

Abstract
Fifty nine candidate olfactory receptor (Or) genes have recently been identified in Drosophila melanogaster, one of which is Or43a. In wild-type flies, Or43a is expressed at the distal edge of the third antennal segment in about 15 Or neurons. To identify ligands for the receptor we used the Gal4/UAS system to misexpress Or43a in the third antennal segment. Or43a mRNA expression in the antenna of transformed and wild-type flies was visualized by in situ hybridization with a digoxigenin-labeled probe. Electroantennogram recordings from transformed and wild-type flies were used to identify cyclohexanol, cyclohexanone, benzaldehyde, and benzyl alcohol as ligands for the Or43a. This in vivo analysis reveals functional properties of one member of the recently isolated Or family in Drosophila and will provide further insight into our understanding of olfactory coding.
The ability to identify a huge diversity of ordorants is essential for the survival of many animals. Diverse transduction mechanisms contribute to the initial events underlying olfactory perception in the primary olfactory receptor neurons (ORNs) in vertebrates and invertebrates. More than 1,000 different odorants were shown to attract the insect species Daus dorsalis (1). Also fruit flies have been shown to be capable of odor discrimination (2). The initial steps in order perception take place at the olfactory receptor (Or) localized in the ORNs.
Recently, in Drosophila melanogaster, 59 genes were found based on structural criteria that identified them as putative Or genes (3–5). These receptor genes encode for seven transmembrane-domain proteins, which exhibit low homology to known vertebrate and invertebrate Ors. By in situ hybridization RNA of 32 Ors were found in the soma of ORNs of the third antennal segment and seven exclusively in the maxillary palps, a second olfactory organ in Drosophila. ORNs are situated in different types of sensory hairs, which are arranged in a specific manner on the surface of the antenna. Each sensory hair bears 2–4 ORNs that send their afferent projections to one of the 43 glomeruli in the antennal lobe, the first olfactory processing center (6). Using transformed flies, Vosshall et al. (7) and Gao et al. (8) could show that subsets of ORNs express only one type of Or and that individual ORNs with the same Or converge to the same glomerulus. This organization reflects the known model of odortopic representation both in the antennal lobe and on the third antennal segment. Thus far, none of these putative Ors have been proven to have olfactory function.
In this study we identified ligands for the Or Or43a and thereby give evidence for its olfactory function. For this approach we used the GAL4/UAS system to misexpress Or43a, described previously as DOR87 or 43B1 (3–5), in the third antennal segment. Or43a is expressed at the distal edge of the third antennal segment and is not found in the maxillary palps. Increasing the amount of ORNs that express this receptor should result in an elevated electroantennogram (EAG) when stimulated with a ligand. By in vivo measurements, we show that responses to benzaldehyde (Bz), benzyl alcohol (Bzl), cyclohexanol (Cyl), and cyclohexanone (Cyc) are increased.
Materials and Methods
Fly Stocks and Construction of Transformants.
Flies were reared on a standard cornmeal-molasses-agar medium and kept at 25°C in an incubator with 12-h day/night cycle. Wild-type Oregon R and transformed flies with yellow-white mutant background were used for in situ hybridization experiments and EAG recordings from the antenna.
The complete reading frame of Or43a or Or22b was cloned into pUAST vector to generate the final transformation vector. yellow-white flies were transformed by using standard procedures. Three of the original transformed individuals were used to derive the tested strains designated as UAS-Or43a-1c, UAS-Or43a-3c, and UAS-Or22b. These lines had P-element insertions in chromosomes 1 or 3, respectively. All UAS lines were homozygous viable and were crossed with the Gal4 driver line GH320 to generate 87-1c, 87-3c, and 67-3c as was done with the UAS-LacZ line to isolate a Gal4/UAS-LacZ line.
In Situ Hybridization and Immunocytochemistry.
