen and the Fick principle to assess CBF, oxygen delivery, and metabolism in the brain reported no differences in CBF between the pregnant and nonpregnant states (58, 59). Transcranial Doppler (TCD) studies have measured CBF velocity in cerebral arteries in pregnant women across gestation that revealed CBF velocity decreases during gestation (11, 75, 82). However, changes in vascular resistance and blood flow calculated from these measurements may not accurately reflect CBF due to the lack of information about vessel diameter (49). Using dual-beam angle-independent digital Doppler ultrasonography, diameter and blood flow volume of the internal carotid artery were measured during pregnancy. This study found that CBF increased ∼20% across gestation based on these measurements (62). In contrast, a study using MR reported an ∼20% decrease in CBF during pregnancy; however, this was compared with postpartum and not pre-pregnancy values (83). Studies have used microspheres to measure absolute CBF in late-pregnant rats and found a nonsignificant 5–10% increase in CBF compared with the nonpregnant state (15, 26). Overall, although studies reporting the effect of pregnancy on CBF in women are contradictory, the use of animals that allowed invasive measurements suggests CBF remains similar in pregnancy to the nonpregnant state.
Function and Structure of the Cerebrovasculature
Understanding changes occurring in the structure and function of cerebral arteries and arterioles during pregnancy may shed some light on potential changes in vascular resistance that may drive changes in CBF and autoregulation. Cerebral arteries and arterioles exist in a state of partial constriction and thus have basal tone. A major contributor to basal tone in the cerebral circulation is the myogenic response of vascular smooth muscle cells (9, 53). Pregnancy does not appear to affect myogenic tone in cerebral pial arteries or penetrating brain arterioles (16, 26). Although myogenic tone refers to the degree of basal constriction of a vessel relative to its passive diameter at a constant pressure, the myogenic response refers to the dynamic response of cerebral arteries and arterioles to changes in intravascular pressure (50). The myogenic response is a main contributor to CBF autoregulation through increasing and decreasing cerebrovascular resistance in response to changes in intravascular pressure and appears to be different in the pregnant vs. nonpregnant state (18, 27, 38, 39, 50). In isolated cerebral arteries from pregnant rats, increased intravascular pressure caused forced dilatation at lower pressures than arteries from nonpregnant rats, suggesting a lower capacity to maintain cerebrovascular resistance in the face of increased pressure during pregnancy (27). However, the CBF autoregulatory curve is shifted to higher, not lower, pressures during pregnancy, suggesting other contributors to CBF autoregulation may act in a compensatory way during pregnancy. In contrast, cerebral arteries from pregnant rats had an exaggerated myogenic dilatory response to decreased intravascular pressure, with enhanced vasodilation compared with arteries from nonpregnant rats (18). This may be a protective mechanism by which the maternal cerebrovasculature responds to reductions in intravascular pressure to better maintain blood flow and may contribute to the leftward shift of the lower limit of the autoregulatory curve during pregnancy.
