

Abstract
The cochlea uses active amplification to capture faint sounds. It has been proposed that the amplifier comprises a set of self-tuned critical oscillators: each hair cell contains a force-generating dynamical system that is maintained at the threshold of an oscillatory instability, or Hopf bifurcation. While the active response to a pure tone provides frequency selectivity, exquisite sensitivity, and wide dynamic range, its intrinsic nonlinearity causes tones of different frequency to interfere with one another in the cochlea. Here we determine the response to two tones, which provides a framework for understanding how the ear processes the more complex sounds of speech and music. Our calculations of two-tone suppression and the spectrum of distortion products generated by a critical oscillator accord with experimental observations of basilar membrane motion and the nervous response. We discuss how the response of a set of self-tuned oscillators, covering a range of characteristic frequencies, represents the structure of a complex sound. The frequency components of the stimulus can be inferred from the timing of neural spikes elicited by the vibrating hair cells. Passive prefiltering by the basilar membrane improves pitch discrimination by reducing interference between tones. Our analysis provides a general framework for examining the relation between the physical nature of the peripheral detection apparatus and psychophysical phenomena such as the sensation of dissonance and auditory illusions.
The nonlinear nature of sound detection has been known for more than 250 years, ever since Tartini described the perception of combination tones that are not present in a complex sound stimulus (1). More recently, experimental techniques have made it possible directly to observe nonlinearities in the basilar membrane motion (2–8) and in the signals of the auditory nerve (9) and, in some instances, to trace them to the transduction process in the hair cells of the inner ear (10, 11). Nonlinearities imply that the response to two tones is not simply a superposition of single frequency responses—different frequencies interfere with and distort one another (10). Manifestations of this interference, which have been identified experimentally, include two-tone suppression (7–9) and the generation of distortion product (DP) frequencies (5, 6). Here we demonstrate that these physiological observations are a direct consequence of the active system of signal detection (12), which the ear uses to amplify weak signals. This insight permits us to draw conclusions about the way that the auditory system infers information about pitch and to shed light on a variety of psychophysical observations.
Active Amplification
That an active mechanism operates in hearing was predicted as long ago as 1948 by Gold (13), who argued that the cochlea might work analogously to a regenerative radio receiver and use a source of energy to counteract the limiting effects of friction and actively amplify the stimulus. General acceptance of this viewpoint has been hampered by the lack of a quantitative theory to confront with experimental data and also by doubts about how the hypothesized amplificatory feedback could be correctly regulated. The recently introduced notion of self-tuned criticality (14) addresses these issues. According to this concept, each hair cell contains a force-generating dynamical system that is poised on the verge of an oscillatory instability (a Hopf bifurcation) and is kept at that critical point by a self-adjustment mechanism. Such a critical oscillator is especially responsive to weak sinusoidal stimuli applied at its characteristic frequency, and calculations have demonstrated that a Hopf resonance provides frequency selectivity, extreme sensitivity, and a broad dynamic range as a result of nonlinear amplification (14, 15).
Here, we recall the response of a mechanical amplifier, which is self-tuned to a Hopf bifurcation, to a pure tone of frequency f, characterized by the amplitude Ff of a periodic stimulus force. The system responds with a deflection X, which is dominated by the Fourier amplitude Xf at the same frequency. Stimulus and response are related by an expansion of the form (14)
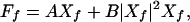 |
1 |
where the principal nonlinearity is cubic. The complex coefficients A(f) and B(f) are frequency-dependent. The Hopf bifurcation is characterized by the fact that A vanishes for a characteristic frequency fc:
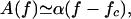 |
2 |
with a complex coefficient α. If the stimulus frequency is close to the characteristic frequency, the linear term in Eq. 1 is insignificant and the system displays a nonlinear amplified response, Xf ∼ F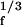 . This occurs when |f − fc| < Δfa, where Δfa ≡ |7B1/3F
. This occurs when |f − fc| < Δfa, where Δfa ≡ |7B1/3F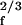 /(4α)| denotes the bandwidth of active amplification, which depends on the stimulus amplitude. For frequencies outside this bandwidth, the response is linear, Xf ∼ Ff/|f − fc|. Within the sensitive bandwidth Δfa the system amplifies with a gain |Xf|/|Ff| ∼ F
/(4α)| denotes the bandwidth of active amplification, which depends on the stimulus amplitude. For frequencies outside this bandwidth, the response is linear, Xf ∼ Ff/|f − fc|. Within the sensitive bandwidth Δfa the system amplifies with a gain |Xf|/|Ff| ∼ F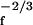 , which becomes very high for weak stimuli. The power-law response of the system allows it to operate over a wide dynamic range; it compresses the 12 orders of magnitude in stimulus intensity that the ear can hear into deflections that vary by only a factor of 100.
