


Abstract
Transcripts representing mRNAs of three Xenopus cyclins, B1, B4 and B5, were hybridised to arrays of oligonucleotides scanning the first 120 nt of the coding region to assess the ability of the immobilised oligonucleotides to form heteroduplexes with their targets. Oligonucleotides that produced high heteroduplex yield and others that showed little annealing were assayed for their effect on translation of endogenous cyclin mRNAs in Xenopus egg extracts and their ability to promote cleavage of cyclin mRNAs in oocytes by RNase H. Excellent correlation was found between antisense potency and affinity of oligonucleotides for the cyclin transcripts as measured by the array, despite the complexity of the cellular environment.
INTRODUCTION
Antisense oligonucleotides are single-stranded DNA sequences, typically 15–25 nt in length, that bind to complementary sites in mRNAs and inhibit translation by promoting cleavage of the RNA strand in mRNA–DNA hybrids by RNase H (1,2). They are regarded as powerful tools for studying the biological function of genes, as well as providing potential anti-virus and anti-cancer drugs (3,4). In practice, however, rather few oligonucleotides prove to be effective antisense reagents (5). Antisense efficiency depends on the ability of an oligonucleotide to hybridise with its target mRNA. There is compelling evidence to suggest that heteroduplex formation is primarily constrained by local secondary structure and folding of mRNAs; the composition and the length of the oligonucleotides seem to be of relatively less importance (6–11). Identification of the most accessible site of a given mRNA could therefore be a straightforward strategy to select potent antisense oligonucleotides. Unwanted non-specific cleavage of mRNAs is another common problem that will be exacerbated if high concentrations of oligonucleotide are used (12). Selection of the most effective oligonucleotide sequences that work at low concentrations is thus an important prerequisite to a wider application of the antisense approach (13).
It is not possible to predict the ability of oligonucleotides to form heteroduplexes with a mRNA from thermodynamic considerations alone, mainly because present methods for secondary structure prediction of mRNAs are not reliable. In living cells mRNA accessibility is less predictable as the folding of actively translated mRNAs is continually altered by passing ribosomes. Proteins that bind to mRNA may also contribute to problems in heteroduplex formation.
Oligonucleotide scanning arrays provide an empirical approach to choosing antisense oligonucleotides (8,11,14). By hybridising mRNAs to such arrays oligonucleotides of different length, targeted to many or all regions of the mRNA, can be assayed simultaneously for their ability to bind their target. No clear evidence exists, however, that hybridisation of naked transcripts to oligonucleotides provides a useful estimate of antisense efficiency in vivo. There is no reason a priori to suppose that it should, considering the crowded and heterogeneous conditions of cytoplasm compared to the simplicity of an in vitro annealing experiment that takes place at 1 M salt in the presence of EDTA.
We report here that hybridisation of mRNAs to oligonucleotide scanning arrays is an effective method to select antisense reagents with high potency in vivo.
MATERIALS AND METHODS
Synthesis of oligonucleotides
All oligonucleotides used in this study were deoxyribophosphodiesters made in an ABI 394 DNA/RNA synthesiser using standard nucleotide CE-phosphoramidites (Cruachem) on a 1 µmol scale. A 12mer oligonucleotide library (dN12) was made on a column containing CPG-amidite precursors for all four bases and by adding all four phosphoramidites at each position. It was also made on a 1 µmol scale. These were then desalted by Sephadex G-25 chromatography through NAP-10 columns (Amersham Pharmacia). Antisense oligonucleotides were desalted at least four times through NAP-10 columns. Purity of the oligonucleotides was evaluated on a 20% denaturing polyacrylamide gel after 5′-end-labeling with 32P in the presence of [γ-32P]ATP (>5000 Ci/mmol; Amersham) and T4 polynucleotide kinase (New England Biolabs).