The vasculature of many organ systems, particularly within the uteroplacental circulation, changes structurally to accommodate the physiological adaptation of normal pregnancy (64). There is evidence that the cerebrovasculature also structurally remodels during pregnancy in a selective manner. Structural remodeling describes changes in luminal diameter and vascular wall thickness in response to physiological or pathological stimuli (57). Remodeling can be directed outward or inward, depending on whether the luminal diameter increases or decreases (61). Furthermore, remodeling can be hypo-, hyper-, or eutrophic, depending on whether the vessel wall thickness decreases, increases, or stays the same, respectively (61). During rat pregnancy, no changes in luminal diameter or wall thickness have been measured in cerebral pial arteries, suggesting pregnancy-induced remodeling does not occur in the pial vasculature (16). However, brain parenchymal arterioles, precapillary resistance vessels that branch off pial vessels and perfuse the brain tissue, undergo outward hypotrophic remodeling during pregnancy, resulting in larger vascular lumens and thinner vessel walls than in the nonpregnant state (26). Although there is no change in myogenic tone in these vessels during pregnancy, the intravascular pressure vs. luminal diameter curve is shifted upward due to the structural changes (26). This selective remodeling of parenchymal arterioles during pregnancy is through arteriogenesis and is driven by peroxisome proliferator-activated receptor gamma (PPARγ) activation by the hormone (ser)relaxin (17). Interestingly, the primary relaxin receptor is not expressed in parenchymal arterioles (17). However, circulating relaxin appears to cross the BBB and is thought to activate PPARγ on astrocytes and neurons that in turn exert a paracrine effect on parenchymal arterioles to drive outward remodeling (17), as illustrated in FIGURE 3. In contrast to parenchymal arterioles, pial arteries are not intimately associated with brain parenchymal cell types such as astrocytes and neurons, and this may explain the selective effect of pregnancy-induced remodeling on parenchymal arterioles. This process of arteriogenesis is unique compared with angiogenesis; however, angiogenesis also occurs in the brain during pregnancy. In fact, capillary density increases during pregnancy in the posterior cerebral cortex, a finding that has also been linked to increased activation of PPARγ by relaxin (17, 26). Thus, although pregnancy does not affect the structure of the pial vasculature, it seems to have an outward hypotrophic remodeling effect on parenchymal arterioles that may contribute to the extension of the CBF autoregulatory curve.
FIGURE 3.
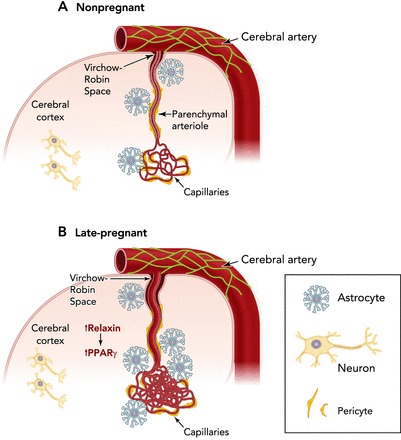
Structural changes of cerebral arteries and arterioles during pregnancy
The cerebral arteries on the brain surface undergo little to no change in luminal diameter or vessel wall thickness during pregnancy. However, intraparenchymal arterioles that branch off the pial surface arteries and perfuse the brain parenchyma undergo outward hypotrophic remodeling, resulting in larger vascular lumens and thinner vessel walls compared with the nonpregnant state (26). Structural remodeling of cerebral arterioles is through a mechanism involving increased circulating relaxin that crosses the BBB and activates PPARγ on astrocytes and neurons, production of VEGF, and activation of matrix metalloproteinases (17, 26). In addition, capillary density increases in the posterior brain region during pregnancy through a similar mechanism (26). These changes may be occurring due to increased demand of blood supply in neuroendocrine glands that enlarge during pregnancy (e.g., the pituitary gland) (43) or areas involved in production and/or secretion of pregnancy hormones. The increase in vascular volume of the cerebral circulation during pregnancy combined with expanded plasma volume may underlie the susceptibility to edema formation during acute hypertension and other forms of brain injury such as seizure. Figure was adapted with permission from Ref. 26.
