

Abstract
Dengue represents a serious threat to human health, with billions of people living at risk of the disease. Wolbachia pipientis is a bacterial endosymbiont common to many insect species. Wolbachia transinfections in mosquito disease vectors have great value for disease control given the bacterium’s ability to spread into wild mosquito populations, and to interfere with infections of pathogens, such as dengue virus. Aedes fluviatilis is a mosquito with a widespread distribution in Latin America, but its status as a dengue vector has not been clarified. Ae. fluviatilis is also naturally infected by the wFlu Wolbachia strain, which has been demonstrated to enhance infection with the avian malarial parasite Plasmodium gallinaceum. We performed experimental infections of Ae. fluviatilis with DENV-2 and DENV-3 isolates from Brazil via injection or oral feeding to provide insight into its competence for the virus. We also examined the effect of the native Wolbachia infection on the virus using a mosquito line where the wFlu infection had been cleared by antibiotic treatment. Through RT-qPCR, we observed that Ae. fluviatilis could become infected with both viruses via either method of infection, although at a lower rate than Aedes aegypti, the primary dengue vector. We then detected DENV-2 and DENV-3 in the saliva of injected mosquitoes, and observed that injection of DENV-3-infected saliva produced subsequent infections in naïve Ae. aegypti. However, across our data we observed no difference in prevalence of infection and viral load between Wolbachia-infected and -uninfected mosquitoes, suggesting that there is no effect of wFlu on dengue virus. Our results highlight that Ae. fluviatilis could potentially serve as a dengue vector under the right circumstances, although further testing is required to determine if this occurs in the field.
Introduction
Dengue virus (DENV) represents a serious threat to human health across the tropical regions of the world. There are four genetically distinct DENV serotypes, and infection can cause dengue, which has fever-like symptoms, or the comparatively rare severe dengue, which can lead to haemorrhaging and death. Subsequent infection with a different serotype greatly increases the risk that severe dengue will develop (WHO). There are an estimated 96 million cases that manifest clinically, each year [1], and nearly 4 billion people living at risk of infection [2], with severe, endemic transmission occurring in Latin America, and South and Southeast Asia.
In the last 40 years there has been an expansion in the incidence and prevalence of dengue cases [3]. This has occurred in conjunction with an increase in the geographic distribution of the mosquito vectors of the disease; the primary vector, Aedes aegypti, and the secondary vector Aedes albopictus [4]. Changes in climate, increased travel to disease-endemic areas, and increased deforestation and urbanization have served to bring humans and mosquitoes into contact more frequently, increasing the chance of viral transmission [1, 5–8]. Over the same time period there has been an increase in the prevalence of genetic resistance to commonly used insecticidal chemicals in mosquito populations, which has led to a decline in the efficacy of historically successful mosquito control programs [9, 10]. Urbanization and deforestation could also lead to a change in the dynamics of a pathogen transmission cycle, by increasing the frequency of contact between humans and other mosquito species that might become infected with the virus in the field [6, 11, 12].
Aedes (Ochlerotatus) fluviatilis (Lutz, 1904) is an anthropophilic species of mosquito native to Latin America, which lives in a wide variety of habitats, including urban and undeveloped areas [13]. Ae. fluviatilis has been described as being competent for infection with yellow fever virus, and was subsequently hypothesised as a potential vector for the disease in Latin America [14]. It has also been described as a good laboratory model for the avian malaria parasite Plasmodium gallinaceum [15]. However, little is known about the species’ ability to maintain infections with or transmit DENV.
Wolbachia pipientis is a maternally transmitted, bacterial endosymbiont that commonly infects many insect species [16]. Wolbachia strains are genetically diverse [17, 18], and maintain intricate relationships with their different host species [19, 20]. Many of these relationships are characterised by symbiont-induced manipulation of host reproduction, which allow the bacterium to persist in insect populations even in the face of fitness costs, and can permit the bacterium to spread through uninfected populations [21–24].
Many mosquito species are naturally infected by Wolbachia, including Ae. fluviatilis, which is infected by the wFlu strain [25, 26]. However, several key vector species, including most Ae. aegypti populations [27], are naturally uninfected. In these cases, stable infections have been generated by transinfection—the transfer of Wolbachia between species to form a stable infection [28]. Ae. aegypti has been independently transinfected with multiple Wolbachia strains [29–31], including the wMel strain, originally found in the fruit fly Drosophila melanogaster [32]. Wolbachia transinfections in mosquitoes are strongly associated with pathogen interference—the ability to interfere with the replication and infection of pathogens such as the dengue and Zika viruses, and the human malarial parasite Plasmodium falciparum in mosquito tissues and saliva [32–35].
Transinfections in mosquitoes typically have a more severe physiological effect on their host than native infections, likely due to relatively novelty of infection, and subsequent activation of the host immune response [36]. Native infections are capable of altering host biology at the physiological and transcriptional level [25, 37], although wFlu has little apparent fitness cost [25]. They can also influence interactions between the host and pathogens, although not in the same way as transinfections. In Ae. fluviatilis, wFlu has been demonstrated to enhance infection with P. gallinaceum, increasing the intensity of infection at the oocyst stage [25]. A similar effect occurs for Culex pipiens and its native Wolbachia, wPip, during infection with Plasmodium relictum [38]. This enhancement effect has also been observed in some instances of transient Wolbachia infection in mosquitoes [39–41], where Wolbachia is injected into adult mosquitoes to produce a somatic infection, but not in all cases [42, 43]. Enhancement has only been observed rarely, and never with a stable Wolbachia transinfection for a pathogen of humans. Unless this changes, the potential impact on mosquito control programs that involve Wolbachia is likely to be limited [44, 45]. However, given the enhancement of P. gallinaceum in wFlu-infected Ae. fluviatilis, we sought to determine if a similar effect occurred for DENV.
We performed a series of experimental infections with 2 different DENV isolates in order to examine the ability of Ae. fluviatilis to harbour infection with the virus. These experiments were performed on mosquitoes with and without the native wFlu infection in order to determine if the native Wolbachia infection of Ae. fluviatilis influenced infection with the virus. We also examined the effect of wFlu on the production of reactive oxygen species (ROS), as this had been linked to pathogen interference in other Wolbachia-infected insects [46, 47].
Methods
Mosquito rearing
Two lines of Aedes fluviatilis mosquitoes were involved in these experiments—one naturally infected with the wFlu Wolbachia strain (Flu), and the other where this infection had been cleared by treatment with tetracycline (Flu.Tet), as previously described [25]. Experiments were also performed on three lines of Aedes aegypti—one infected with the wMel Wolbachia strain (Mel), and a second where the wMel infection had been removed by treatment with tetracycline (Mel.Tet), as previously described [48]. These lines were previously backcrossed to a Brazilian genetic background [48], and were regularly outcrossed with Wolbachia-uninfected mosquitoes (WT) collected from Rio de Janeiro, RJ, Brazil, thereafter, in order to maintain genetic diversity [49]. All experiments took place at least two years after tetracycline treatments had finished. Saliva injection experiments were performed using the WT line. Microbiota re-colonization was performed as previously described [25, 49].