In situ hybridization was carried out as described by Vosshall et al. (5). Briefly, flies mounted in OCT (Microm, Walldorf, Germany) were shock-frozen in liquid nitrogen, and sections of 10 μm were tested. After fixation with 4% paraformaldehyde, acetylation, and subsequent prehybridization of sections was carried out at 58°C for 1 h. Hybridization with the biotinylated Or43a probe was performed overnight at 58°C. Immuncytochemical labeling was accomplished according to Störtkuhl et al. (9). After blocking with normal goat serum in phosphate buffer (PBS + 0.1% Triton X-100) the digoxigenin probe was stained with an alkaline phosphatase-coupled antidigoxigenin antibody (1:1,000 in PBS + 0.1% Triton X-100) and color development was controlled under a Zeiss microscope for 20–60 min.
Double-labeling of cryosections from antenna with anti-elav and anti-LacZ antibody was carried out according to Störtkuhl et al. (9). Flies were fixed in 4% paraformaldehyde for 3 h at 4°C. After blocking with 1% BSA, the monoclonal anti-elav antibody and the polyclonal anti-LacZ antibody (Rockland, Gilbertsville, PA) were diluted 1:100 and 1:1,000, respectively, in PBS + 1% Triton X-100. Flies were incubated overnight at 4°C in the antibody solution. After washing in phosphate buffer, and subsequent incubation with a second fluorescence-coupled antibody (anti-rabbit Cy3-coupled antibody to identify anti-LacZ antibody and anti-mouse Cy2-coupled antibody to identify anti-elav antibody), staining was analyzed under a Zeiss microscope. Using a fluorescence filter system with an excitation of 485 nm and 578 nm, fluorescence of both secondary antibodies could be analyzed simultaneously.
Electrophysiological Recording on the Antenna.
To measure electrophysiological responses to different odorants on the antenna in vivo, we recorded EAGs using the method described in refs. 10–12. Briefly, 1-day-old flies were mounted in truncated micropipette tips with the anterior portion of the head protruding from the end of the tip. The tip was seated on wax in the middle of a Petri dish that was filled with Drosophila Ringer and was grounded by an electrode immersed by the liquid that filled the Petri dish. The indifferent electrode was inserted into the hemolymph of the head capsule. The recording electrode was placed on the frontal surface of the anterior aspect of the antenna. After obtaining a stable baseline, EAG recordings were initiated by a short pulse of odor, applied through a syringe into an air stream (1 liter/min) that was directed toward the antenna. All odorants were dissolved in paraffin oil at the given concentrations with the exception of 1,4-cyclohexanedione and vanillin, which were dissolved in water (0.25 g/ml = saturated solution). Odorants used were acetone (J. T. Baker), Bz (Fluka), Bzl, butanol (J. T. Baker), cinnamyl alcohol (Riedel-de Häen, Seelze, The Netherlands), Cyl (J. T. Baker), Cyc (J. T. Baker), 1,4-cyclohexanedione (Fluka), ethylacetate (Ea; J. T. Baker), hexanone (Fluka), propionaldehyde (J. T. Baker), and vanillin (Fluka).
Results and Discussion
Or43a Misexpression in the Adult Antenna.
To demonstrate olfactory properties for Ors in Drosophila, we ectopically expressed one candidate Or, Or43a (Drosophila Odorant Receptor Nomenclature Committee, 2000). Using the Gal4/UAS technique (13, 14), we increased the number of ORNs that expresses that Or. We reasoned that an electrophysiological response measured from such transformed flies would be elevated for ligands specific to the Or, whereas unspecific odorants would evoke responses unchanged in wild-type flies.
We used receptor gene Or43a, described previously as DOR87 or 43B1, that is expressed in wild-type flies, in a limited number of cells (15) at the distal edge of the antenna (5) (Fig. 1A) for this approach. To generate transformed flies, in which Or43a was ectopically expressed in additional ORNs, we cloned cDNA Or43a (a kind gift from L. Vosshall, Rockefeller University, New York) into the pUAST vector. We isolated two lines, UAS-Or43a-1c and UAS-Or43a-3c, that contained Or43a cDNA and crossed these flies with a Gal4 line, GH320 (a kind gift from G. Heimbeck, University of Fribourg, Fribourg, Switzerland), that expresses Gal4 in ORNs of the third antennal segment (see below). The progeny of these crosses were the transformed lines designated 87-1c and 87-3c. These lines exhibited Gal4-driven ectopic Or43a expression in the third antennal segment as revealed by in situ hybridization with a digoxigenin-labeled antisense probe. Compared with wild-type, the number of Or43a-expressing cells was drastically increased in 87-1c and 87-3c (Fig. 1B). In addition to the cells at the distal edge of the antenna, which are labeled in wild-type flies, there were Or43a-expressing cells in the transformed flies in a more proximal region that is covered mainly by large sensilla basiconica.