During pregnancy, outward remodeling of parenchymal arterioles and increased capillary density coupled with the ∼10% hemodilution that occurs could decrease cerebrovascular resistance and increase CBF (44). Despite these vascular and hemodynamic changes, pregnancy has little effect on cerebrovascular resistance when measured under normotensive conditions in the rat (26). The substantial contribution of large cerebral arteries to vascular resistance is unique to the cerebral circulation (38). In fact, large extracranial and intracranial cerebral arteries contribute ∼50% of cerebrovascular resistance (38, 40). Although downstream arterioles undergo structural changes that may decrease small vessel resistance, the pial vasculature does not change structurally during pregnancy and may compensate to maintain normal vascular resistance (FIGURE 3). The structural remodeling of the cerebral circulation may be important when hypertensive pathologies of pregnancy such as preeclampsia are considered. Under experimental conditions of acute hypertension, cerebrovascular resistance of cerebral arteries was decreased in pregnant rats, leading to autoregulatory breakthrough and an ∼40% increase in CBF (26). The decrease in cerebrovascular resistance with autoregulatory breakthrough was further associated with increased BBB permeability due to greater hydrostatic pressure on the microcirculation (26). Thus, although the BBB seems to remain intact during normal pregnancy under physiological conditions, it also appears to be at greater risk of injury during pathological states such as acute hypertension. Under conditions of elevated intravascular pressure when large arteries become ineffective in regulating CBF due to forced dilatation of myogenic tone, outward hypotrophic remodeling of parenchymal arterioles may predispose the microcirculation to injury by transmitting high hydrostatic pressure downstream. Outward hypotrophic remodeling of brain arterioles could also contribute to the maternal brain being more sensitive to vasogenic edema formation after acute hypertension (22, 36). The increase in capillary density may further contribute to the susceptibility of the brain to hypertension-induced vasogenic edema during pregnancy by increasing the potential sites of BBB disruption. In addition, pregnancy both prevents and reverses remodeling of cerebral arteries that occurs in response to chronic hypertension (5, 23, 25). Chronic hypertension in the nonpregnant state leads to inward hypertrophic remodeling of cerebral arteries, resulting in smaller lumen diameters and thicker vascular walls (8, 23). This is considered a protective adaptation by which cerebrovascular resistance is increased, thus protecting the microcirculation from elevated arterial blood pressure (16, 23). The prevention and/or reversal of this remodeling during pregnancy may be related to the downregulation of angiotensin type 1 receptor (AT1R) expression in the cerebral circulation that also occurs during pregnancy (16). Although the reduction in AT1R expression may be a physiological adaptation to normal pregnancy, reversal and/or prevention of hypertensive remodeling may make the cerebral microcirculation even more susceptible to injury during states associated with hypertension. Again, this may be important during preeclampsia, where acute elevations in blood pressure are thought to lead to cerebral vasogenic edema formation and subsequent neurological complications, including eclamptic seizure (21).
The adaptation of parenchymal arterioles during pregnancy may be further implicated in neurological complications associated with preeclampsia and eclampsia. Parenchymal arterioles are the primary vessels involved in small vessel disease in the brain (71). Women with eclampsia seem to be prone to white matter lesions later in life, which may indicate the presence of small vessel disease in the brain (4). It is possible, albeit speculative at this time, that failure of parenchymal arterioles to outward remodel during preeclampsia underlies the potential for white matter lesions later in life. Lack of spiral artery remodeling during pregnancy is a feature of some women with preeclampsia that may indicate a type of small vessel disease (35). We further speculate that the lack of adaptation of small vessels in the uterine circulation may be similar to the lack of adaptation of the small vessels in the brain, both of which may be occurring in women with severe preeclampsia and eclampsia. Furthermore, if the lack of remodeling occurs in parenchymal arterioles in the brain, as it does in the uterine circulation in some eclamptic women, this may suggest a common pathology that leads to white matter lesions and cognitive impairment associated with eclampsia (4, 6). However, the association of small vessel disease of the brain later in life in formerly eclamptic women with impaired spiral artery remodeling during pregnancy is speculative, and further studies are needed to understand these processes.
Summary
Pregnancy is associated with many adaptations of the cerebral circulation, including changes in receptor and transporter activity, keeping increased permeability factors in balance to maintain brain homeostasis, and protecting against increases in BBB permeability. Furthermore, structural and functional changes occur in certain segments of the cerebral vasculature; however, CBF and cerebrovascular resistance appear unchanged under normotensive conditions. The CBF autoregulation curve appears to extend in the pregnant state, protecting the maternal brain against acute and drastic fluctuations in blood pressure. It is remarkable that BBB permeability and CBF are affected so minimally during pregnancy, especially in the face of substantially increased factors that have direct effects on vascular filtration and flow in many other organ systems. However, this supports the principle that the adaptation of the cerebral circulation to normal pregnancy functions to maintain essential oxygen and nutrient delivery and waste removal similar to the nonpregnant state, especially in the face of tremendous systemic hemodynamic changes associated with pregnancy.
Footnotes
No conflicts of interest, financial or otherwise, are declared by the author(s).