Flu and Flu.Tet larvae were reared at a density of approximately 400 larvae in 3L of distilled water and were fed 15–20 balls of Alcon Goldfish Colour fish food per day. Mel and Mel.Tet larvae were reared at a density of 150 larvae in 3L of distilled water, and were fed a ½ tetramin tropical tablet (Tetramin) per day. Pupae were moved to large cages in groups of 500. For experimental infections, adult females were transferred to small, cylindrical cages (diameter– 16cm, height– 18cm) containing approximately 50–70 individuals. Adults were maintained on 10% sucrose, which was changed daily for virus-infected mosquitoes, and three times per week for other cages. All mosquitoes were reared in a climate-controlled insectary (temperature—27 ± 1°C, RH - 70 ± 10%, photoperiod—12 hours light: dark).
Dengue viruses & infection processes
Infections were performed using one of two DENV isolates. The DENV-2 isolate was isolated in Ribeirão Preto, SP, Brazil in 2000. The DENV-3 isolate, DENV-3 MG20 (375), was isolated in Contagem, MG, Brazil in 2013. Viruses were serially passaged in Aedes albopictus C6/36 cells, and infected supernatant harvested, titered via plaque forming assay, and then frozen at -80°C. Viral titres were 2.0x104 pfu/mL for the DENV-2 isolate, and 1.9x106 pfu/mL for the DENV-3. Virus aliquots were thawed only at the time of infection.
For injection experiments, viral stocks were injected, undiluted using a Nanoject II handheld injector (Nanoject) and glass capillaries. Each mosquito was injected intrathoracically with 69nL of virus (Approximate viral titre injected: DENV-2–1.4x100, DENV-3–1.3x102). Mosquitoes were collected at 5 days post-injection and stored at -80°C.
For oral infection experiments, viral stocks were mixed 1:1 with freshly drawn human blood, and fed to mosquitoes via glass feeders on a waterbath system. For the dilution experiment, one tube of frozen DENV-3 was thawed and diluted in L15 media, supplemented with 10% sterile, inactivated foetal bovine serum, to produce additional viral stocks at concentrations of, 1.9x104, 1.9x103 and 1.9x102 pfu/mL. These were mixed with human blood and fed, as above. Immediately post-feeding, mosquitoes were screened visually for the presence of blood in the abdomen, and non-fed mosquitoes were discarded. Mosquitoes were collected at 7 or 14 days post-infection and stored at -80°C.
Saliva collection & injection
Saliva was collected from mosquitoes that were infected with either DENV-2 or DENV-3 via injection, at 5 dpi, as previously described [34, 35]. Briefly, legs and wings were removed from anesthetized mosquitoes on ice, and mosquitoes were allowed 30 mins to expectorate into pipet tips containing 5μL of sterile, inactivated, foetal bovine serum and 30% sucrose (1:1), prepared fresh on the day of collection. These saliva samples were stored at -80°C and either assayed directly for the presence of DENV, or injected into naïve, wildtype, female Aedes aegypti mosquitoes, collected near Rio de Janeiro in 2016, as previously described [34]. Ae. aegypti mosquitoes were selected for injections given the propensity of that species to harbour infection with DENV. Each saliva was injected into 12–15 mosquitoes (volume injected: 138–276nL), which were collected at 5 dpi, and stored at -80°C. The presence or absence of DENV was determined by RT-qPCR for 7–8 injected mosquitoes per saliva sample.
DENV quantification
DENV levels in mosquitoes from the Ae. fluviatilis and Ae. aegypti oral infection experiments, the DENV-3 dilution experiment, and the DENV-2 and DENV-3 injection experiments were quantified using TaqMan-based RT-qPCR, using a previously described protocol, primers and TaqMan probe [49]. Briefly, RNA from whole mosquito samples was extracted using the TRIzol protocol (Invitrogen), first-strand cDNA synthesis was performed using the M-MLV RT protocol (Promega), and qPCR was performed using a DENV general primer set and using a Viia 7 Real-Time PCR System (ThermoFisher Scientific). Absolute DENV levels were obtained by comparison with a serially diluted (107 to 103 copies) fragment of the PCR product, previously described [49], and were normalized per 1μg of total RNA. A minimum threshold of infection of 100 copies was applied to data across all experiments, as the 102 standard did not amplify reliably.
Total RNA from saliva-injected mosquitoes was extracted using the High Pure Viral Nucleic Acid Kit (Roche), according to manufacturer’s instructions. The presence or absence of DENV was then determined via a duplex assay for DENV and the host RpS17 gene using the TaqMan Fast Virus 1-Step Master Mix (ThermoFisher Scientific), and run on a Lightcycler 96 (Roche). The qPCR mastermix contained the following components per reaction: 4.25μL water, 2μL RT-qPCR Mix, 0.5μL DENV primers (10μM), 0.1μL DENV probe (10μM—Texas Red), 0.5μL RpS17 primers (10μM), 0.1μL RpS17 probe (10μM—Hex), 0.05μL 200 X Reverse transcriptase enzyme. The water, RT-qPCR Mix, and enzyme were provided in the Fast Virus kit. The run profile was as follows: (1) 50°C for 10 mins, (2) 95°C for 30 sec, (3) 35 cycles of 95°C for 5 sec followed by 60°C for 30 sec, (4) 37°C for 30 sec.
H2O2 quantification
H2O2 quantification was performed for 6 day-old Flu and Flu.Tet adult mosquitoes using the Amplex Red Hydrogen Peroxide/Peroxidase Assay Kit (ThermoFisher Scientific cat A22188). Three types of samples were collected across two experimental replicates: (1) whole mosquito, (2) midgut, (3) fat body, with the later two obtained by dissection using a light microscope in ice cold PBS. Total samples collected ranged from 12–22 per treatment. Samples were collected on ice, and then immediately processed according to manufacturer’s instructions, as described elsewhere [50]. H2O2 levels were quantified using a microplate reader (SpectraMax M5, Molecular Devices, Sunnyvale, CA, USA), with an excitation wavelength of 530nm and an emission wavelength of 590nm, and values compared against a standard curve generated from H2O2 samples of known concentration. Mosquitoes in these assays were not infected by DENV.
Data analysis
DENV prevalence of infection was compared using Fisher’s exact test. For saliva-injected mosquitoes, prevalence of infection was compared on the individual mosquito level. Comparisons of viral load were only made for samples that tested positive for DENV. These data were all non-parametrically distributed, and were thus compared using Mann-Whitney U tests. No viral load comparisons were made for treatments with fewer than 3 positive samples, in order to satisfy the requirements of the Mann-Whitney U test. For these pairwise comparisons, only statistics for prevalence are presented. H2O2 data were all normally distributed, and were compared using unpaired t tests. All analyses were performed using Graphpad Prism V 6.0g. Figures were prepared using Graphpad Prism and Microsoft PowerPoint for Mac 2011.
Human ethics
Human blood was drawn from willing adult volunteers by trained medical personnel after obtaining written consent. This process was conducted according to established guidelines, and approved by The Committee for Ethics in Research (CEP)/ FIOCRUZ (License—CEP 732.621), and performed in accordance with the Brazilian laws (196/1996 and 01/1988), which govern human ethics issues in scientific research in compliance with the National Council of Ethics in Research (CONEP).