, which becomes very high for weak stimuli. The power-law response of the system allows it to operate over a wide dynamic range; it compresses the 12 orders of magnitude in stimulus intensity that the ear can hear into deflections that vary by only a factor of 100.
Two-Tone Interference
Two-tone suppression and the generation of DPs can be explained by the two-tone interferences generated at a Hopf bifurcation. Because the response of a Hopf oscillator is generic near its critical point, we can describe the main features of these effects without detailed knowledge of the underlying molecular mechanisms.
We are interested in the response to two tones (frequencies f1 and f2, frequency difference Δf = |f2 − f1|) with amplitudes Ff1 and Ff2 acting on a critical Hopf oscillator with fc = f1. The spectrum of the response contains the corresponding amplitudes Xf1 and Xf2. Both amplitudes, however, are systematically smaller than each of them would be separately in the absence of the second tone, as is observed experimentally (7). This two-tone suppression is illustrated in Fig. 1, which displays the numerical solutions of a simple model for a Hopf bifurcation (see Appendix A). The diagram shows that the nonlinear amplification of a tone at the oscillator's characteristic frequency can be extinguished by the presence of the second tone, especially when Ff2 > Ff1 and Δf < Δfa. Two-tone suppression near a Hopf bifurcation is generic and follows from nonlinearities in the expansion of the Fourier modes (see Appendix B). Analysis reveals that the presence of Xf2 in the response spectrum generates an effective linear term in the equation for Xf1. This mode thus behaves as if the oscillator were not tuned precisely to the bifurcation point. The corresponding loss of amplification would lead to an increased detection threshold when a second tone (or noise) is introduced. This phenomenon is referred to as masking (16).
Figure 1.

Two-tone suppression at a Hopf bifurcation. A Hopf oscillator of characteristic frequency fc is stimulated by an oscillating force that contains two frequency components: a fixed frequency f1 = fc and a variable frequency f2. The amplitudes of the stimulus force at frequencies f1 and f2 are denoted Ff1 and Ff2, respectively. (a) Response of the system for the case of equal stimulus amplitudes Ff1 = Ff2 as a function of the frequency difference Δf = f1 − f2. The bandwidth of nonlinear amplification of the Hopf oscillator is denoted Δfa. Displayed are the Fourier amplitudes of the displacement at the two stimulus frequencies, Xf1 and Xf2. As f2 approaches f1, the response Xf1 is suppressed. Similarly, the response Xf2 is suppressed as compared to the response of the system to a single tone at frequency f2, shown as a faint line for comparison. For Δf = 0 (i.e. f1 = f2), only a single frequency is present; the Fourier amplitude of the response at this frequency is shown as an individual point at the top in the center. (b) Displacement amplitudes Xf1 and Xf2 of the response to a two-frequency stimulus with a small relative frequency difference (Δf/Δfa = 0.05) as a function of the stimulus magnitude Ff1, for fixed Ff2. The response Xf2 is increasingly suppressed as Ff1 is raised. The response Xf1 is also diminished in comparison to the single-tone response (i.e. Ff2 = 0), which is displayed as a faint red line for comparison. The suppression of Xf1 is negligible when Ff1 ≫ Ff2; similarly, the suppression of Xf2 is negligible when Ff1 ≫ Ff2.