Author contributions: A.J. and M.C. prepared figures; A.J. and M.C. drafted manuscript; A.J. and M.C. edited and revised manuscript; A.J. and M.C. approved final version of manuscript.
References
- 1.Aagaard-Tillery KM, Silver R, Dalton J. Immunology of normal pregnancy. Semin Fetal Neonatal Med 11: 279–295, 2006. [DOI] [PubMed] [Google Scholar]
- 2.Amburgey OA, Chapman AC, May V, Bernstein IM, Cipolla MJ. Plasma from preeclamptic women increases blood-brain barrier permeability: role of vascular endothelial growth factor signaling. Hypertension 56: 1003–1008, 2010. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 3.Amburgey OA, Reeves SA, Bernstein IM, Cipolla MJ. Resistance artery adaptation to pregnancy counteracts the vasoconstricting influence of plasma from normal pregnant women. Reprod Sci 17: 29–39, 2010. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 4.Aukes AM, de Groot JC, Aarnoudse JG, Zeeman GG. Brain lesions several years after eclampsia. Am J Obstet Gynecol 200: e501–e505, 2009. [DOI] [PubMed] [Google Scholar]
- 5.Aukes AM, Vitullo L, Zeeman GG, Cipolla MJ. Pregnancy prevents hypertensive remodeling and decreases myogenic reactivity in posterior cerebral arteries from Dahl salt-sensitive rats: a role in eclampsia? Am J Physiol Heart Circ Physiol 292: H1071–H1076, 2007. [DOI] [PubMed] [Google Scholar]
- 6.Aukes AM, Wessel I, Dubois AM, Aarnoudse JG, Zeeman GG. Self-reported cognitive functioning in formerly eclamptic women. Am J Obstet Gynecol 197: e361–e366, 2007. [DOI] [PubMed] [Google Scholar]
- 7.Bauer B, Hartz AM, Miller DS. Tumor necrosis factor alpha and endothelin-1 increase P-glycoprotein expression and transport activity at the blood-brain barrier. Mol Pharmacol 71: 667–675, 2007. [DOI] [PubMed] [Google Scholar]
- 8.Baumbach GL, Heistad DD. Remodeling of cerebral arterioles in chronic hypertension. Hypertension 13: 968–972, 1989. [DOI] [PubMed] [Google Scholar]
- 9.Bayliss WM. On the local reactions of the arterial wall to changes of internal pressure. J Physiol 28: 220–231, 1902. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 10.Begley DJ. ABC transporters and the blood-brain barrier. Curr Pharm Des 10: 1295–1312, 2004. [DOI] [PubMed] [Google Scholar]
- 11.Belfort MA, Tooke-Miller C, Allen JC Jr, Saade GR, Dildy GA, Grunewald C, Nisell H, Herd JA. Changes in flow velocity, resistance indices, and cerebral perfusion pressure in the maternal middle cerebral artery distribution during normal pregnancy. Acta Obstet Gynecol Scand 80: 104–112, 2001. [PubMed] [Google Scholar]
- 12.Binder DK, Oshio K, Ma T, Verkman AS, Manley GT. Increased seizure threshold in mice lacking aquaporin-4 water channels. Neuroreport 15: 259–262, 2004. [DOI] [PubMed] [Google Scholar]
- 13.Brightman MW, Reese TS. Junctions between intimately apposed cell membranes in the vertebrate brain. J Cell Biol 40: 648–677, 1969. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 14.Brown LF, Detmar M, Claffey K, Nagy JA, Feng D, Dvorak AM, Dvorak HF. Vascular permeability factor/vascular endothelial growth factor: a multifunctional angiogenic cytokine. EXS 79: 233–269, 1997. [DOI] [PubMed] [Google Scholar]
- 15.Buelke-Sam J, Nelson CJ, Byrd RA, Holson JF. Blood flow during pregnancy in the rat: I. Flow patterns to maternal organs. Teratology 26: 269–277, 1982. [DOI] [PubMed] [Google Scholar]