Results
DENV injections
We injected adult female Ae. fluviatilis and Ae. aegypti with either DENV-2 or DENV-3, collected these mosquitoes 5 days later, and then quantified levels of DENV using RT-qPCR. For Ae. fluviatilis mosquitoes injected with DENV-2 (Fig 1A), we observed that 15/22 Flu mosquitoes, and 18/22 Flu.Tet mosquitoes were positive for the virus, and that there was no effect of wFlu infection on prevalence of infection (Fisher’s exact test; P = 0.4876). Likewise, we saw no difference in viral load between Flu and Flu.Tet mosquitoes (Mann-Whitney U test; U = 89, P = 0.5045). For Ae. aegypti, we observed a decrease in DENV-2 prevalence associated with wMel infection (Fig 1B), as 8/20 Mel mosquitoes were positive for the virus compared with 12/14 Mel.Tet mosquitoes (Fisher’s exact test; P = 0.0128). Likewise, DENV-2 viral load was also decreased for Mel mosquitoes (Mann-Whitney U test; U = 16, P = 0.0283).
Fig 1. DENV-2 or DENV-3 infection via intrathoracic injection in Aedes fluviatilis and Aedes aegypti.

Prevalence of infection and DENV load for Ae. fluviatilis and Ae. aegypti mosquitoes 5 days after intrathoracic injection with DENV-2 (A&B) or DENV-3 (C&D), as determined via RT-qPCR with absolute quantification. FT (orange)—Flu.Tet. F (black)—Flu. MT (red)—Mel.Tet. M (blue)—Mel. Prevalence data analysed by Fisher’s exact test. DENV load data analysed by Mann-Whitney U test. ns = P > 0.05, * = P < 0.05, *** = P < 0.001, **** = P < 0.0001. Black lines represent treatment medians.
For Ae. fluviatilis mosquitoes challenged with DENV-3 by injection (Fig 1C), we saw 16/22 Flu and 17/22 Flu.Tet mosquitoes became infected (Fisher’s exact test; P = 1.000). However, in this case wFlu infection led to a significant decrease in DENV load (Mann-Whitney U test; U = 38, P = 0.0004). We observed no decrease in prevalence for Ae. aegypti injected with DENV-3 (Fig 1D), with 16/19 Mel mosquitoes and 17/19 Mel.Tet mosquitoes infected (Fisher’s exact test; P = 1.000). However, wMel infection did significantly reduce DENV-3 load amongst infected mosquitoes (Mann-Whitney U test; U = 21.5, P < 0.0001).
DENV oral infections
We then orally infected Flu and Flu.Tet mosquitoes with either DENV-2 or DENV-3. Infection with the DENV-2 isolate produced only a very small number of positives from either line. Accordingly, we examined data from both experimental infections together. At 7dpi (Fig 2A), we saw that 4/40 Flu and 2/42 Flu.Tet mosquitoes became infected, but this did not amount to a significant difference in prevalence (Fisher’s exact test; P = 0.6763). At 14dpi (Fig 2B), 1/32 Flu mosquitoes, and 4/39 Flu.Tet mosquitoes became infected, and again these proportions were not significantly different (Fisher’s exact test; P = 0.3806). Numbers of infected mosquitoes were not sufficient to compare viral load.
Fig 2. DENV-2 or DENV-3 oral infection in Aedes fluviatilis.

Prevalence of infection and DENV load for Ae. fluviatilis orally challenged with DENV-2 at 7 dpi (A) and 14 dpi (B), or DENV-3 at 7 dpi (C) and 14 dpi (D), as determined via RT-qPCR with absolute quantification. Data represent 2 independent infections. FT (orange)—Flu.Tet. F (black)—Flu. Prevalence data analysed by Fisher’s exact test. DENV load data analysed by Mann-Whitney U test. ns = P > 0.05. Black lines represent treatment medians.
Oral infection with the DENV-3 isolate produced a slightly higher rate of infection. Across the two experiments, we saw prevalence of infection of 15/29 for Flu mosquitoes and 13/33 for Flu.Tet mosquitoes at 7dpi (Fig 2C; Fisher’s exact test; P = 0.6503), and 11/28 for Flu and 13/25 for Flu.Tet mosquitoes at 14dpi (Fig 2D; Fisher’s exact test; P = 0.6280). Critically, there was no difference in prevalence due to Wolbachia. Likewise, we saw no difference in viral load between the two mosquito lines at either time point (Mann-Whitney U test; 7dpi—U = 71, P = 0.2352: 14dpi—U = 66, P = 0.7648).
We also examined DENV infection in Ae. aegypti, as a point of comparison. As the DENV-2 isolate did not infect Ae. fluviatilis well, we only performed experiments with the DENV-3 isolate. At 7dpi in the first experiment (Fig 3A), 27/29 Mel.Tet became infected compared to 3/30 Mel mosquitoes (Fisher’s exact test; P < 0.0001). Those Mel mosquitoes that became infected had a significantly lower viral load than Mel.Tet mosquitoes (Mann-Whitney U test; U = 4, P = 0.0054). At 14dpi (Fig 3B), 28/30 Mel.Tet were infected compared to 6/30 Mel mosquitoes (Fisher’s exact test; P <0.0001), and viral load was again significantly lower in the infected Mel mosquitoes (Mann-Whitney U test; U = 14, P = 0.0006).
Fig 3. DENV-3 oral infection in Aedes aegypti.

Prevalence of infection and DENV load for Ae. aegypti orally challenged with DENV-3 in experiment 1 (7 dpi (A) and 14 dpi (B)), and in experiment 2 (7 dpi (C) and 14 dpi (D)), as determined via RT-qPCR with absolute quantification. MT (red)—Mel.Tet. M (blue)—Mel. Prevalence data analysed by Fisher’s exact test. DENV load data analysed by Mann-Whitney U test. ** = P < 0.01, *** = P < 0.001, **** = P < 0.0001. Black lines represent treatment medians.
In the second Ae. aegypti oral infection experiment, 19/30 Mel.Tet mosquitoes and 0/30 Mel mosquitoes became infected at 7dpi (Fig 3C). Prevalence of infection was significantly lower amongst Mel mosquitoes than either Mel.Tet (Fisher’s exact test; P < 0.0001). At 14dpi (Fig 3D), 14/29 Mel.Tet mosquitoes and 1/30 Mel mosquitoes became infected. Prevalence of infection was significantly lower amongst Mel mosquitoes than Mel.Tet mosquitoes (Fisher’s exact test; P = 0.0002). As less than 3 Mel mosquitoes became infected at either time point, comparison of viral load was not performed.