Preparation of radiolabelled transcripts
Radiolabelled transcripts for probing the scanning arrays were obtained by in vitro transcription. The template plasmids were linearised with an appropriate restriction endonuclease. Transcriptions were carried out at 30°C for 90 min in the presence of 20 µCi [α-32P]UTP (∼3000 Ci/mmol; Amersham), 750 µM each ATP, CTP and GTP and 18.75 µM UTP with T7 RNA polymerase (Promega). The products were purified by filtration through MicroSpin G-25 columns (Pharmacia Biotech) and analysed on a 5% denaturing polyacrylamide gel. 5′-End-labelled transcripts for the RNase H mapping were prepared in the presence of 50 µCi [γ-32P]GTP (>5000 Ci/mmol; Amersham) essentially as described above, except that the concentration of unlabelled GTP was 75 µM and that of UTP was 750 µM.
Mapping of the cyclin B5 transcripts for accessible sites with RNase H/dN12
Each RNase mapping reaction contained 5 fmol 5′-end-labelled transcript, 10 mM MgCl2, 50 mM KCl, 50 mM Tris–HCl pH 7.4, 1 mM DTT, 0.5 U RNase H, 0.5 U RNase inhibitor (Promega) and dN12 (10, 20, 50, 100, 250 and 500 pmol, respectively) in a total volume of 10 µl. The reactions were incubated at 30°C for 30 min and terminated by addition of 10 µl of formamide gel loading dye buffer. A 5 µl aliquot was analysed on a 6% denaturing polyacrylamide gel.
Fabrication of the antisense oligonucleotide scanning array
The arrays were made essentially as described (11), except that aminated polypropylene (Beckman Instruments, Fullerton, CA) (15) was used as substrate instead of glass. Standard nucleotide CE-phosphoramidites were used in the synthesis which was carried out on an adapted ABI 394 DNA synthesiser (Applied Biosystems). To make anti-B1, anti-B4 and anti-B5 arrays, a diamond-shaped mask with a 30 mm diagonal was used to deliver reagents to the polypropylene surface for in situ synthesis of the oligonucleotides. The mask was pressed against the polypropylene to create a cell. DNA synthesis reagents were introduced at the bottom of the cell and removed either from the top or from the bottom. After each nucleotide coupling the mask was displaced by a 1.5 mm step: the result was a template comprising overlapping footprints of the mask, with each smaller diamond shape representing an individual oligonucleotide. The longest oligonucleotides were 20mers, made along the centre line of the array. A second anti-B5 array was also made essentially as above except that a diamond-shaped mask with a 42 mm diagonal was used and the increment at each coupling step was 2 mm, thus producing 21mers along the center line of the array. The arrays were deprotected in 30% ammonia solution at 55°C for 16 h in a closed chamber (11).
Hybridisation conditions and analysis of the array images
Arrays were hybridised in 1 M NaCl, 10 mM Tris–HCl pH 7.4, 1 mM EDTA and 0.01% SDS at 22°C for 3 h. Approximately 50–60 fmol radiolabelled transcript was diluted in 12 ml of hybridisation solution and hybridisation was carried out in a hybridisation tube rolling at 3–4 r.p.m. The array was then washed in hybridisation solution briefly and dried in layers of Whatman paper. Autoradiography and analysis of hybridisation were carried out as described (16).
Theoretical folding of mRNAs
Folded structures of cyclin mRNAs with global minimum free energy were predicted using a commonly used folding program mfold (Genetics Computer Group, Madison, WI) according to Zuker’s latest energy minimisation calculations, which was provided on the University of Oxford Molecular Biology Data Center UNIX. The results were printed as ‘squiggles’ output.
Translation of endogenous mRNA in Xenopus egg extracts
Cytostatic factor (CSF)-arrested Xenopus egg extracts were prepared as described (17). A translation mix was prepared by mixing equal volumes of egg extract and nuclease-treated rabbit reticulocyte lysate (Ambion) containing 25 mM creatine phosphate, 2 mM KCl, 11 mM MgCl2, 1 mM EDTA, 3 mM l-leucine and l-valine and 2 mM all other amino acids except l-methionine and l-cysteine. The mixture was incubated at 22°C for 30 min in the presence of antisense oligonucleotides at the indicated final concentrations (normally 0.25–1 µM). After this preincubation 1.4 µCi (1 µl) 35S-labeled Pro-mix (>1000 Ci/mmol; Amersham) was added per 20 µl reaction volume to initiate labelling and incubated at 22°C for 90 min. An aliquot of 1 µl was saved for analysis of protein synthesis by SDS–PAGE. The remainder was divided into aliquots for use in immunoprecipitations.