- 16.Chan SL, Chapman AC, Sweet JG, Gokina NI, Cipolla MJ. Effect of PPARgamma inhibition during pregnancy on posterior cerebral artery function and structure. Front Physiol 1: 130, 2010. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 17.Chan SL, Cipolla MJ. Relaxin causes selective outward remodeling of brain parenchymal arterioles via activation of peroxisome proliferator-activated receptor-gamma. FASEB J 25: 3229–3239, 2011. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 18.Chapman AC, Cipolla MJ, Chan SL. Effect of pregnancy and nitric oxide on the myogenic vasodilation of posterior cerebral arteries and the lower limit of cerebral blood flow autoregulation. Reprod Sci 20: 1046–1054, 2013. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 19.Charnock-Jones DS, Kaufmann P, Mayhew TM. Aspects of human fetoplacental vasculogenesis and angiogenesis. I. Molecular regulation. Placenta 25: 103–113, 2004. [DOI] [PubMed] [Google Scholar]
- 20.Chung FS, Eyal S, Muzi M, Link JM, Mankoff DA, Kaddoumi A, O'Sullivan F, Hsiao P, Unadkat JD. Positron emission tomography imaging of tissue P-glycoprotein activity during pregnancy in the non-human primate. Br J Pharmacol 159: 394–404, 2010. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 21.Cipolla MJ. Cerebrovascular function in pregnancy and eclampsia. Hypertension 50: 14–24, 2007. [DOI] [PubMed] [Google Scholar]
- 22.Cipolla MJ, Bishop N, Chan SL. Effect of pregnancy on autoregulation of cerebral blood flow in anterior versus posterior cerebrum. Hypertension 60: 705–711, 2012. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 23.Cipolla MJ, DeLance N, Vitullo L. Pregnancy prevents hypertensive remodeling of cerebral arteries: a potential role in the development of eclampsia. Hypertension 47: 619–626, 2006. [DOI] [PubMed] [Google Scholar]
- 24.Cipolla MJ, Pusic AD, Grinberg YY, Chapman AC, Poynter ME, Kraig RP. Pregnant serum induces neuroinflammation and seizure activity via TNFalpha. Exp Neurol 234: 398–404, 2012. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 25.Cipolla MJ, Smith J, Bishop N, Bullinger LV, Godfrey JA. Pregnancy reverses hypertensive remodeling of cerebral arteries. Hypertension 51: 1052–1057, 2008. [DOI] [PubMed] [Google Scholar]
- 26.Cipolla MJ, Sweet JG, Chan SL. Cerebral vascular adaptation to pregnancy and its role in the neurological complications of eclampsia. J Appl Physiol 110: 329–339, 2011. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 27.Cipolla MJ, Vitullo L, McKinnon J. Cerebral artery reactivity changes during pregnancy and the postpartum period: a role in eclampsia? Am J Physiol Heart Circ Physiol 286: H2127–H2132, 2004. [DOI] [PubMed] [Google Scholar]
- 28.Clapp JF 3rd, Capeless E. Cardiovascular function before, during, and after the first and subsequent pregnancies. Am J Cardiol 80: 1469–1473, 1997. [DOI] [PubMed] [Google Scholar]
- 29.Coles LD, Lee IJ, Hassan HE, Eddington ND. Distribution of saquinavir, methadone, and buprenorphine in maternal brain, placenta, and fetus during two different gestational stages of pregnancy in mice. J Pharm Sci 98: 2832–2846, 2009. [DOI] [PubMed] [Google Scholar]
- 30.Coles LD, Lee IJ, Voulalas PJ, Eddington ND. Estradiol and progesterone-mediated regulation of P-gp in P-gp overexpressing cells (NCI-ADR-RES) and placental cells (JAR). Mol Pharm 6: 1816–1825, 2009. [DOI] [PubMed] [Google Scholar]
- 31.Dewerchin M, Carmeliet P. PlGF: a multitasking cytokine with disease-restricted activity. Cold Spring Harb Perspect Med 2: a011056, 2012. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 32.Dobrogowska DH, Lossinsky AS, Tarnawski M, Vorbrodt AW. Increased blood-brain barrier permeability and endothelial abnormalities induced by vascular endothelial growth factor. J Neurocytol 27: 163–173, 1998. [DOI] [PubMed] [Google Scholar]