Figure 1.

Or43a is expressed in neuronal cells. mRNA expression of Or43a in a 10-μm thick cryosection through the third antennal segment of (A) wild type and (B) line 87-3c as visualized by in situ hybridization with a digoxigenin-labeled cDNA probe. Although few cells are stained in wild type, the number of Or43a-expressing cells is increased in the antenna of 87-3c. Immunostaining of 10-μm horizontal cryosections of the antenna. (C) Interphase contrast picture of the third antennal segment of the Gal4/UAS-LacZ line. (D) The same section with anti Elav-antibody (green) as neuronal marker and with (E) anti LacZ-antibody (red). (F) Fluorescence labeling was visualized with a Zeiss filter Nr.24 (emission at 460 nm, 530 nm, and 610 nm). Colocalization of Elav and LacZ can be identified by yellow color indicating neuronal expression of both proteins. Few neuronal cells do not have LacZ expression (arrows). (Bar = 5 μm.)
Before testing flies from the transformed lines 87-1c and 87-3c, we wanted to verify that the ectopic expression of Or43a in the antenna was in ORNs. Because there is no anti-Or43a antibody available to visualize Or43a expression in the antenna directly, we alternatively determined LacZ expression in the driver line GH320. Double labeling was performed in a GH320/UAS LacZ line to show Gal4-driven LacZ expression in the antenna. Elav antibody was used to identify neuronal cells. Labeling with anti-LacZ as well as anti-Elav antibody was ubiquitous over the entire antenna (Fig. 1 D and E). Colocalization (Fig. 1F) of both antigens indicated Gal4 expression in the third antennal segment, mainly in neuronal cells. From these observations we concluded that in the transformed lines 87-1c and 87-3c Or43a was expressed ectopically in ORNs.
Misexpression of Or43a in the Antenna Revealed the Ligand Bz.
To identify the ligands for Or43a, we recorded EAGs from the lines 87-1c and 87-3c as well as from wild-type and parental lines (GH320, and transformants: UAS-Or43a-1c and UAS-Or43a-3c). In addition, we used a control line 67-3c to exclude unspecific increase of the EAG response based on different genetic background (9). This line misexpressed a different Or, namely OR22b, in the whole antenna (data not shown). We chose a number of odorants, many of which have been used as biological relevant stimuli in previous experiments (15–19). EAG recordings of 87-1c and 87-3c showed increased responses in the EAG to Bz (Fig. 2 A and B). Bz diluted 10−2 in paraffin oil elicited an EAG amplitude of 12.42 mV (±0.87 SEM) or 13.70 mV (±0.45 SEM) in 87-1c and 87-3c, respectively, and a significantly lower amplitude in wild-type flies (8 mV ± 0.44 SEM) and line 67-3c (8.43 mV ± 0.47 SEM). In contrast, butanol, acetone, Ea, and propionaldehyde elicited EAGs that were similar in amplitude in all Gal4/UAS lines and wild type (Fig. 2C). For further experiments we used line 87-3c because it exhibited the highest increment in the EAG response to Bz.
Figure 2.

Increased EAG response of lines 87-1c and 87-3c to Bz. (A) Average of 10 EAG-recordings each from 1) transformed line 87-3c misexpressing Or43a and 2) wild-type flies. (Upper) Black bar indicates the given odorant pulse (1.0 sec) with a concentration of 10−2 in paraffin oil. (B) EAG response of wild type, Or43a misexpressing lines 87-1c and 87-3c, and Or22b misexpressing line 67-3c to Bz diluted 10−2 in paraffin oil (n = 20) for each genotype and (C) to acetone (Ac), butanol (But), Ea, and propionaldehyde (Pa), all diluted 10−2 in paraffin oil (oil). n = 20 for each genotype tested with each odorant.