In a further oral feeding assay (S1 Fig), we offered the DENV-3 isolate at dilutions of 1.9x106 (conc. 1), 1.9x104 (conc. 2), 1.9x103, and 1.9x102 pfu/mL. The prevalence of infection across the lower concentrations was less than 5% for all treatments, and the data were not considered for further analysis. For Ae. fluviatilis, 13/22 Flu mosquitoes became infected at conc. 1 at 7 dpi, compared to 3/14 Flu.Tet (Fisher’s exact test; P = 0.0407), however, we saw no effect on viral load (Mann-Whitney U test; U = 15, P = 0.6107). Six Flu and 2 Flu.Tet mosquitoes became infected at conc. 2 for the same time point, but this was not a significant difference in prevalence (Fisher’s exact test; P = 0.2127). At 14dpi, 9/18 Flu samples, and 7/17 Flu.Tet samples became infected after feeding on conc. 1 (Fisher’s exact test; P = 0.7380), with no significant difference in viral load (Mann-Whitney U test; U = 31, P > 0.999). While at conc. 2 1/18 Flu, and 2/18 Flu.Tet samples were positive for DENV-3 (Fisher’s exact test; P = 1.000). For Ae. aegypti, no Mel mosquitoes became infected at any time point, for any concentration. There was a significant decrease in prevalence associated with wMel infection at conc. 1, at both 7 and 14dpi (Fisher’s exact test; P < 0.0001). For conc. 2, Mel.tet prevalence was 0/15 and 2/11 at 7 and 14dpi, respectively.
Salivation assays
We quantified levels of DENV directly in saliva collected from Flu and Flu.Tet mosquitoes that had been injected with either the DENV-2 (Fig 4A) or DENV-3 isolates (Fig 4B). For DENV-2, 8/19 Flu saliva and 7/17 Flu.Tet saliva were positive for the virus (Fisher’s exact test; P = 1.000). While for DENV-3, 4/23 Flu saliva, and 7/28 Flu.Tet saliva were positive (Fisher’s exact test; P = 0.7338). Comparison of viral load in these saliva samples revealed that there was no significant of wFlu infection for either DENV-2 (Mann-Whitney U test; U = 27, P = 0.9551) or DENV-3 (Mann-Whitney U test; U = 9, P = 0.4121).
Fig 4. DENV-2 or DENV-3 load in the saliva of Aedes fluviatilis.

Prevalence of infection and DENV load in saliva collected from Ae. fluviatilis mosquitoes injected with DENV-2 (A) or DENV-3 (B), as determined via RT-qPCR with absolute quantification. FT (orange)—Flu.Tet. F (black)—Flu. Prevalence data analysed by Fisher’s exact test. DENV load data analysed by Mann-Whitney U test. ns = P > 0.05. Black lines represent treatment medians.
We then took 5 saliva samples collected from Flu (Fig 5A), Flu.Tet (Fig 5B), Mel (Fig 5C), and Mel.Tet (Fig 5D) mosquitoes that had been injected with DENV-3, and injected these into WT mosquitoes. For saliva from Ae. fluviatilis, 4/5 Flu and 4/5 Flu.Tet saliva produced subsequent infections, at an average infection rate of 65% and 45%, respectively. We saw no significant difference in the overall prevalence of saliva-injected mosquitoes (Fisher’s exact test; P = 0.1716). For saliva from Ae. aegypti, 2/5 Mel and 4/5 Mel.Tet saliva produced subsequent infections, at average infection rates of 20% and 47.5%, respectively. In this instance there was a decrease in overall prevalence associated with wMel infection in the mosquitoes that produced the saliva (Fisher’s exact test; P = 0.0093).
Fig 5. DENV-3 infectivity after the injection of saliva from infected Aedes fluviatilis and Aedes aegypti into naïve mosquitoes.

The presence of absence of DENV was determined in 7–8 individual mosquitoes injected with saliva collected from DENV-3-infected Flu.Tet (A), Flu (B), Mel.Tet (C), or Mel (D) mosquitoes, as determined by RT-qPCR. Each letter code (A-T) represents a single saliva sample. In each bar, black represents the proportion positive for DENV, and white represents the proportion where DENV was not detected. + = saliva that produced a subsequent infection.— = saliva that produced no subsequent infection.
Reactive oxygen species
We compared levels of ROS (H2O2) in Flu and Flu.Tet mosquitoes for the whole mosquito, midgut or fat bodies (Fig 6). We saw that H2O2 levels were not significantly different in whole mosquitoes (Unpaired t test; t = 0.8635, P = 0.3972) or in the fat body (Unpaired t test; t = 0.0514, P = 0.9594), however, H2O2 levels were slightly, but significantly elevated in the midguts of Flu mosquitoes compared to Flu.Tet (Unpaired t test; t = 2.739, P = 0.0090). Mean (± s.e.m.) H2O2 levels were 0.7563 ± 0.027 for Flu mosquitoes, and 0.6222 ± 0.041 for Flu.Tet mosquitoes.
Fig 6. H2O2 levels in Aedes fluviatilis.
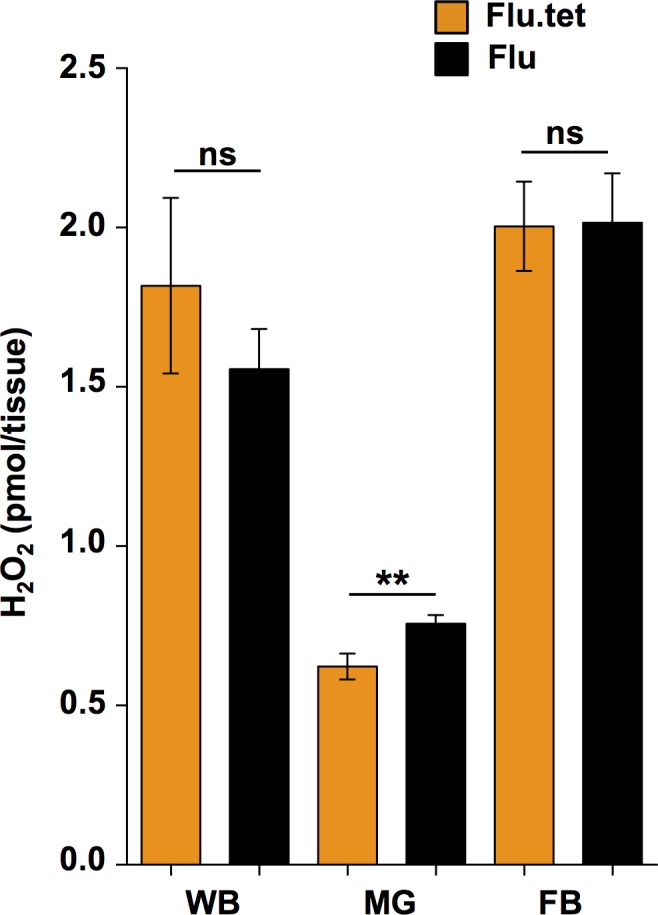
Levels of H2O2 in the whole bodies (WB), midguts (MG) and fat bodies (FB) of Flu.Tet (orange) and Flu (black) mosquitoes, quantified using the Amplex Red Hydrogen Peroxide/Peroxidase Assay Kit with a fluorometric plate reader. Data were compared using unpaired t tests. ns = P > 0.05, ** = P < 0.01.
Discussion
Our results from the experimental infection of Ae. aegypti with DENV-3 showed high prevalence of infection and DENV load in Mel.Tet mosquitoes, and a strong pathogen interference effect in Mel mosquitoes. This persisted even when the mode of infection was via injection. Several recent papers have demonstrated that wMel interferes with Zika virus and chikungunya virus in Ae. aegypti populations from Brazil and Colombia [34, 51, 52]. Our results provide further evidence that the pathogen interference phenotype operates effectively in Latin American Ae. aegypti. These results also suggested that the DENV-3 isolate that we used was sufficiently infectious to serve as a good model strain for examining DENV infection in Ae. fluviatilis. It should be noted that the our results were obtained using whole mosquito samples, and thus likely underreport the blocking effects of wMel, as this strain is a strong blocker of disseminated and salivary infection [34].
