
Figure 4.

Models of DIC transport in marine diatoms. (a) In P. tricornutum, plasma-membrane-located-SLC4s (PtSLC4-2, possibly PtSLC4-1 and PtSLC4-4) transport 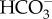 from seawater in a Na+-dependent manner, most probably Na+–
from seawater in a Na+-dependent manner, most probably Na+–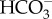 cotransport. This would require an unidentified ATP-dependent primary Na+ pump or a proton-dependent secondary pump to maintain a [Na+] gradient across the plasma membrane. The mechanism of DIC transport from the cytosol to the stroma is unknown. Unidentified transporters on CER and CE might be involved in this process in combination with CAs in intermembrane spaces. In the pyrenoid, CO2 is generated by luminal θ-CA in the pyrenoid-penetrating thylakoid (PPT). CO2 released from the lumen is fixed by RubisCO, or otherwise recaptured at specialized pyrenoidal loci by β-type CAs, PtCA1 and/or PtCA2 by which
cotransport. This would require an unidentified ATP-dependent primary Na+ pump or a proton-dependent secondary pump to maintain a [Na+] gradient across the plasma membrane. The mechanism of DIC transport from the cytosol to the stroma is unknown. Unidentified transporters on CER and CE might be involved in this process in combination with CAs in intermembrane spaces. In the pyrenoid, CO2 is generated by luminal θ-CA in the pyrenoid-penetrating thylakoid (PPT). CO2 released from the lumen is fixed by RubisCO, or otherwise recaptured at specialized pyrenoidal loci by β-type CAs, PtCA1 and/or PtCA2 by which 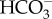 is regenerated to be transported back to the thylakoid lumen. (b) In T. pseudonana, extracellular CA accelerates the dehydration of
is regenerated to be transported back to the thylakoid lumen. (b) In T. pseudonana, extracellular CA accelerates the dehydration of 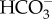 , resulting in high diffusive permeation of CO2. Active
, resulting in high diffusive permeation of CO2. Active 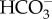 transport into the chloroplast would lower cytoplasmic [
transport into the chloroplast would lower cytoplasmic [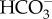 ], and then CO2 that diffuses into the cell is readily converted into
], and then CO2 that diffuses into the cell is readily converted into 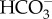 by cytosolic CA (alternatively this could occur in the PPC). As in P. tricornutum, transporters on CER and CE have not been identified. In T. pseudonana, a putative θ-CA is predicted in the PPT, suggesting a role for θ-CA in CO2 generation as proposed in P. tricornutum. There is no known CA localized in the pyrenoid. (c) In C. reinharditii,
by cytosolic CA (alternatively this could occur in the PPC). As in P. tricornutum, transporters on CER and CE have not been identified. In T. pseudonana, a putative θ-CA is predicted in the PPT, suggesting a role for θ-CA in CO2 generation as proposed in P. tricornutum. There is no known CA localized in the pyrenoid. (c) In C. reinharditii, 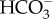 is delivered into the stroma by plasma-membrane-located HLA3 and chloroplast-envelope-located LCIA. LCIA is shown to be on the chloroplast envelope, but its exact localization, whether inner or outer membrane, is unknown. Here, we assumed that LCIA is on the inner envelope because the outer membrane is generally permeable to low molecular weight compounds which most probably allows spontaneous
is delivered into the stroma by plasma-membrane-located HLA3 and chloroplast-envelope-located LCIA. LCIA is shown to be on the chloroplast envelope, but its exact localization, whether inner or outer membrane, is unknown. Here, we assumed that LCIA is on the inner envelope because the outer membrane is generally permeable to low molecular weight compounds which most probably allows spontaneous 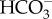 permeation. LCI1 on plasma membrane was strongly suggested to be a DIC transporter, but the inorganic carbon species transported by LCI1 have not been determined. CAH3, a luminal CA in the thylakoid tubule, generates CO2 from
permeation. LCI1 on plasma membrane was strongly suggested to be a DIC transporter, but the inorganic carbon species transported by LCI1 have not been determined. CAH3, a luminal CA in the thylakoid tubule, generates CO2 from 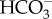 . Unlike P. tricornutum, the luminal CAH3 is α-type CA, suggesting convergent evolution of a mechanism to supply CO2 by using luminal acidification. Similarly, the LCIB-C complex is a putative θ-CA, another example of convergent evolution of pyrenoidal CA which may play a role in recapturing leaking out CO2. CE, chloroplast envelopes; CER, chloroplast ER; PM, plasma membrane; PPC, periplastidal compartment; PPS, periplasmic space.
. Unlike P. tricornutum, the luminal CAH3 is α-type CA, suggesting convergent evolution of a mechanism to supply CO2 by using luminal acidification. Similarly, the LCIB-C complex is a putative θ-CA, another example of convergent evolution of pyrenoidal CA which may play a role in recapturing leaking out CO2. CE, chloroplast envelopes; CER, chloroplast ER; PM, plasma membrane; PPC, periplastidal compartment; PPS, periplasmic space.