The nonlinearities of a Hopf bifurcation also generate new frequencies. The amplitudes Xf1 and Xf2 couple to the amplitudes with frequencies 2f1 − f2 and 2f2 − f1, which are therefore also present in the response (see Appendix B). They subsequently excite further DPs. This leads to a hierarchy of DPs with frequencies fk ≡ f1 + (k − 1)Δf, where k is a positive or negative integer. For large |k − 3/2| > Δfa/Δf the amplitudes decrease exponentially
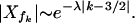 |
3 |
This characteristic spectrum of DPs is apparent in Fig. 2, where the response of a simple model (see Appendix A) is displayed for three different values of Δf, together with the corresponding waveforms. The coefficient λ−1, characterizing the number of strongly excited DPs, decreases as Δf increases. These findings are consistent with experimental data (6). For Δf < Δfa, a large number of modes is excited and deviations from the exponential law appear. In the limit of vanishing Δf, a singular limit is attained for which the DP amplitudes decay as a power law |Xfk| ∼ |k − 3/2|−v, with v = 4/3 (see Appendix B). This is confirmed by numerical solutions of the simple model described in Appendix A, for which we find v ≃ 1.31 ± 0.05.
Figure 2.

Two-tone distortion products at a Hopf bifurcation. A Hopf oscillator of characteristic frequency fc is stimulated by an oscillating force that contains two frequency components, f1 = fc and f2, of equal magnitude. (a) Spectral representation of the amplitude Xf of the response as a function of frequency f for several values of the frequency difference Δf = f1 − f2 (Δf/fc = 0.2, 0.05 and 0.01). The response exhibits distinct spikes corresponding to distortion products of frequencies fk = f1 + (k − 1)Δf, where k is an integer. The examples shown correspond to a high-level stimulus (see Appendix A). (b) Complex waveforms of the response X as a function of time t that correspond to the spectra displayed in a.
It has previously been suggested that two-tone suppression might be explained by a passive nonlinearity sandwiched between two linear bandpass filters (17). Such a system also could produce a prominent DP at frequency 2f1 − f2 on the assumption that the nonlinearity is cubic (17). This picture has no physical foundation, however, and also fails to account for the observed amplitude dependence of the DP spectrum. By contrast, a self-tuned Hopf bifurcation simultaneously describes two-tone suppression and DP generation, as well as explaining why the nonlinear response to a single tone at the characteristic frequency is cubic (14). Moreover, the model has a sound physical basis, because the presence of an active system of amplification is well-established (12).
Passive Prefilter
Although a Hopf bifurcation provides excellent amplification, especially for weak signals, its ability to filter frequencies is less impressive. An oscillator with fc = f1 can have a significant contribution Xf2 in its response spectrum, even when Δf > Δfa. This interference, which would pose problems for the detection of complex sounds, can be reduced by prefiltering the stimulus. It is well-established that the mechanical properties of the basilar membrane provide such a filter (19), which, owing to the tonotopic organization of hair cells (20), is centered on the characteristic frequency of each oscillator. The bandwidth Δfp of this passive prefilter sets a frequency interval above which two-tone interference is suppressed, whatever the level of the stimulus. This bandwidth is therefore roughly equal to the critical bandwidth ΔfCB (16) measured at moderate to high intensities, for which the active system does not significantly sharpen the tuning. At low intensities, Δfa ≪ Δfp and the active amplifier is itself very effective at suppressing interference; in this case we expect the critical bandwidth to diminish, as is indeed observed experimentally (18).
Pitch Extraction
Our analysis contributes to the longstanding debate about whether frequency is encoded by place or by timing. An example of the response of a full set of hair cells, from which frequency information must be derived, is shown in Fig. 3. Could frequency be represented by the spatial distribution of the neural response, which is maximal where the disturbance is greatest? This notion (which was originally promoted by Békésy's experiments on cochlear mechanics) suffers a drawback. The passive filter is too broad to account for the observed pitch discrimination (19) and, although the active amplifier certainly sharpens the tuning for weak stimuli, it has little effect at high intensities. Many have argued that the majority of information about frequency is derived from the detailed time course of the response of the hair cells, via the timing of neural spikes (21, 22). When stimulated by two tones, each excited oscillator vibrates in a pattern that contains components at f1 and f2 and also at the DP frequencies. Both the absolute and the relative sizes of each component vary along the cochlea, as the characteristic frequency of the hair cells changes (Fig. 3). The nervous system is provided with only partial information about each of these complex waveforms, in the form of a time series of nervous spikes. We argue below that clear identification of a frequency component is possible only if it is dominant in the motion. This suggests a role for basilar membrane resonance in contraposition to place coding. By prefiltering the stimulus and limiting interference, it permits accurate inference of frequency from nervous timing.