Can Aedes fluviatilis transmit DENV?
We performed experimental infections of Ae. fluviatilis mosquitoes with DENV-2 and DENV-3 isolates via oral feeding and intrathoracic injection. With the oral feeding experiments we observed very low infection rates for the DENV-2 isolate (3–10%), with this likely being due to a low viral titre (2.0x104 pfu/mL). For the DENV-3 isolate, we observed combined infection rates of 27–44% at 7dpi, and 35–37% at 14dpi for both Flu and Flu.Tet mosquitoes, across 3 experiments. These infection rates were far lower than what we observed when the Mel.Tet line (Ae. aegypti without a Wolbachia infection) was infected with the same virus, where overall infection rates were 78% at 7dpi and 71% at 14dpi, across 2 experiments. We observed higher rates of infection after Ae. fluviatilis mosquitoes were infected by intrathoracic injection, with an overall infection rate of 68–82% for DENV-2, and 73–76% for DENV-3. These values were more similar to the infection rates we observed for the Mel.Tet line (DENV-2: 86%, DENV-3: 89%). These data suggest that Ae. fluviatilis can become infected by DENV after experimental infection in the laboratory. As the titre of the DENV isolates used in our experiments was somewhat low, it is possible that Ae. fluviatilis may prove to be more susceptible to infection with different DENV isolates, or those fed at a higher titre.
We also examined the saliva of Ae. fluviatilis mosquitoes for the presence of DENV using two different techniques, after the mosquitoes had been infected with either DENV-2 or DENV-3 by injection. When viral load was quantified directly in the saliva, we observed that 41–42% of saliva tested positive for DENV-2, and 17–25% were positive for DENV-3, for Flu and Flu.Tet mosquitoes. When these saliva were injected into WT Ae. aegypti we were also able to detect DENV infection in a high percentage of mosquitoes. As virus load in these mosquitoes was quantified 5 days post-injection, this may indicate that the virus was able to replicate, suggesting that infectious vius can be found in the saliva of Ae. fluviatilis.
Our results provide some evidence that Ae. fluviatilis could potentially transmit DENV under the right circumstances. However, we state this with the acknowledgment of several caveats. Firstly, all of our data showing DENV infection in saliva were obtained after intrathoracic injection and not oral feeding. Given the lower prevalence of infection associated with oral feeding this may suggest that that the midgut is an important barrier to DENV infection in Ae. fluviatilis, and it is possible that this species does not possess some of the receptors for DENV that are present in other mosquito species [53]. Secondly, the Ae. fluviatilis lines we used are not necessarily representative of the species in the field, given that the colony has been maintained in the laboratory since 1975 [25]. It is also unclear how likely it is that Ae. fluviatilis in the field would encounter and bite a human infected with dengue, or if a DENV-infected Ae. fluviatilis would be capable of transmitting the virus to a new human host. To that end, we suggest that future research on this topic should involve a survey of wild Ae. fluviatilis populations for the presence of DENV, and that saliva be collected from further experimental infections where field populations of Ae. fluviatilis are orally infected with fresh, recently circulating DENV strains.
Influence of Wolbachia
With respect to a potential effect of wFlu on DENV infection, we observed high infection rates in Flu mosquitoes across multiple experiments, for both modes of infection. However, we saw similar results for Flu.Tet mosquitoes. In one oral feeding experiment there was increased prevalence of infection associated with wFlu, and in one injection experiment, there was decreased viral load for DENV-3 associated with wFlu. For all other experiments we saw no effect of wFlu infection on prevalence or intensity of infection. At the saliva level, there was slightly higher prevalence of infection for mosquitoes injected with saliva from DENV-infected Flu mosquitoes than Flu.Tet mosquitoes, although this difference was not significant. Taken together these results provide no evidence of a consistent effect of wFlu on DENV, which stands in contrast to what was previously observed for P. gallinaceum [25].
The ability of a Wolbachia strain to interfere with pathogens has been strongly linked to high bacterial density, and high Wolbachia density in mosquitoes is typically associated with transinfections, rather than native associations [29, 32]. In Ae. fluviatilis, wFlu is most abundant in the ovaries, which facilitates maternal transmission, but it is much less abundant in somatic tissues than transinfections, including that of wMel [25, 26, 32]. Given these observations it was not surprising that we observed no pathogen interference effect of wFlu.
ROS induction
ROS induction (higher levels of H2O2 in the presence of Wolbachia infection) in mosquitoes is a trait that is most commonly associated with Wolbachia transinfections [19, 46]. We recently demonstrated that this induction effect occurs for wMel-infected Ae. aegypti [49], while a similar effect has been observed for wAlbB-infected Ae. aegypti [46]. However, the effect has not been associated with native infections in mosquitoes. The reason for this is hypothesized to be due to a longer period of host-symbiont co-adaptation, and potentially restored redox homeostasis in native infections [54]. Our data suggest that wFlu does cause a moderate level of ROS induction in Ae. fluviatilis, but only in the midgut. Our recent transcriptomic profile of Ae. fluviatilis revealed wFlu-induced changes in the expression of several genes linked to redox process [37]. However, the fact that we observed no difference in whole mosquitoes, or in fat bodies, suggests that wFlu is unlikely to have a major impact on redox homeostasis, which could be indicative of extensive co-evolution between host and symbiont [54]. While the small scale ROS induction that we observed in the midguts was likely insufficient to have a significant effect on a pathogen.
ROS induction has also been shown to occur with some native Wolbachia infections in Drosophila, and its presence correlates well with the presence of pathogen interference [47]. But, higher ROS levels are not found in all Wolbachia-host associations where pathogen interference has been observed [55], and it is still unclear how Wolbachia-induced changes in ROS contribute to the phenotype. Not all mosquito midgut cells become infected with DENV, and there are foci of infection [56, 57]. It is unclear if the Wolbachia-induced increase in ROS is homogeneously or heterogeneously distributed through tissues and cells. If the latter were true, cells with higher ROS levels could become more resistant to the virus, leaving fewer “entry points” available.
Conclusions
We have demonstrated that the mosquito Ae. fluviatilis can become infected with DENV after intrathoracic injection or oral feeding, although at lower rates than what is observed with Ae. aegypti, a proven vector of DENV in the field. We observed that DENV can be detected in the saliva of Ae. fluviatilis after viral challenge via injection, and that this virus is likely infectious, given that it can be used to infect naïve Ae. aegypti. The vector status of this species is still uncertain, but our results suggest that it could potentially play a role in DENV transmission. Our results also indicate that the native Wolbachia infection of Ae. fluviatilis does not influence DENV infection, but does increase ROS levels in the host midgut, with neither of these observations being particularly unexpected for a native Wolbachia-host association.