The increased response of line 87-3c to Bz is concentration dependent (Fig. 3A). Or43a-misexpressing flies (87-3c) as well as parental lines (UAS- Or43a-3c; GH320) showed electrophysiological responses that were not significantly different from responses of wild-type flies when tested with dilutions of Bz higher than 10−3. However, application of higher concentrations of Bz elicited significantly higher responses in the transformed line 87-3c (Fig. 3A). In response to a 10−1 dilution of Bz, line 87-3c had an EAG amplitude that was twice as high as that of wild-type flies, 16.89 mV ± 0.31 SEM for 87-3c versus 8.89 ± 0.34 SEM for wild type. There was no further increment in the EAG of either wild-type or 87-3c flies that were stimulated with undiluted Bz (Fig. 3A). The odorant Ea, in contrast, evoked similar responses in control and transformed flies at all concentrations tested (Fig. 3B).
Figure 3.

Increased EAG response to Bz is concentration dependent. EAG response to different concentrations of (A) Bz and (B) Ea dissolved in paraffin oil. In most cases error bars are too small to be shown on the graphs. GH230 and UAS-Or43a-3c are parental lines whereas 87-3c misexpresses Or43a. n = 25 tests for each genotype.
Our results are in good agreement with previous findings from both behavior experiments and electrophysiological measurments. Several mutants have been isolated that have a defect in the perception of Bz but were still able to respond normally to Ea or vice versa (16, 18).
Moreover there is an odortopic presentation on the antenna. Siddiqi (20) showed that the distal part of the antenna possesses a high sensitivity for Bz and Cyc, which is suitable with that region, and Or43a mRNA is detectable by in situ hybridization. In contrast, Ea sensitivity was detected at a proximal portion of the antenna.
Or43a Recognizes Cyclic Molecules with a Polar Group.
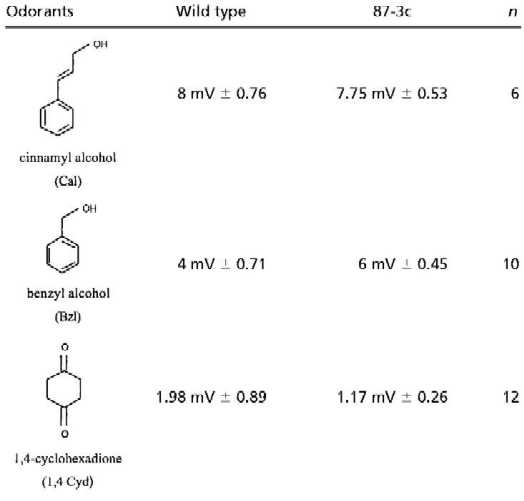
Is Bz the only ligand for the Drosophila Or Or43a? Drosophila is thought to sense hundreds of odorants, suggesting that some of the 59 candidate Ors, at least, bind more than one ligand. To determine whether this is the case for Or43a, we tested the structurally related chemicals Cyl, Cyc, and Bzl. We also tested flies with hexane and hexanol, which are composed of a six-carbon chain. All of the compounds with a cyclic ring elicited increased electrophysiological responses in the EAG of transformed flies (Fig. 4 and Table 1). The most dramatic effect was seen with Cyl (Fig. 4A). Dilutions of 10−2 elicited EAGs with amplitudes of 14.00 mV (±0.63 SEM) in transformed flies versus 5.63 mV (±1.00 SEM) in wild-type flies. Line 87-3c showed a dosage-dependent increase of the EAG to both Cyc and Cyl (Fig. 4). Application of these substances with increasing concentration led to increasing increment of the EAG of transformed flies. In contrast, stimulation with a 10−2 dilution of hexanone or hexanol elicited responses in transformed flies that were not significantly different from those of wild-type flies (Table 1).
Figure 4.

EAG response to different concentrations for (A) Cyl and (B) Cyc dissolved in paraffin oil. In most cases error bars are too small to be shown on the graphs. n = 25 tests for each genotype.
Table 1.