Figure 3.

Two-tone response of a set of Hopf oscillators. The oscillators cover a range of critical frequencies fc. They are forced by an oscillating stimulus that contains two frequency components f1 and f2 (f1/f2 = 1.05). Both components have an equal amplitude of moderate level (see Appendix A). Shown are the Fourier amplitudes Xf of the displacement at the two stimulus frequencies and at the frequencies of the principal distortion products (2f1 − f2, 2f2 − f1, 3f1 − 2f2 and 3f2 − 2f1), as a function of the critical frequency fc of the oscillators. In the cochlea, the critical frequency varies with position. The diagram can thus be interpreted as the predicted cochlear response along the basilar membrane. Over a wide range of fc, the excited oscillators respond with three significant, but unequal, frequency components and therefore display complex, nonlinear waveforms.
We base our model of pitch discrimination on the generic nervous response of hair cells: (i) spikes are elicited whenever the hair bundle deflection traverses a threshold (for frequencies below 5 kHz, at least); and (ii) hair cells with a range of different thresholds are present at each characteristic frequency (23). How might the nervous system extract information about frequency and intensity from the resulting spike trains? A simple, but effective, algorithm is to compute a histogram of inter-spike intervals T, summing over all the hair cells in the cochlea (and integrating over a fraction of a second). Perceived tones correspond to peaks in the histogram and are assigned pitch 1/T, while the perceived loudness of a tone is related to the height of the peak. Examples of this procedure are shown in Fig. 4a. Three regimes are apparent, depending on the ratio of the stimulus frequencies. (i) When the two frequencies are very close, Δf ≪ Δfp, a single tone is perceived with pitch (f1 + f2)/2. In addition, there are strong loudness fluctuations at the beat frequency f2 − f1. The quality of the beats depends on Δf; the ratio of silence to loudness diminishes as the frequencies approach one another (reflecting the waveform in Fig. 2b). (ii) When the two frequencies differ by a few percent, Δf < Δfp, the perceived pitch is more ambiguous. The histogram becomes much broader, making pitch assignment less accurate. Furthermore, in the case where two pitches can be discriminated, both undergo rapid fluctuations in loudness with different phases. The resulting variability of the perceived pitch would account for the roughness of sensation that is experienced in this situation (24). (iii) At larger frequency differences, Δf > Δfp, the two pitches f1 and f2 are accurately and clearly distinguished. Although we do not rule out the possibility that the nervous system uses a more sophisticated algorithm to infer pitch, the results summarized in Fig. 4b concur with a wide variety of psychophysical observations (16).
Figure 4.

Pitch determination in the presence of a two-tone stimulus. The response of a Hopf oscillator can be used to generate a model nervous spike train by assuming that a spike is elicited whenever the displacement amplitude X of the oscillator traverses a threshold value. The time intervals T between consecutive spikes provide information about the frequency components in the stimulus. (a) Pitch extraction. Histogram of the reciprocal, T−1, of the inter-spike intervals for three different values of f2/f1, where f1 and f2 are the frequency components in the stimulus (marked by red arrows). The histogram is summed over a number of oscillators with different characteristic frequencies and a range of thresholds (see Appendix A). For f1/f2 = 1.3, two pitches corresponding to the stimulus frequencies can be clearly identified. For f1/f2 = 1.1, two pitches can be distinguished, but the corresponding peaks are broadened and distorted, suggesting greater ambiguity in pitch assignment. For sufficiently close frequencies f1/f2 = 1.03, only one pitch, corresponding to the mean stimulus frequency, is extracted. (Inset) The time-dependent height of this histogram peak, indicating the occurrence of beats in the loudness. (b) Pitch discrimination. The locations of maxima in the histograms of inverse inter-spike intervals are marked by points (for larger values of f1/f2, when two main peaks are clearly resolved, each peak is split; the subsidiary maximum is marked by a smaller point). Gray bars provide an indication of the width of the peaks: they extend over the range for which the histogram height exceeds one-tenth of the peak height. The diagram shows that for f1/f2 > 1.15, the two pitches can be clearly discriminated. For small f1/f2 < 1.05, a single pitch is inferred and beats occur. In the intermediate range, two pitches are distinguished but they are more ambiguous, suggesting a sensation of roughness.