- 33.Duckles SP, Krause DN. Cerebrovascular effects of oestrogen: multiplicity of action. Clin Exp Pharmacol Physiol 34: 801–808, 2007. [DOI] [PubMed] [Google Scholar]
- 34.Dvorak HF. Vascular permeability factor/vascular endothelial growth factor: a critical cytokine in tumor angiogenesis and a potential target for diagnosis and therapy. J Clin Oncol 20: 4368–4380, 2002. [DOI] [PubMed] [Google Scholar]
- 35.Egbor M, Ansari T, Morris N, Green CJ, Sibbons PD. Morphometric placental villous and vascular abnormalities in early- and late-onset pre-eclampsia with and without fetal growth restriction. BJOG 113: 580–589, 2006. [DOI] [PubMed] [Google Scholar]
- 36.Euser AG, Cipolla MJ. Cerebral blood flow autoregulation and edema formation during pregnancy in anesthetized rats. Hypertension 49: 334–340, 2007. [DOI] [PubMed] [Google Scholar]
- 37.Evans P, Wheeler T, Anthony F, Osmond C. Maternal serum vascular endothelial growth factor during early pregnancy. Clin Sci (Lond) 92: 567–571, 1997. [DOI] [PubMed] [Google Scholar]
- 38.Faraci FM, Heistad DD. Regulation of large cerebral arteries and cerebral microvascular pressure. Circ Res 66: 8–17, 1990. [DOI] [PubMed] [Google Scholar]
- 39.Faraci FM, Heistad DD, Mayhan WG. Role of large arteries in regulation of blood flow to brain stem in cats. J Physiol 387: 115–123, 1987. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 40.Faraci FM, Mayhan WG, Heistad DD. Segmental vascular responses to acute hypertension in cerebrum and brain stem. Am J Physiol Heart Circ Physiol 252: H738–H742, 1987. [DOI] [PubMed] [Google Scholar]
- 41.Feng D, Nagy JA, Hipp J, Dvorak HF, Dvorak AM. Vesiculo-vacuolar organelles and the regulation of venule permeability to macromolecules by vascular permeability facto, histamine, and serotonin. J Exp Med 183: 1981–1986, 1996. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 42.Fenstermacher J, Gross P, Sposito N, Acuff V, Pettersen S, Gruber K. Structural and functional variations in capillary systems within the brain. Ann NY Acad Sci 529: 21–30, 1988. [DOI] [PubMed] [Google Scholar]
- 43.Gonzalez JG, Elizondo G, Saldivar D, Nanez H, Todd LE, Villarreal JZ. Pituitary gland growth during normal pregnancy: an in vivo study using magnetic resonance imaging. Am J Med 85: 217–220, 1988. [DOI] [PubMed] [Google Scholar]
- 44.Gordon Maternal Physiology MC. In: Gabbe SG, Niebyl JR, Simpson JL (editors). Obstetrics: Normal and Problem Pregnancies (5th ed.). London: Churchill Liverstone, 2007. [Google Scholar]
- 45.Harper AM. Autoregulation of cerebral blood flow: influence of the arterial blood pressure on the blood flow through the cerebral cortex. J Neurol Neurosurg Psychiatry 29: 398–403, 1966. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 46.Hayman LA, Berman SA, Hinck VC. Correlation of CT cerebral vascular territories with function: II. Posterior cerebral artery. AJR Am J Roentgenol 137: 13–19, 1981. [DOI] [PubMed] [Google Scholar]
- 47.Janzarik WG, Ehlers E, Ehmann R, Gerds TA, Schork J, Mayer S, Gabriel B, Weiller C, Prompeler H, Reinhard M. Dynamic cerebral autoregulation in pregnancy and the risk of preeclampsia. Hypertension 63: 161–166, 2014. [DOI] [PubMed] [Google Scholar]
- 48.Johnson A, Tremble S, Cipolla M. Decreased seizure threshold during pregnancy and experimental preeclampsia: roles for GABAA receptors and microglial activation. Reprod Sci 21: 100A, 2014. [Google Scholar]