Production of specific antisera against Xenopus cyclins
N-terminal fragments covering the first 108 amino acids of cyclins B1, B2 and B4 were subcloned into pET21b and expressed as C-terminally His-tagged proteins in Escherichia coli BL21(DE3). They were purified using Ni–agarose columns (Qiagen) and used to immunise rabbits to generate polyclonal antisera by standard methods. For cyclin B5 a 16 amino acid synthetic peptide corresponding to the N-terminus (WAAMRTGQMDNAVVKK) coupled to KLH was used for immunisation, as bacterial expression of the B5 N-terminus was poor.
Immunoprecipitation
Aliquots of the translation reactions were diluted 10-fold with immunoprecipitation buffer (50 mM Tris pH 8.0, 150 mM NaCl, 50 mM NaF, 5 mM EDTA, 0.1% NP40, 1 mM PMSF, 1 mM DTT, 2 mM benzamidine, 2 µg/ml each leupeptin and aprotinin). Polyclonal antisera were added at a final dilution of 1/100. After rotation at 4°C for 2 h the immunoprecipitates were collected using Affiprep protein A beads (Bio-Rad), washed five times in immunoprecipitation buffer and analysed by SDS–PAGE and autoradiography using a PhosphorImager.
Microinjection of Xenopus oocytes
Oocytes were suspended in OR2 medium (82.5 mM NaCl, 2 mM KCl, 1 mM MgCl2, 5 mM K–HEPES pH 7.5) (18) and treated with 1 mg/ml collagenase (Boehringer-Mannheim) for 2 h, washed repeatedly, after which stage VI oocytes were manually sorted and left at 18°C in modified Barth medium (19) for 12–24 h. They were induced to mature with 5 µg/ml progesterone (Sigma) at 22°C. Antisense oligonucleotides were dissolved in water and 50 nl was injected either 1–2 h after adding the progesterone or when the white spot had formed. For each experiment 20 oocytes were injected. After 1 h incubation or at the indicated times they were frozen in dry ice and stored at –70°C.
RNA extraction and RNase protection assay
RNA was extracted from samples of 20 oocytes following the proteinase K/phenol method described in section 7.16 of Sambrook et al. (20). An aliquot of 5 µg of total RNA was used for RNase protection assays. Antisense RNA probes of N-terminal fragments of different cyclins were transcribed from pGEM vectors using T7 polymerase and α-32P-labeled CTP (Amersham). RNase protection assays were performed using an Ambion RPAII Kit following the supplier’s protocol. The protected fragments were analysed by gel electrophoresis on an 8 M urea, 5% acrylamide gel and autoradiography.
Immunoblotting
Cell-free extracts of Xenopus oocytes were prepared by crushing 10 oocytes in 100 µl of EB buffer (80 mM sodium β-glycerophosphate, 20 mM EGTA, 25 mM NaF), followed by centrifugation for 10 min in an Eppendorf centrifuge. The clear portion of the supernatants was precipitated in 3 vol acetone, harvested and dissolved in SDS sample buffer. An aliquot corresponding to one oocyte was analysed by SDS–PAGE. The lanes were electrophoretically transferred to nitrocellullose and immunoblotted with the indicated affinity-purified antisera, which were generally used at a final dilution of 1:50, or 1 µg/ml of purified monoclonal antibodies. The bound antibodies were detected with the appropriate HRP-coupled secondary antibody and the Amersham-Pharmacia ECL kit.