Supporting information
Prevalence of infection and DENV load for Ae. fluviatilis at 7 (A) and 14dpi (B), and Ae. aegypti at 7 (C) and 14dpi (D), as determined via RT-qPCR with absolute quantification. The figure shows data for two DENV-3 concentrations that were fed to mosquitoes: 1.9 x106 pfu/mL (1), and 1.9 x 104 pfu/mL (2). FT (orange)—Flu.Tet. F (black)—Flu. MT (red)—Mel.Tet. M (blue)—Mel. Prevalence data analysed by Fisher’s exact test. DENV load data analysed by Mann-Whitney U test. ns = P > 0.05, * = P < 0.05, **** = P < 0.0001. Black lines represent treatment medians.
(TIFF)
Acknowledgments
The authors wish to thank Prof Scott L. O’Neill for providing the original wMel-infected Aedes aegypti line, Dr João Trinidade Marques, Dr Roenick Proveti Olmo, and Dr Marcele Neves Rocha for preparation or donation of dengue virus stocks, and Anna Paula de Souza Canuto for assistance with the Aedes fluviatilis colony. JBLS was a fellow from CAPES; EPC was a fellow from CNPq. LAM is a fellow from CNPq and a member of the INCT-EM, Brazil.
Data Availability
All relevant data are within the paper and its Supporting Information files.
Funding Statement
This work was supported by Conselho Nacional de Desenvolvimento Científico e Tecnológico (EPC, LAM); Fundação de Amparo à Pesquisa do Estado de Minas Gerais (EPC, LAM); Institutos Nacionais de Ciência e Tecnologia- Entomologia Molecular (ICNT-EM) (LAM). The funders had no role in study design, data collection and analysis, decision to publish, or preparation of the manuscript.
References
- 1.Bhatt S, Gething PW, Brady OJ, Messina JP, Farlow AW, Moyes CL, et al. The global distribution and burden of dengue. Nature. 2013;496(7446):504–7. doi: 10.1038/nature12060 ; PubMed Central PMCID: PMCPMC3651993. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 2.Brady OJ, Gething PW, Bhatt S, Messina JP, Brownstein JS, Hoen AG, et al. Refining the global spatial limits of dengue virus transmission by evidence-based consensus. PLoS Negl Trop Dis. 2012;6(8):e1760 doi: 10.1371/journal.pntd.0001760 ; PubMed Central PMCID: PMCPMC3413714. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 3.Kyle JL, Harris E. Global spread and persistence of dengue. Annu Rev Microbiol. 2008;62:71–92. doi: 10.1146/annurev.micro.62.081307.163005 . [DOI] [PubMed] [Google Scholar]
- 4.Kraemer MU, Sinka ME, Duda KA, Mylne AQ, Shearer FM, Barker CM, et al. The global distribution of the arbovirus vectors Aedes aegypti and Ae. albopictus. Elife. 2015;4:e08347 doi: 10.7554/eLife.08347 ; PubMed Central PMCID: PMCPMC4493616. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 5.Campbell LP, Luther C, Moo-Llanes D, Ramsey JM, Danis-Lozano R, Peterson AT. Climate change influences on global distributions of dengue and chikungunya virus vectors. Philos Trans R Soc Lond B Biol Sci. 2015;370(1665). doi: 10.1098/rstb.2014.0135 ; PubMed Central PMCID: PMCPMC4342968. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 6.Dutra HL, Caragata EP, Moreira LA. The re-emerging arboviral threat: Hidden enemies: The emergence of obscure arboviral diseases, and the potential use of Wolbachia in their control. Bioessays. 2017;39(2). doi: 10.1002/bies.201600175 . [DOI] [PubMed] [Google Scholar]
- 7.Lambrechts L, Scott TW, Gubler DJ. Consequences of the expanding global distribution of Aedes albopictus for dengue virus transmission. PLoS Negl Trop Dis. 2010;4(5):e646 doi: 10.1371/journal.pntd.0000646 ; PubMed Central PMCID: PMCPMC2876112. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 8.Wilder-Smith A, Gubler DJ. Geographic expansion of dengue: the impact of international travel. Med Clin North Am. 2008;92(6):1377–90, x. doi: 10.1016/j.mcna.2008.07.002 . [DOI] [PubMed] [Google Scholar]
- 9.Ranson H, Lissenden N. Insecticide Resistance in African Anopheles Mosquitoes: A Worsening Situation that Needs Urgent Action to Maintain Malaria Control. Trends Parasitol. 2016;32(3):187–96. doi: 10.1016/j.pt.2015.11.010 . [DOI] [PubMed] [Google Scholar]
- 10.Smith LB, Kasai S, Scott JG. Pyrethroid resistance in Aedes aegypti and Aedes albopictus: Important mosquito vectors of human diseases. Pestic Biochem Physiol. 2016;133:1–12. doi: 10.1016/j.pestbp.2016.03.005 . [DOI] [PubMed] [Google Scholar]
- 11.Auguste AJ, Adams AP, Arrigo NC, Martinez R, Travassos da Rosa AP, Adesiyun AA, et al. Isolation and characterization of sylvatic mosquito-borne viruses in Trinidad: enzootic transmission and a new potential vector of Mucambo virus. Am J Trop Med Hyg. 2010;83(6):1262–5. doi: 10.4269/ajtmh.2010.10-0280 ; PubMed Central PMCID: PMCPMC2990042. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 12.de Figueiredo ML, de C. Gomes A, Amarilla AA, de S Leandro A, de S Orrico A, de Araujo RF, et al. Mosquitoes infected with dengue viruses in Brazil. Virol J. 2010;7:152 doi: 10.1186/1743-422X-7-152 ; PubMed Central PMCID: PMCPMC2913956. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 13.Consoli RA, Williams P, Moreno EC, Santos BS. Features of reproduction in laboratory-reared Aedes fluviatilis (Lutz, 1904) (Diptera: Culicidae). Mem Inst Oswaldo Cruz. 1983;78(1):37–47. . [DOI] [PubMed] [Google Scholar]
- 14.Davis N, Shannon R. Studies on yellow fever in South America: attempts to transmit the virus with certain Aedine and Sabethine mosquitoes and with Triatomas (Hemiptera). Am J Trop Med Hyg. 1931;s1-11(1):21–9. doi: 10.4269/ajtmh.1931.s1-11.21 [Google Scholar]
- 15.de Camargo MV, Consoli RA, Williams P, Krettli AU. Factors influencing the development of Plasmodium gallinaceum in Aedes fluviatilis. Mem Inst Oswaldo Cruz. 1983;78(1):83–94. . [DOI] [PubMed] [Google Scholar]
- 16.Zug R, Hammerstein P. Still a host of hosts for Wolbachia: analysis of recent data suggests that 40% of terrestrial arthropod species are infected. PLoS One. 2012;7(6):e38544 doi: 10.1371/journal.pone.0038544 ; PubMed Central PMCID: PMCPMC3369835. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 17.Augustinos AA, Santos-Garcia D, Dionyssopoulou E, Moreira M, Papapanagiotou A, Scarvelakis M, et al. Detection and characterization of Wolbachia infections in natural populations of aphids: is the hidden diversity fully unraveled? PLoS One. 2011;6(12):e28695 doi: 10.1371/journal.pone.0028695 ; PubMed Central PMCID: PMCPMC3236762. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 18.Bordenstein SR, Paraskevopoulos C, Dunning Hotopp JC, Sapountzis P, Lo N, Bandi C, et al. Parasitism and mutualism in Wolbachia: what the phylogenomic trees can and cannot say. Mol Biol Evol. 2009;26(1):231–41. doi: 10.1093/molbev/msn243 ; PubMed Central PMCID: PMCPMC2721558. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 19.Caragata EP, Dutra HL, Moreira LA. Exploiting intimate relationships: Controlling mosquito-transmitted disease with Wolbachia. Trends in Parasitology. 2016. doi: doi.org/10.1016/j.pt.2015.10.011 [DOI] [PubMed] [Google Scholar]