EAG of wild-type and transformant 87-3c
 |
|---|
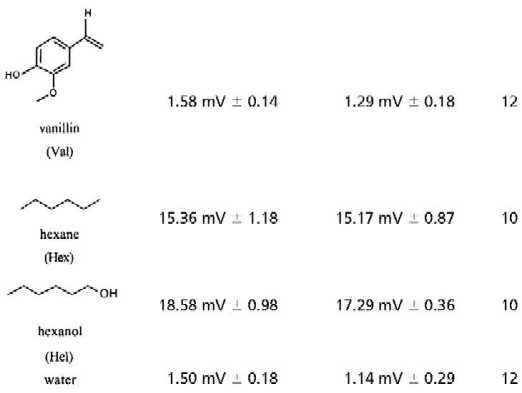 |
Mean amplitudes of EAG responses in mV from wild-type and transformed-line 87-3c. Odorants were tested at a concentration of 10−2 diluted in paraffin oil with the exception of 1,4 cyclohexadione and vanillin, which were saturated in water (0.25 g/ml). n = number of tested flies; ± is SEM.
What happens if one increases the number of polar groups attached to the six-carbon ring? The water-soluble compound 1,4-cyclohexanedione with two polar oxygen groups, and vanillin with two more polar groups bound to the cyclic ring, elicited EAG responses in transformed and wild-type flies that were not higher than those obtained by water (Table 1). Similarly, cinnamyl alcohol with an extended polar residue attached to the cyclic ring showed no elevated EAG in transformed flies (Table 1). Obviously, additional polar group or a prolonged polar residue attached to a six-carbon ring inhibits these odorants from binding to the receptor Or43a.
In summary, we identified four ligands, Cyl, Bz, Cyc, and Bzl, for Or43a by using an in vivo system and by detecting an increased amplitude in the antennal EAG to these ligands. The EAG measured reflects the overall electrophysiological activity recorded at the proximal portion of the antenna. Therefore we cannot exclude modulations of the EAG amplitude based on cellular interactions in the antenna. In contrast to the methods used to identify ligands in mammals (21–25) we used transformed flies that ectopically expressed Or43a in ORNs of the third antennal segment. These identified odorants have in common a six-carbon ring and one polar group attached to the ring. Further evidence for ligand specificity is provided from a heterologous system.
Flies' Ors are, in at least one respect, similar to those of higher organisms. They obviously are also able to recognize multiple odorants, as shown by analysis of Or43a. Further studies with this straightforward approach for identifying ligands without the necessity of having mutant alleles of Ors will show whether other Drosophila Ors are similar to Or43a with respect to ligand perception.
Acknowledgments
We are very grateful to S. Wagner for excellent technical assistance. This work was performed in the laboratory of B. Hovemann. We thank M. Heisenberg, B. Hovemann, L. Tompkins, and V. Rodrigues for offering valuable comments on the manuscript. We also grateful to L. Vosshall and G. Heimbeck for giving us clones Or34a and Or22b and Gal4 line GH320, respectively. This work was supported by the Deutsche Forschungsgemeinschaft (HO 714/9–1) to B. Hovemann.
Abbreviations
- Bz
benzaldehyde
- Bzl
benzyl alcohol
- Cyl
cyclohexanol
- Cyc
cyclohexanone
- Ea
ethylacetate
- EAG
electroantennogram
- Or
olfactory receptor
- ORN
Or neuron
Footnotes
This paper was submitted directly (Track II) to the PNAS office.
See commentary on page 8936.