RESULTS
RNase H mapping of the cyclin mRNAs reveals non-uniform accessibility to oligonucleotides
To identify the approximate positions in the mRNAs for cyclins B1, B4 and B5 that were accessible to intramolecular hybridisation 5′-end-labelled transcripts corresponding to each of the three mRNAs were incubated for 20 min with RNase H in the presence of increasing concentrations of a random 12mer oligonucleotide library (see the diagram in Fig. 1A) and analysed by PAGE (7). Each reaction contained ∼3 × 109 molecules of transcript. Figure 1B shows that addition of 500 pmol 12mers per reaction (which represents a ratio of mRNA to each individual 12mer of 166) resulted in cleavage of most of the full-length transcripts into discrete fragments. Each transcript gave a different and characteristic cleavage pattern, but in all three cases the accessible sites included some close to the first AUG codon (Fig. 1B). We therefore decided to scan the first 120 nt of each transcript using oligonucleotide arrays to precisely identify the sequences and the lengths of the optimally binding antisense oligonucleotides.
Figure 1.

Identification of accessible regions on the cyclin transcripts with RNase H/dN12 (A) Schematic representation of RNase H digestion of end-labelled mRNA. (B) Autoradiographs showing gel analysis of the cleaved products of 5′-end-labelled mRNA with RNase H at various dN12 concentrations. The transcripts were titrated against various concentrations of the oligomer library, ranging from 10 to 500 pmol/reaction (lane 0, native transcript; lane 1, plus RNase H; lane 2, 10 pmol; lane 3, 20 pmol; lane 4, 50 pmol; lane 5, 100 pmol; lane 6, 250 pmol; lane 7, 500 pmol). Each reaction contained ∼3 × 109 transcript molecules. The dN12 library contains 1.67 × 107 different 12mers. There are ∼1.8 × 107 molecules of each oligonucleotide in 500 pmol, which resulted in almost complete cutting of the full-length transcripts. How can this number of oligonucleotides mediate cleavage of 3 × 109 target molecules? Since regions of complementarity between RNA and an oligonucleotide of only 4 nt can recruit RNase H activity (25), the oligonucleotides in the library that were only partially complementary to the accessible sites would also be expected to participate in RNase H-mediated cutting of the transcript. In addition, each oligonucleotide can participate more than once in successive reactions.
Oligonucleotide arrays precisely locate regions of high and low accessibility to oligonucleotides in mRNA
The oligonucleotide arrays started at the first AUG of the three B-type cyclin mRNAs, ‘walking’ along the following 119 residues one residue at a time in lengths up to 20 nt. These arrays contained all possible complementary oligonucleotides corresponding to the mRNA sequence, covalently bound via their 3′-end to the polypropylene substrate. The arrays were fabricated by adding precursors via a diamond-shaped mask that was moved along 1.5 mm after each reaction so that, as shown in the cartoon in Figure 2, the oligonucleotides were 20 residues long at the centre of the array.
Figure 2.

The effects on binding capacity of sequences and lengths of antisense oligonculeotides determined on scanning arrays. (A) Schematic representation of a scanning array. The sequences given in the cartoon correspond to cyclin B5. (B) Each of the three panels shows an image obtained after hybridisation of the radiolabelled transcripts to the corresponding scanning array. The histograms, obtained with the program xvseq, represent binding of the 15mer oligonucleotides to the transcript on the array. Histograms corresponding to only half of the arrays are shown. The sequences of the antisense oligonucleotides used in various assays are shown on the histograms and underlined in the 120 nt sequence of the cyclin mRNAs.
Each of the three arrays was annealed with its respective radiolabelled transcript as described in Materials and Methods. The autoradiographs of the hybridised arrays showed several chevron-shaped regions of strong annealing to the mRNAs of all three cyclins (Fig. 2). Remarkably, in some cases, such as oligonucleotide B4-1, strong annealing was largely confined to a single chevron, so that the starting position of the oligonucleotide appears to be more important than its length (since annealing was still quite strong for the 10mer starting in the right place). Analysis of the images revealed the sequences and lengths of the oligonucleotides that bound strongly to the transcripts (Fig. 2). Sets of four 15mer oligonucleotides were picked from each hybridisation; each set comprised three oligonucleotides that showed strong binding and one from a region that showed very poor binding of the labelled transcript (Fig. 2 and Table 1).