- 49.Kontos HA. Validity of cerebral arterial blood flow calculations from velocity measurements. Stroke 20: 1–3, 1989. [DOI] [PubMed] [Google Scholar]
- 50.Kontos HA, Wei EP, Navari RM, Levasseur JE, Rosenblum WI, Patterson JL Jr. Responses of cerebral arteries and arterioles to acute hypotension and hypertension. Am J Physiol Heart Circ Physiol 234: H371–H383, 1978. [DOI] [PubMed] [Google Scholar]
- 51.Krauss T, Pauer HU, Augustin HG. Prospective analysis of placenta growth factor (PlGF) concentrations in the plasma of women with normal pregnancy and pregnancies complicated by preeclampsia. Hypertens Pregnancy 23: 101–111, 2004. [DOI] [PubMed] [Google Scholar]
- 52.Lassen NA. Cerebral blood flow and oxygen consumption in man. Physiol Rev 39: 183–238, 1959. [DOI] [PubMed] [Google Scholar]
- 53.MacKenzie ET, Farrar JK, Fitch W, Graham DI, Gregory PC, Harper AM. Effects of hemorrhagic hypotension on the cerebral circulation. I. Cerebral blood flow and pial arteriolar caliber. Stroke 10: 711–718, 1979. [DOI] [PubMed] [Google Scholar]
- 54.Maguire J, Ferando I, Simonsen C, Mody I. Excitability changes related to GABAA receptor plasticity during pregnancy. J Neurosci 29: 9592–9601, 2009. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 55.Marchi N, Angelov L, Masaryk T, Fazio V, Granata T, Hernandez N, Hallene K, Diglaw T, Franic L, Najm I, Janigro D. Seizure-promoting effect of blood-brain barrier disruption. Epilepsia 48: 732–742, 2007. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 56.Marchi N, Tierney W, Alexopoulos AV, Puvenna V, Granata T, Janigro D. The etiological role of blood-brain barrier dysfunction in seizure disorders. Cardiovasc Psychiatry Neurol 2011: 482415, 2011. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 57.Martinez-Lemus LA, Hill MA, Meininger GA. The plastic nature of the vascular wall: a continuum of remodeling events contributing to control of arteriolar diameter and structure. Physiology 24: 45–57, 2009. [DOI] [PubMed] [Google Scholar]
- 58.McCall M. Cerebral blood flow and metabolism in toxemias of pregnancy. Surg Gynecol Obstet 89: 715–721, 1949. [PubMed] [Google Scholar]
- 59.McCall M. Cerebral circulation and metabolism in toxemia of pregnancy; observations on the effects of veratrum viride and apresoline (1-hydrazinophthalazine). Am J Obstet Gynecol 66: 1015–1030, 1953. [DOI] [PubMed] [Google Scholar]
- 60.McHenry LC Jr, West JW, Cooper ES, Goldberg HI, Jaffe ME. Cerebral autoregulation in man. Stroke 5: 695–706, 1974. [DOI] [PubMed] [Google Scholar]
- 61.Mulvany MJ. Vascular remodelling of resistance vessels: can we define this? Cardiovasc Res 41: 9–13, 1999. [DOI] [PubMed] [Google Scholar]
- 62.Nevo O, Soustiel JF, Thaler I. Maternal cerebral blood flow during normal pregnancy: a cross-sectional study. Am J Obstet Gynecol 203: e471–e476, 2010. [DOI] [PubMed] [Google Scholar]
- 63.Oby E, Janigro D. The blood-brain barrier and epilepsy. Epilepsia 47: 1761–1774, 2006. [DOI] [PubMed] [Google Scholar]
- 64.Osol G, Mandala M. Maternal uterine vascular remodeling during pregnancy. Physiology 24: 58–71, 2009. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 65.Oura H, Bertoncini J, Velasco P, Brown LF, Carmeliet P, Detmar M. A critical role of placental growth factor in the induction of inflammation and edema formation. Blood 101: 560–567, 2003. [DOI] [PubMed] [Google Scholar]
- 66.Paulson OB, Strandgaard S, Edvinsson L. Cerebral autoregulation. Cerebrovasc Brain Metab Rev 2: 161–192, 1990. [PubMed] [Google Scholar]