References
- 1.Beroza M, Green N. Materials Tested as Insect Attractants. U.S. Department of Agriculture, Washington, DC: Agricultural Research Service; 1963. [Google Scholar]
- 2.Siddiqi O. Olfactory Neurogenetics of Drosophila. New Delhi: Oxford; 1983. [Google Scholar]
- 3.Clyne P J, Warr C G, Freeman M R, Lessing D, Kim J, Carlson J R. Neuron. 1999;22:327–338. doi: 10.1016/s0896-6273(00)81093-4. [DOI] [PubMed] [Google Scholar]
- 4.Gao Q, Chess A. Genomics. 1999;60:31–39. doi: 10.1006/geno.1999.5894. [DOI] [PubMed] [Google Scholar]
- 5.Vosshall L B, Amrein H, Morozov P S, Rzhetsky A, Axel R. Cell. 1999;96:725–736. doi: 10.1016/s0092-8674(00)80582-6. [DOI] [PubMed] [Google Scholar]
- 6.Stocker R F. Cell Tissue Res. 1994;275:3–26. doi: 10.1007/BF00305372. [DOI] [PubMed] [Google Scholar]
- 7.Vosshall L B, Wong A M, Axel R. Cell. 2000;102:147–159. doi: 10.1016/s0092-8674(00)00021-0. [DOI] [PubMed] [Google Scholar]
- 8.Gao Q, Yuan B, Chess A. Nat Neurosci. 2000;3:780–785. doi: 10.1038/77680. [DOI] [PubMed] [Google Scholar]
- 9.Störtkuhl K F, Hofbauer A, Keller V, Gendre N, Stocker R F. Cell Tissue Res. 1994;275:27–38. doi: 10.1007/BF00305373. [DOI] [PubMed] [Google Scholar]
- 10.Ayer R K, Jr, Carlson J. J Neurobiol. 1992;23:965–982. doi: 10.1002/neu.480230804. [DOI] [PubMed] [Google Scholar]
- 11.Riesgo-Escovar J, Raha D, Carlson J R. Proc Natl Acad Sci USA. 1995;92:2864–2868. doi: 10.1073/pnas.92.7.2864. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 12.Störtkuhl K F, Hovemann B T, Carlson J R. J Neurosci. 1999;19:4839–4846. doi: 10.1523/JNEUROSCI.19-12-04839.1999. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 13.Brand A H, Perrimon N. Development (Cambridge, UK) 1993;118:401–415. doi: 10.1242/dev.118.2.401. [DOI] [PubMed] [Google Scholar]
- 14.Brand A H, Manoukian A S, Perrimon N. Methods Cell Biol. 1994;44:635–654. doi: 10.1016/s0091-679x(08)60936-x. [DOI] [PubMed] [Google Scholar]
- 15.Lilly M, Carlson J. Genetics. 1990;124:293–302. doi: 10.1093/genetics/124.2.293. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 16.Ayyub C, Paranjape J, Rodrigues V, Siddiqi O. J Neurogenet. 1990;6:243–262. doi: 10.3109/01677069009107114. [DOI] [PubMed] [Google Scholar]
- 17.Ayer R K, Jr, Carlson J. Proc Natl Acad Sci USA. 1991;88:5467–5471. doi: 10.1073/pnas.88.12.5467. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 18.Helfand S L, Carlson J R. Proc Natl Acad Sci USA. 1989;86:2908–2912. doi: 10.1073/pnas.86.8.2908. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 19.Charro M J, Alcorta E. J Comp Physiol A. 1994;175:761–766. doi: 10.1007/BF00191847. [DOI] [PubMed] [Google Scholar]
- 20.Siddiqi O. Trends Genet. 1987;3:137–142. [Google Scholar]
- 21.Araneda R C, Kini A D, Firestein S. Nat Neurosci. 2000;3:1248–1255. doi: 10.1038/81774. [DOI] [PubMed] [Google Scholar]
- 22.Mori K, Nagao H, Yoshihara Y. Science. 1999;286:711–715. doi: 10.1126/science.286.5440.711. [DOI] [PubMed] [Google Scholar]
- 23.Duchamp-Viret P, Chaput M A, Duchamp A. Science. 1999;284:2171–2174. doi: 10.1126/science.284.5423.2171. [DOI] [PubMed] [Google Scholar]
- 24.Krautwurst D, Yau K W, Reed R R. Cell. 1998;95:917–926. doi: 10.1016/s0092-8674(00)81716-x. [DOI] [PubMed] [Google Scholar]
- 25.Malnic B, Hirono J, Sato T, Buck L B. Cell. 1999;96:713–723. doi: 10.1016/s0092-8674(00)80581-4. [DOI] [PubMed] [Google Scholar]