Table 1. A list of antisense oligonucleotides.
| Name |
Length |
Sequence (5′) |
Bindinga |
Inhibition (at 1 µM) |
Nucleotide content (a, g, c, t) |
| B1-1 | 15 | gac tcg tag cga cat | 7 | no | 4, 4, 4, 3 |
| B1-2 | 15 | gtt ttc tgc att tgc | 389 | yes | 1, 3, 3, 8 |
| B1-3 | 15 | ttt tca cat tgt ttt | 222 | yes | 2, 1, 2, 10 |
| B1-4 | 15 | cct ctt tcc agc caa | 128 | yes | 3, 1, 7, 4 |
| B4-1 | 15 | ctc ctt gtc ata atc | 341 | yesb | 3, 1, 5, 6 |
| B4-2 | 15 | aca ttc tct gca ttg | 217 | yes | 3, 2, 4, 6 |
| B4-3 | 15 | agc ttt ctt tcc agc | 335 | yes | 2, 2, 5, 6 |
| B4-4 | 15 | tag gct ttg gta cag | 3 | yesb | 3, 5, 2, 5 |
| B5-1c | 15 | agc cca agt agc cat | 2 | no | 5, 3, 5, 2 |
| B5-2c | 15 | tcc atc tgt cct gta | 376 | yes | 2, 2, 5, 6 |
| B5-3c | 15 | cac tcc acc cac gac | 51 | yes | 4, 1, 9, 1 |
| B5-4c | 15 | gtc cat gta ctt tca | 110 | yes | 3, 2, 4, 6 |
| B5-a | 21 | tgt cca tgt act ttc act ttg | 288 | yes | 3, 3, 5, 10 |
| B5-b | 16 | caa gta gcc ata ctc c | 2 | no | 5, 2, 6, 3 |
| B5-c | 21 | gtc cat gta ctt tca ctt tgg | 319 | yes | 3, 4, 5, 9 |
| B5-d | 21 | tac gca tag cag ccc aag tag | 3 | no | 7, 5, 6, 3 |
| B5-e | 21 | ttg tcc atc tgt cct gta cgc | 497 | yes | 1, 4, 7, 8 |
| B5-f | 18 | gtc cat gta ctt tca ctt | 221 | yes | 3, 2, 5, 8 |
| B5-g | 21 | ctg cca cca ctc cac cca cga | 81 | yes | 5, 2, 12, 2 |
| B5-h | 18 | cag ccc aag tag cca tac | 4 | no | 6, 3, 7, 2 |
| B5-i | 18 | agc agc cca agt agc cat | 4 | no | 6, 4, 6, 2 |
| B5-j | 17 | tgt acg cat agc agc cc | 3 | no | 4, 4, 6, 3 |
| B5-k | 20 | ttg tcc atc tgt cct gta cg | 463 | yes | 2, 4, 6, 8 |
| B5-l | 18 | ctg cat tgt cca tct gtc | 202 | yes | 2, 3, 6, 7 |
| B5-m | 16 | ctg cat tgt cca tct g | 125 | yes | 2, 3, 5, 6 |
| B5-n | 18 | act ttc act ttg gct gcc | 52 | yes | 2, 3, 6, 7 |
| B5-o | 16 | gtc cat gta ctt tca c | 127 | yes | 3, 2, 5, 6 |
aArbitrary units.
bBoth these oligonucleotides inhibited only one of the two bands (see text for details).
cOligonucleotides used in microinjection.
Secondary structure analysis of the cyclin mRNAs does not predict accessible regions
The regions of strong hybridisation did not obviously correlate with sequence composition and we presumed that the most likely explanation for the surprisingly specific patterns of annealing lay in the secondary structure of the mRNA or the oligonucleotides. To test the possibility that accessible regions could be differentiated from inaccessible regions on theoretically predicted RNA folds we used the computer program mfold to predict structures of the three cyclin transcripts with global minimum free energy. However, analysis of the binding sites of the various antisense oligonucleotides on these structures, denoted by the bold lines in Figure 3, revealed no clear structural features distinguishing accessible from inaccessible sites. The best oligonucleotides do not, for example, correspond to the positions of loops in the predicted structures.
Figure 3.

Folded structures of the three cyclin mRNAs obtained using mfold. Only the relevant sections of the folds are shown. The sequences analysed on arrays are marked (start and end are indicated). The binding sites of the oligonucleotides are marked with bold lines on each structure. The grey line in the B1 mRNA fold represents overlap between oligonucleotides B1-2 and B1-3.