- 67.Pickering TG. Effects of stress and behavioral interventions in hypertension. Pain and blood pressure. J Clin Hypertens (Greenwich) 5: 359–361, 2003. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 68.Quick AM, Cipolla MJ. Pregnancy-induced up-regulation of aquaporin-4 protein in brain and its role in eclampsia. FASEB J 19: 170–175, 2005. [DOI] [PubMed] [Google Scholar]
- 69.Reese TS, Karnovsky MJ. Fine structural localization of a blood-brain barrier to exogenous peroxidase. J Cell Biol 34: 207–217, 1967. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 70.Riazi K, Galic MA, Kuzmiski JB, Ho W, Sharkey KA, Pittman QJ. Microglial activation and TNFalpha production mediate altered CNS excitability following peripheral inflammation. Proc Natl Acad Sci USA 105: 17151–17156, 2008. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 71.Rincon F, Wright CB. Current pathophysiological concepts in cerebral small vessel disease. Front Aging Neurosci 6: 24, 2014. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 72.Roberts TJ, Chapman AC, Cipolla MJ. PPAR-gamma agonist rosiglitazone reverses increased cerebral venous hydraulic conductivity during hypertension. Am J Physiol Heart Circ Physiol 297: H1347–H1353, 2009. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 73.Rubin LL, Staddon JM. The cell biology of the blood-brain barrier. Annu Rev Neurosci 22: 11–28, 1999. [DOI] [PubMed] [Google Scholar]
- 74.Schreurs MP, Houston EM, May V, Cipolla MJ. The adaptation of the blood-brain barrier to vascular endothelial growth factor and placental growth factor during pregnancy. FASEB J 26: 355–362, 2012. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 75.Serra-Serra V, Kyle PM, Chandran R, Redman CW. Maternal middle cerebral artery velocimetry in normal pregnancy and postpartum. Br J Obstet Gynaecol 104: 904–909, 1997. [DOI] [PubMed] [Google Scholar]
- 76.Shibuya M. Vascular endothelial growth factor and its receptor system: physiological functions in angiogenesis and pathological roles in various diseases. J Biochem 153: 13–19, 2013. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 77.Siegel GJ. Basic Neurochemistry: Molecular, Cellular, and Medical Aspects (6th ed.). Philadelphia, PA: Lippincott Williams & Wilkins, 1999. [Google Scholar]
- 78.Szarka A, Rigo J Jr, Lazar L, Beko G, Molvarec A. Circulating cytokines, chemokines and adhesion molecules in normal pregnancy and preeclampsia determined by multiplex suspension array. BMC Immunol 11: 59, 2010. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 79.Ueno M. Molecular anatomy of the brain endothelial barrier: an overview of the distributional features. Curr Med Chem 14: 1199–1206, 2007. [DOI] [PubMed] [Google Scholar]
- 80.Verkman AS, Binder DK, Bloch O, Auguste K, Papadopoulos MC. Three distinct roles of aquaporin-4 in brain function revealed by knockout mice. Biochim Biophys Acta 1758: 1085–1093, 2006. [DOI] [PubMed] [Google Scholar]
- 81.Wahl M, Unterberg A, Baethmann A, Schilling L. Mediators of blood-brain barrier dysfunction and formation of vasogenic brain edema. J Cereb Blood Flow Metab 8: 621–634, 1988. [DOI] [PubMed] [Google Scholar]
- 82.Williams K, Wilson S. Maternal middle cerebral artery blood flow velocity variation with gestational age. Obstet Gynecol 84: 445–448, 1994. [PubMed] [Google Scholar]
- 83.Zeeman GG, Hatab M, Twickler DM. Maternal cerebral blood flow changes in pregnancy. Am J Obstet Gynecol 189: 968–972, 2003. [DOI] [PubMed] [Google Scholar]
- 84.Zlokovic BV. The blood-brain barrier in health and chronic neurodegenerative disorders. Neuron 57: 178–201, 2008. [DOI] [PubMed] [Google Scholar]