They also add to our understanding of the enigmatic relation between harmony and the ratio of small integers, on which musical scales are based. Helmholtz (1) overturned the Pythagorean doctrine by arguing that consonant intervals are not perfect harmonies, but simply less jarring dissonances. He ascribed dissonance to close, but inexact, matches in frequency between some of the harmonics that are generated when notes are played on a musical instrument. Subsequent experiments using pure tones lent weight to his argument (24). No preference for integer frequency ratios was expressed; rather, the roughness of two tones was found to be most intrusive when the difference amounted to a few percent and to diminish smoothly as Δf increased. Helmholtz attributed the roughness to beats, but his explanation is unsatisfactory because it fails to explain why dissonance persists above absolute frequencies of 1 kHz, when the beats are too fast to be distinguished. It is little surprise that his linear argument proves inadequate for a nonlinear system. We argue that dissonance arises from the difficulty of inferring frequency components from partial information about a complex waveform, which results in an indeterminancy of pitch. Both dissonance and pitch discrimination depend on the degree of interference between the two tones in the cochlea, so we would expect the interval of dissonance to have the same dependence on frequency as the interval of pitch discrimination, as is indeed observed (24). We also predict that these intervals should diminish at low intensities, when the active amplifier is most effective at sharpening the filter.
In addition, our analysis accounts for two different types of auditory illusion and indicates that they have distinct origins. Many investigators have confirmed that the DP frequencies can be heard (25). Although the DPs are generated by the Hopf resonance, they are not produced as the strongest component. Thus the active amplifier cannot, by itself, account for their audibility. Nevertheless, any nonlinearity in the prefilter would generate small DP components (1), which subsequently would be magnified by oscillators of the corresponding characteristic frequency. The Tartini effect, then, can be explained by the combination of a nonlinear prefilter and active amplifier. The second type of illusion is the residue pitch (26, 27). When f1 and f2 are neighboring harmonics, this pitch is identified as the missing fundamental Δf. But when the frequencies are less simply related, one or more pitches close to, but not equal to, Δf are heard. These pitches are derived from the complex waveform of hair-bundle motion and appear as peaks in the histogram of spike intervals (Fig. 5). This adds substance to previous suggestions (26–28) that they are artefacts that arise from the coding of hair-bundle vibration as a time series of nervous spikes.
Figure 5.

Residue pitch. In the presence of two stimulus frequencies f1 and f2 = f1 + Δf, one or more pitches in the vicinity of the frequency Δf can be extracted. (a) Displayed are subsidiary peaks in the histogram of the reciprocal, T−1, of the inter-spike intervals, in the vicinity of the pitch that corresponds to Δf. The responses to three different combinations of stimuli frequency are shown (f2:f1 = 120:100, 130:110 and 135:115), indicating that the inferred residue pitch varies as a function of f1/Δf. (b) Comparison between the location of histogram peaks (red bars) and the experimentally determined perceived pitch of residues (black and gray circles; data of ref. 27 for two different subjects).
Summary
In summary, we have provided a unifying physical description of two-tone interference effects and shown that many aspects of our perception of sound may be traced to the physiology of the inner ear, where they originate in one (or a combination) of the three stages of sound detection: prefiltering by the basilar membrane, active amplification by hair cells, and neural coding of hair-bundle motion.
Acknowledgments
We thank S. Camalet, E. F. Evans, A. J. Hudspeth, P. Martin, and J. Prost for fruitful discussions. F.J. is grateful for the hospitality of the Cavendish Laboratory and acknowledges support from the Engineering and Physical Sciences Research Council. T.D. is a Royal Society University Research Fellow.
Abbreviation
- DP
distortion product
Numerical Model
Active Amplifier.