Array-selected oligonucleotides inhibit translation of their cognate targets in Xenopus egg extracts
To compare the efficiency of the various oligonucleotides as antisense reagents we first tested their effects on translation of endogenous mRNAs in frog egg extracts. The oligonucleotides were added to a 1:1 mixture of frog egg extracts and nuclease-treated rabbit reticulocyte lysate, which boosted translation. After 30 min at 20°C [35S]methionine was added to assay protein synthesis. After 90 min further incubation the reactions were analysed by SDS–PAGE and autoradiography. Figure 4A shows that some oligonucleotides, for example B1-4 and B4-3, showed quite strong inhibition of total protein synthesis. This was usually due to impurities in the samples and could be eliminated in most cases by further purification of the oligonucleotide (see Materials and Methods). Even so, many of the most highly purified oligonucleotides caused the disappearance of high molecular weight translation products (see asterisk in Fig. 4A). To test the degree of inhibition of synthesis of the targeted cyclins, specific polyclonal antisera were used to prepare immunoprecipitates that were also analysed by SDS–PAGE and autoradiography. Figure 4B shows that the oligonucleotides designed to ablate cyclins B1 and B5 strongly reduced synthesis of the targeted cyclins, whereas those with weak binding to the array (B1-1 and B5-1) had no or little effect. The antiserum against cyclin B4 immunoprecipitated two bands and oligonucleotides B4-2 and B4-3, both of which showed strong binding on the array, ablated both of these translation products. The third strongly binding oligonucleotide, B4-1, inhibited the synthesis of only the slower migrating band, possibly revealing the existence of two related but different versions of cyclin B4 mRNA in this animal. Unexpectedly, oligonucleotide B4-4, which showed weak hybridisation on the array, gave a similar result to B4-1 and ablated the slower but not the faster migrating band in the cyclin B4 immunoprecipitate.
Figure 4.

Effects of oligonucleotides on cell-free protein synthesis in frog egg extract. (A) The pattern of radioactive proteins displayed on SDS–PAGE after addition of 1 µM of the indicated oligonucleotides. The asterisk indicates an unidentified high molecular weight translation product whose synthesis is particularly sensitive to non-specific inhibition by the added DNA. (B) The radiolabelled immunoprecipitates obtained with the indicated antisera in response to the various oligonucleotides.
The immunoprecipitates also tested the effect of the antisense oligonucleotides on the closely related cyclins that were not their intended target. Some of the oligonucleotides did ablate non-targeted cyclins. For example, the anti-B1 oligonucleotides B1-2 and B1-4 partially inhibited the synthesis of cyclin B4 and the anti-B4 oligonucleotides B4-2 and B4-3 had significant inhibitory effect on cyclin B1 (Fig. 4B). Such non-specificity was due to close matching of the two sequences. Thus, oligonucleotide B1-2 contains two stretches of 8 nt complementary to cyclin B4 mRNA (one within the binding site of B4-2 and the other further downstream). Oligonucleotide B1-4 contained a stretch of 10 nt complementary to cyclin B4, lying within the binding site of B4-3. Both of these oligonucleotides significantly inhibited the translation of cyclin B4 mRNA, with B1-2 having a stronger effect than B1-4. Conversely, oligonucleotides B4-2 and B4-3 inhibited cyclin B1 synthesis, due to their extensive sequence similarity.
Correlation between hybridisation intensities and antisense efficiency for different cyclin B5 oligonucleotides
We made a more detailed analysis of the anti-cyclin B5 oligonucleotides, selecting a further 15 sequences of varying length and hybridisation intensity to assay for their antisense activities in egg extracts. These oligonucleotides were chosen from a fresh array that covered the sequence corresponding to the three areas showing major hybridisation in the first experiment. Figure 5A shows that the hybridisation results were very similar to those of the anti-B5 array shown in Figure 2. Each of the indicated antisense oligonucleotides was assayed as described above at a final concentration of 1 µM. Figure 5B shows that several oligonucleotides (a, c, e–g and k–o) strongly inhibited synthesis of cyclin B5. All these sequences gave strong hybridisation intensities and none of them showed serious inhibition of general protein synthesis, although we regularly noticed that the ‘best’ antisense oligonucleotides (e.g. e and k) tended to cause some inhibition of the synthesis of higher molecular weight proteins.