- 20.Werren JH, Baldo L, Clark ME. Wolbachia: master manipulators of invertebrate biology. Nat Rev Microbiol. 2008;6(10):741–51. doi: 10.1038/nrmicro1969 . [DOI] [PubMed] [Google Scholar]
- 21.Bouchon D, Rigaud T, Juchault P. Evidence for widespread Wolbachia infection in isopod crustaceans: molecular identification and host feminization. Proc Biol Sci. 1998;265(1401):1081–90. doi: 10.1098/rspb.1998.0402 ; PubMed Central PMCID: PMCPMC1689171. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 22.Fialho RF, Stevens L. Male-killing Wolbachia in a flour beetle. Proc Biol Sci. 2000;267(1451):1469–73. doi: 10.1098/rspb.2000.1166 ; PubMed Central PMCID: PMCPMC1690698. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 23.Turelli M, Hoffmann AA. Rapid spread of an inherited incompatibility factor in California Drosophila. Nature. 1991;353(6343):440–2. doi: 10.1038/353440a0 . [DOI] [PubMed] [Google Scholar]
- 24.Zchori-Fein E, Faktor O, Zeidan M, Gottlieb Y, Czosnek H, Rosen D. Parthenogenesis-inducing microorganisms in Aphytis (Hymenoptera: Aphelinidae). Insect Mol Biol. 1995;4(3):173–8. . [DOI] [PubMed] [Google Scholar]
- 25.Baton LA, Pacidonio EC, Goncalves DS, Moreira LA. wFlu: characterization and evaluation of a native Wolbachia from the mosquito Aedes fluviatilis as a potential vector control agent. PLoS One. 2013;8(3):e59619 doi: 10.1371/journal.pone.0059619 ; PubMed Central PMCID: PMCPMC3608659. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 26.Moreira LA, Iturbe-Ormaetxe I, Jeffery JA, Lu GJ, Pyke AT, Hedges LM, et al. A Wolbachia symbiont in Aedes aegypti limits infection with Dengue, Chikungunya, and Plasmodium. Cell. 2009;139(7):1268–78. doi: 10.1016/j.cell.2009.11.042 ISI:000273048700015. [DOI] [PubMed] [Google Scholar]
- 27.Coon KL, Brown MR, Strand MR. Mosquitoes host communities of bacteria that are essential for development but vary greatly between local habitats. Mol Ecol. 2016;25(22):5806–26. doi: 10.1111/mec.13877 ; PubMed Central PMCID: PMCPMC5118126. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 28.Hughes GL, Rasgon JL. Transinfection: a method to investigate Wolbachia-host interactions and control arthropod-borne disease. Insect Mol Biol. 2014;23(2):141–51. doi: 10.1111/imb.12066 ; PubMed Central PMCID: PMCPMC3949162. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 29.Joubert DA, Walker T, Carrington LB, De Bruyne JT, Kien DH, Hoang Nle T, et al. Establishment of a Wolbachia Superinfection in Aedes aegypti Mosquitoes as a Potential Approach for Future Resistance Management. PLoS Pathog. 2016;12(2):e1005434 doi: 10.1371/journal.ppat.1005434 ; PubMed Central PMCID: PMCPMC4758728. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 30.McMeniman CJ, Lane RV, Cass BN, Fong AWC, Sidhu M, Wang YF, et al. Stable introduction of a life-shortening Wolbachia infection into the mosquito Aedes aegypti. Science. 2009;323(5910):141–4. doi: 10.1126/science.1165326 ISI:000262104100061. [DOI] [PubMed] [Google Scholar]
- 31.Xi Z, Khoo CC, Dobson SL. Wolbachia establishment and invasion in an Aedes aegypti laboratory population. Science. 2005;310(5746):326–8. Epub 2005/10/15. doi: 10.1126/science.1117607 . [DOI] [PubMed] [Google Scholar]
- 32.Walker T, Johnson PH, Moreira LA, Iturbe-Ormaetxe I, Frentiu FD, McMeniman CJ, et al. The wMel Wolbachia strain blocks dengue and invades caged Aedes aegypti populations. Nature. 2011;476(7361):450–3. doi: 10.1038/nature10355 . [DOI] [PubMed] [Google Scholar]
- 33.Bian G, Joshi D, Dong Y, Lu P, Zhou G, Pan X, et al. Wolbachia invades Anopheles stephensi populations and induces refractoriness to Plasmodium infection. Science. 2013;340(6133):748–51. doi: 10.1126/science.1236192 . [DOI] [PubMed] [Google Scholar]
- 34.Dutra HL, Rocha MN, Dias FB, Mansur SB, Caragata EP, Moreira LA. Wolbachia Blocks Currently Circulating Zika Virus Isolates in Brazilian Aedes aegypti Mosquitoes. Cell Host Microbe. 2016;19(6):771–4. doi: 10.1016/j.chom.2016.04.021 . [DOI] [PMC free article] [PubMed] [Google Scholar]
- 35.Ferguson NM, Kien DT, Clapham H, Aguas R, Trung VT, Chau TN, et al. Modeling the impact on virus transmission of Wolbachia-mediated blocking of dengue virus infection of Aedes aegypti. Sci Transl Med. 2015;7(279):279ra37 doi: 10.1126/scitranslmed.3010370 ; PubMed Central PMCID: PMCPMC4390297. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 36.Rancès E, Ye YH, Woolfit M, McGraw EA, O'Neill SL. The relative importance of innate immune priming in Wolbachia-mediated Dengue interference. Plos Pathogens. 2012;8(2). doi: e100254810.1371/journal.ppat.1002548 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 37.Caragata EP, Pais FS, Baton LA, Silva JB, Sorgine MH, Moreira LA. The transcriptome of the mosquito Aedes fluviatilis (Diptera: Culicidae), and transcriptional changes associated with its native Wolbachia infection. BMC Genomics. 2017;18(1):6 doi: 10.1186/s12864-016-3441-4 ; PubMed Central PMCID: PMCPMC5210266. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 38.Zélé F, Nicot A, Berthomieu A, Weill M, Duron O, Rivero A. Wolbachia increases susceptibility to Plasmodium infection in a natural system. Proc Biol Sci. 2014;281(1779):20132837 doi: 10.1098/rspb.2013.2837 ; PubMed Central PMCID: PMCPMC3924077. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 39.Dodson BL, Hughes GL, Paul O, Matacchiero AC, Kramer LD, Rasgon JL. Wolbachia enhances West Nile virus (WNV) infection in the mosquito Culex tarsalis. PLoS Negl Trop Dis. 2014;8(7):e2965 doi: 10.1371/journal.pntd.0002965 ; PubMed Central PMCID: PMCPMC4091933. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 40.Hughes GL, Vega-Rodriguez J, Xue P, Rasgon JL. Wolbachia strain wAlbB enhances infection by the rodent malaria parasite Plasmodium berghei in Anopheles gambiae mosquitoes. Appl Environ Microbiol. 2012;78(5):1491–5. doi: 10.1128/AEM.06751-11 ; PubMed Central PMCID: PMCPMC3294472. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 41.Murdock CC, Blanford S, Hughes GL, Rasgon JL, Thomas MB. Temperature alters Plasmodium blocking by Wolbachia. Sci Rep. 2014;4:3932 doi: 10.1038/srep03932 ; PubMed Central PMCID: PMCPMC3909897. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 42.Hughes GL, Koga R, Xue P, Fukatsu T, Rasgon JL. Wolbachia infections are virulent and inhibit the human malaria parasite Plasmodium falciparum in Anopheles gambiae. PLoS Pathog. 2011;7(5):e1002043 doi: 10.1371/journal.ppat.1002043 ; PubMed Central PMCID: PMCPMC3098226. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 43.Joubert DA, O'Neill SL. Comparison of Stable and Transient Wolbachia Infection Models in Aedes aegypti to Block Dengue and West Nile Viruses. PLoS Negl Trop Dis. 2017;11(1):e0005275 doi: 10.1371/journal.pntd.0005275 ; PubMed Central PMCID: PMCPMC5241016. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 44.Hoffmann AA, Montgomery BL, Popovici J, Iturbe-Ormaetxe I, Johnson PH, Muzzi F, et al. Successful establishment of Wolbachia in Aedes populations to suppress dengue transmission. Nature. 2011;476(7361):454–U107. doi: 10.1038/nature10356 [DOI] [PubMed] [Google Scholar]
- 45.Mains JW, Brelsfoard CL, Rose RI, Dobson SL. Female Adult Aedes albopictus Suppression by Wolbachia-Infected Male Mosquitoes. Sci Rep. 2016;6:33846 doi: 10.1038/srep33846 . [DOI] [PMC free article] [PubMed] [Google Scholar]
- 46.Pan X, Zhou G, Wu J, Bian G, Lu P, Raikhel AS, et al. Wolbachia induces reactive oxygen species (ROS)-dependent activation of the Toll pathway to control dengue virus in the mosquito Aedes aegypti. Proceedings of the National Academy of Sciences of the United States of America. 2012;109(1):E23–E31. doi: 10.1073/pnas.1116932108 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 47.Wong ZS, Brownlie JC, Johnson KN. Oxidative stress correlates with Wolbachia-mediated antiviral protection in Wolbachia-Drosophila associations. Appl Environ Microbiol. 2015;81(9):3001–5. doi: 10.1128/AEM.03847-14 ; PubMed Central PMCID: PMCPMC4393424. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 48.Dutra HL, Dos Santos LM, Caragata EP, Silva JB, Villela DA, Maciel-de-Freitas R, et al. From lab to field: the influence of urban landscapes on the invasive potential of Wolbachia in Brazilian Aedes aegypti mosquitoes. PLoS Negl Trop Dis. 2015;9(4):e0003689 doi: 10.1371/journal.pntd.0003689 ; PubMed Central PMCID: PMCPMC4408005. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 49.Caragata EP, Rezende FO, Simões TS, Moreira LA. Diet-induced nutritional stress and pathogen interference in Wolbachia-infected Aedes aegypti. PLoS Negl Trop Dis. 2016;10(11):e0005158 Epub November 28 2016. doi: 10.1371/journal.pntd.0005158 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 50.Oliveira JHM, Talyuli OAC, Goncalves RLS, Paiva-Silva GO, Sorgine MHF, Alvarenga PH, et al. Catalase protects Aedes aegypti from oxidative stress and increases midgut infection prevalence of Dengue but not Zika. PLoS Negl Trop Dis. 2017;11(4):e0005525 doi: 10.1371/journal.pntd.0005525 . [DOI] [PMC free article] [PubMed] [Google Scholar]
- 51.Aliota MT, Peinado SA, Velez ID, Osorio JE. The wMel strain of Wolbachia Reduces Transmission of Zika virus by Aedes aegypti. Sci Rep. 2016;6:28792 doi: 10.1038/srep28792 ; PubMed Central PMCID: PMCPMC4929456. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 52.Aliota MT, Walker EC, Uribe Yepes A, Velez ID, Christensen BM, Osorio JE. The wMel Strain of Wolbachia Reduces Transmission of Chikungunya Virus in Aedes aegypti. PLoS Negl Trop Dis. 2016;10(4):e0004677 doi: 10.1371/journal.pntd.0004677 ; PubMed Central PMCID: PMCPMC4849757. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 53.Cruz-Oliveira C, Freire JM, Conceicao TM, Higa LM, Castanho MA, Da Poian AT. Receptors and routes of dengue virus entry into the host cells. FEMS Microbiol Rev. 2015;39(2):155–70. doi: 10.1093/femsre/fuu004 . [DOI] [PubMed] [Google Scholar]
- 54.Zug R, Hammerstein P. Wolbachia and the insect immune system: what reactive oxygen species can tell us about the mechanisms of Wolbachia-host interactions. Front Microbiol. 2015;6:1201 doi: 10.3389/fmicb.2015.01201 ; PubMed Central PMCID: PMCPMC4621438. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 55.Molloy JC, Sinkins SP. Wolbachia Do Not Induce Reactive Oxygen Species-Dependent Immune Pathway Activation in Aedes albopictus. Viruses. 2015;7(8):4624–39. doi: 10.3390/v7082836 ; PubMed Central PMCID: PMCPMC4576197. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 56.Olson KE, Myles KM, Seabaugh RC, Higgs S, Carlson JO, Beaty BJ. Development of a Sindbis virus expression system that efficiently expresses green fluorescent protein in midguts of Aedes aegypti following per os infection. Insect Mol Biol. 2000;9(1):57–65. . [DOI] [PubMed] [Google Scholar]
- 57.Sanchez-Vargas I, Scott JC, Poole-Smith BK, Franz AW, Barbosa-Solomieu V, Wilusz J, et al. Dengue virus type 2 infections of Aedes aegypti are modulated by the mosquito's RNA interference pathway. PLoS Pathog. 2009;5(2):e1000299 doi: 10.1371/journal.ppat.1000299 ; PubMed Central PMCID: PMCPMC2633610. [DOI] [PMC free article] [PubMed] [Google Scholar]
Associated Data
This section collects any data citations, data availability statements, or supplementary materials included in this article.
Supplementary Materials
Prevalence of infection and DENV load for Ae. fluviatilis at 7 (A) and 14dpi (B), and Ae. aegypti at 7 (C) and 14dpi (D), as determined via RT-qPCR with absolute quantification. The figure shows data for two DENV-3 concentrations that were fed to mosquitoes: 1.9 x106 pfu/mL (1), and 1.9 x 104 pfu/mL (2). FT (orange)—Flu.Tet. F (black)—Flu. MT (red)—Mel.Tet. M (blue)—Mel. Prevalence data analysed by Fisher’s exact test. DENV load data analysed by Mann-Whitney U test. ns = P > 0.05, * = P < 0.05, **** = P < 0.0001. Black lines represent treatment medians.
(TIFF)
Data Availability Statement
All relevant data are within the paper and its Supporting Information files.