Numerical results are solutions of the complex differential equation
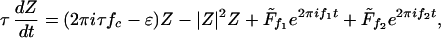 |
where τ is a time scale and the Hopf bifurcation occurs for ɛ = 0. We use frequencies for which f1/Δf is integer and define the response X = x0Re(Z). The amplitudes Xf = 2Δf ∫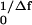 dtX(t)e−2πift, obtained by a Fourier transform of the limit cycle, satisfy Eqs. 1, 2, A4, and A5 with α = (2πiτ/x0)(Ff/F̃f) and B = B̄/2 = C = α/(2πiτx
dtX(t)e−2πift, obtained by a Fourier transform of the limit cycle, satisfy Eqs. 1, 2, A4, and A5 with α = (2πiτ/x0)(Ff/F̃f) and B = B̄/2 = C = α/(2πiτx ). The active bandwidth Δfa is defined as the value of |f − fc| for which the single-frequency response falls to half the peak amplitude. It varies with force, Δfa ∼ F
). The active bandwidth Δfa is defined as the value of |f − fc| for which the single-frequency response falls to half the peak amplitude. It varies with force, Δfa ∼ F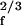 : comparison with the experimental basilar membrane response (4) suggests that Δfa/fc = 0.01, 0.1, and 1 correspond, respectively, to sound pressure levels 10, 40, and 70 db. Data in Figs. 1 and 2 were obtained with τfc = 1, x0 = 1, slightly on the oscillating side of the bifurcation, ɛ = −10−3. In Fig. 2, F̃f1 = F̃f2 = 0.5, giving Δfa/fc ≃ 1.22 (a high-level stimulus). In Figs. 3 and 4, a moderate stimulus was used, such that Δfa/fc ≃ 0.05. Data in Fig. 5 were obtained by using stimuli of slightly higher amplitude, corresponding to Δfa/fc = 0.1.
: comparison with the experimental basilar membrane response (4) suggests that Δfa/fc = 0.01, 0.1, and 1 correspond, respectively, to sound pressure levels 10, 40, and 70 db. Data in Figs. 1 and 2 were obtained with τfc = 1, x0 = 1, slightly on the oscillating side of the bifurcation, ɛ = −10−3. In Fig. 2, F̃f1 = F̃f2 = 0.5, giving Δfa/fc ≃ 1.22 (a high-level stimulus). In Figs. 3 and 4, a moderate stimulus was used, such that Δfa/fc ≃ 0.05. Data in Fig. 5 were obtained by using stimuli of slightly higher amplitude, corresponding to Δfa/fc = 0.1.
Passive Prefilter.
Results in Figs. 3 and 4 were obtained by using a passive prefilter χ(f), which multiplies Ff before excitation of the Hopf resonance. This is equivalent to the coefficients α and B in Eqs. 1 and 2 having the functional form χ−1(f). The form χ(f) = [(f − fc)2/(Δfp)2 + 1]−1 with bandwidth Δfp/fc = 0.15 is a fair approximation of the postmortem amplitude-response of the basilar membrane (3). The prefilter suppresses two-tone interferences for Δf > Δfp. It does not affect the relative amplitudes of DPs.
Neural Response.
Data in Figs. 4 and 5 were obtained by using a set of 100 oscillators with characteristic frequencies in the range fc/f1 = 0.5 − 1.5. A neural spike was elicited by an oscillator every time its response X traversed a given threshold. Histograms of inter-spike intervals were constructed, averaging over all positive thresholds for each oscillator and then summing over all oscillators.
Generic Two-Tone Distortions Near a Hopf Bifurcation
In the presence of a stimulus
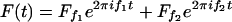 |
A1 |
containing two different frequencies f1 and f2, the response of a dynamical system close to a Hopf bifurcation contains all Fourier amplitudes with frequencies f = nf1 + mf2, where n and m are positive or negative integers. For simplicity, we choose stimuli with commensurate frequencies, for which f1/Δf is integer and the response is given by
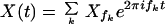 |
A2 |
with fk = f1 + (k − 1)Δf.
Close to the Hopf bifurcation and for small amplitudes Xf, we can write a general expansion of the form
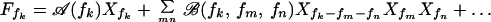 |
A3 |
Throughout this paper, we ignore quadratic terms that occur if the symmetry X → −X of the active system is broken; they renormalize the coefficients B and B̄ of the cubic terms in Eq. A4 below and are only involved in the generation of higher harmonics and difference tones. For small stimulus and large Δf, the dominant terms in the expansion (A3) for the response at frequencies f1 and f2 is given by
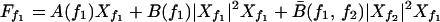 |
A4 |
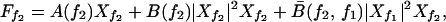 |
where A(f) = 𝒜(f), B(f) = ℬ(f, f, f) and B̄(f1, f2) = 2ℬ(f1, f1, f2). We choose one of the stimuli to be at the characteristic frequency, f1 = fc, therefore A(f1) = 0. For Ff1 = F and Ff2 = 0, it follows that Xf2 = 0 and the nonlinear single-tone response |Xf1| ≃ |F1/3B−1/3| is recovered. The stimulus Ff2 generates Xf2, which in the linear regime obeys Xf2 ≃ Ff2/αΔf. The mode Xf2 creates an effective linear term for Xf1 with Aeff ≃ B̄|Xf2|2. The mode Xf1 therefore behaves as if the system were mistuned and displays a suppressed response that is linear for small Ff1. Similarly, Xf1 creates an effective linear term that renormalizes A(f2) and suppresses the response Xf2.
Distortion products result from nonlinear terms X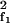 X*f2 and X
X*f2 and X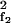 X*f1, which couple to the Fourier modes with frequencies 2f1 − f2 and 2f2 − f1. For example, in the absence of a stimulus of frequency 2f1 − f2, i.e. F2f1−f2 = 0, the corresponding amplitude obeys
X*f1, which couple to the Fourier modes with frequencies 2f1 − f2 and 2f2 − f1. For example, in the absence of a stimulus of frequency 2f1 − f2, i.e. F2f1−f2 = 0, the corresponding amplitude obeys
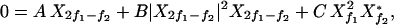 |
A5 |
where C = ℬ(2f1 − f2, f1, − f2). This leads to a DP amplitude |X2f1−f2| ≃ |C/αΔf∥X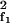 Xf2|. This mode together with Xf1 generates, via the same coupling, the DP at 3f1 − 2f2. Recursively, we therefore obtain a hierarchy of DPs with frequencies fk = f1 + (k − 1)Δf, whose amplitudes decay exponentially according to Eq. 3 with λ ≃ ln(7Δf/12Δfa) for Δf ≫ Δfa.
Xf2|. This mode together with Xf1 generates, via the same coupling, the DP at 3f1 − 2f2. Recursively, we therefore obtain a hierarchy of DPs with frequencies fk = f1 + (k − 1)Δf, whose amplitudes decay exponentially according to Eq. 3 with λ ≃ ln(7Δf/12Δfa) for Δf ≫ Δfa.
For smaller Δf, the response involves a large number of terms of the expansion (A3) and deviations from a pure exponential appear. An interesting limit occurs for Δf → 0 where the linear coefficients A(fk) ≃ αΔf(k − 1) and the frequency dependence of ℬ(f, f′, f") can be neglected for a large number of modes around f1. With this approximation Eq. A3 becomes
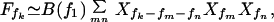 |
A6 |
which corresponds to X(t) ≃ B−1/3F1/3(t). The spectrum of X(t) exhibits, for large |k|, a power law decay of DPs, Xfk ∼ |k − 3/2|−v, centered around the critical frequency. This can be demonstrated explicitly in the case Ff1 = Ff2 = F, for which the response to a two-tone stimulus for small Δf is X(t) ≃ (2F/B)1/3cos1/3(2π(f1 + Δf/2)t)cos1/3(πΔft). The spectrum Xfk in this limit is therefore simply given by the convolution of the spectrum Cn with itself, where Cn are the Fourier components of C(t) ≡ cos1/3(2πft) = ∑nCne2πinft. These Fourier components decay for large n as Cn ∼ n−v with v = 4/3. This power law reflects singularities of dC/dt. Indeed, at the zeros t0 for which C(t0) = 0, dC/dt ∼ (t − t0)−3/2 diverges with a power law that determines the exponent v.
References
- 1.Helmholtz H L F. On the Sensations of Tone. New York: Dover; 1954. [Google Scholar]
- 2.Rhode W S. J Acoust Soc Am. 1971;49:1218–1231. doi: 10.1121/1.1912485. [DOI] [PubMed] [Google Scholar]
- 3.Sellick P M, Patuzzi R, Johnstone B M. J Acoust Soc Am. 1982;72:131–141. doi: 10.1121/1.387996. [DOI] [PubMed] [Google Scholar]
- 4.Ruggero M A, Rich N C, Recio A, Narayan S S, Robles L. J Acoust Soc Am. 1997;101:2151–2163. doi: 10.1121/1.418265. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 5.Robles L, Ruggero M A, Rich N C. Nature (London) 1991;349:413–414. doi: 10.1038/349413a0. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 6.Robles L, Ruggero M A, Rich N C. J Neurophysiol. 1997;77:2385–2399. doi: 10.1152/jn.1997.77.5.2385. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 7.Ruggero M A, Robles L, Rich N C. J Neurophysiol. 1992;68:1087–1099. doi: 10.1152/jn.1992.68.4.1087. [DOI] [PubMed] [Google Scholar]
- 8.Rhode W S, Cooper N P. Hearing Res. 1993;66:31–45. doi: 10.1016/0378-5955(93)90257-2. [DOI] [PubMed] [Google Scholar]
- 9.Sachs M B, Kiang N Y S. J Acoust Soc Am. 1967;43:1120–1129. doi: 10.1121/1.1910947. [DOI] [PubMed] [Google Scholar]
- 10.Jaramillo F, Markin V S, Hudspeth A J. Nature (London) 1993;364:527–529. doi: 10.1038/364527a0. [DOI] [PubMed] [Google Scholar]
- 11.Hudpeth A J, Choe Y, Mehta A D, Martin P. Proc Natl Acad Sci USA. 2000;97:11765–11772. doi: 10.1073/pnas.97.22.11765. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 12.Dallos P. J Neurosci. 1992;12:4575–4585. doi: 10.1523/JNEUROSCI.12-12-04575.1992. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 13.Gold T. Proc R Soc London Ser B. 1948;135:492–498. [Google Scholar]
- 14.Camalet S, Duke T, Jülicher F, Prost J. Proc Natl Acad Sci USA. 2000;97:3183–3188. doi: 10.1073/pnas.97.7.3183. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 15.Eguiluz V M, Ospeck M, Choe Y, Hudspeth A J, Magnasco M O. Phys Rev Lett. 2000;84:5232–5235. doi: 10.1103/PhysRevLett.84.5232. [DOI] [PubMed] [Google Scholar]
- 16.Zwicker E, Fastl H. Psychoacoustics. Berlin: Springer; 1999. [Google Scholar]
- 17.Pfeiffer R R. J Acoust Soc Am. 1970;48:1373–1378. doi: 10.1121/1.1912294. [DOI] [PubMed] [Google Scholar]
- 18.Bourbon W T, Evans T R, Deatherage B H. J Acoust Soc Am. 1968;43:56–59. doi: 10.1121/1.1910762. [DOI] [PubMed] [Google Scholar]
- 19.von Békésy G. Experiments in Hearing. New York: McGraw–Hill; 1960. [Google Scholar]
- 20.Russel I J, Nilsen K E. Proc Natl Acad Sci USA. 1997;94:2660–2664. doi: 10.1073/pnas.94.6.2660. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 21.Rose J E, Brugge J F, Anderson D J, Hind J E. J Neurophysiol. 1967;30:769–793. doi: 10.1152/jn.1967.30.4.769. [DOI] [PubMed] [Google Scholar]
- 22.Evans E F. Philos Trans R Soc London B. 1992;336:295–306. doi: 10.1098/rstb.1992.0062. [DOI] [PubMed] [Google Scholar]
- 23.Winter I M, Robertson D, Yates G K. Hearing Res. 1990;45:191–202. doi: 10.1016/0378-5955(90)90120-e. [DOI] [PubMed] [Google Scholar]
- 24.Plomp R, Levelt W J M. J Acoust Soc Am. 1965;38:548–560. doi: 10.1121/1.1909741. [DOI] [PubMed] [Google Scholar]
- 25.Plomp R. J Acoust Soc Am. 1965;37:1110–1123. doi: 10.1121/1.1909532. [DOI] [PubMed] [Google Scholar]
- 26.Schouten J F, Ritsma R J, Lopes Cardozo B. J Acoust Soc Am. 1962;34:1418–1424. [Google Scholar]
- 27.Smoorenburg G F. J Acoust Soc Am. 1970;48:924–942. doi: 10.1121/1.1912232. [DOI] [PubMed] [Google Scholar]
- 28.Rose J E, Brugge J F, Anderson D J, Hind J E. J Neurophysiol. 1968;32:402–423. doi: 10.1152/jn.1969.32.3.402. [DOI] [PubMed] [Google Scholar]