Figure 5.

(A) Designation, location and hybridisation intensities of the oligonucleotides tested for their effects on translation in (B), which shows both the total pattern of protein synthesis and the specific synthesis of B5 using immunoprecipitation and autoradiography. (C) Titration of selected oligonucleotides from 2 to 0.125 µM in five 2-fold dilution steps as indicated by the wedges. Note that oligonucleotides c, f and o cover the same region of the mRNA, but differ in length and hybridisation intensity as indicated. Oligonucleotide d comes from a poorly scoring region of the array.
We made titrations with five of the oligonucleotides to determine the quantitative relationship between heteroduplex yield, as measured on the array, and their potency as specific inhibitors of cyclin B5 translation. Figure 5C shows that even at the highest concentration the ‘bad’ oligonucleotide, d, hardly inhibited synthesis of cyclin B5, whereas all of the other oligonucleotides tested completely inhibited cyclin B5 translation at a concentration of 2 µM. At this concentration, however, all the oligonucleotides, except for o, showed considerable general inhibition of protein translation (data not shown). As the concentration of the oligonucleotides was lowered, translation of cyclin B5 was inhibited less, but oligonucleotide c, which was the most strongly annealing in this set, still gave almost complete ablation of cyclin B5 at 0.5 µM. Oligonucleotides c, f and o share the same 3′-end, but are 21, 18 and 16 nt long, respectively. Thus, the ability of the antisense oligonucleotides to ablate cyclin B5 followed the order of their intensities of hybridisation on the scanning array.
B5 antisense activity in intact Xenopus oocytes correlates well with the predictions from the arrays
To investigate how well hybridisation on the arrays correlated with antisense activity in living cells we injected four 15mer anti-cyclin B5 oligonucleotides (Table 1) into Xenopus oocytes and measured cleavage of B5 mRNA by RNase protection assay as a more direct assay of antisense activity. All the oligonucleotides caused scission of the B5 mRNA to varying degrees, giving rise to smaller protected bands that correspond to the RNase H-cleaved fragments (Fig. 6). Oligonucleotide B5-2 was the most effective, giving ∼92% cutting at even the lowest input, while oligonucleotide B5-1, which gave a weaker signal on the array, gave ∼70% cutting at the highest concentration. The mRNA of cyclin B3, which served as a control, was unaffected in all cases. There was thus a good correlation between the order of intensity of annealing of each oligonucleotide on the array and their effectiveness at promoting the cutting of cyclin B5 mRNA.
Figure 6.

Cleavage of cyclin B5 mRNA by antisense 15mers injected into intact Xenopus oocytes. (A) The location of four antisense oligonucleotides on the ∼350 nt probe and approximate sizes of the 3′-end cut products (drawings only roughly to scale). (B) The RNase protection map for cyclin B5 mRNA in response to antisense-mediated cleavage produced by 25, 50 and 100 ng each oligonucleotide as indicated by the wedges. Cyclin B3 was used as a negative control and the products of cleavage are indicated on the right side of the figure. The additional fainter bands seen in the B5-3 and B5-4 lanes possibly represent further degradation products of the B5 mRNA after cleavage with antisense oligonucleotides, since the oligonucleotides do not have sufficient complementarity in the neighbouring regions (data not shown) for secondary binding to produce faint bands of these sizes.
Antisense oligonucleotide B1-3 cleaves cyclin B1 mRNA and leads to disappearance of the protein in Xenopus oocytes
As a further test of whether antisense oligonucleotides chosen from the scanning array were effective in living cells we injected 100 ng oligonucleotide B1-3 into progesterone-treated Xenopus oocytes at the time of GVBD and monitored cyclin B1 mRNA cleavage 15, 30, 60 and 120 min later, using both RNase protection assays to measure the effects on mRNA and immunoblotting to assess the levels of residual cyclin B1. In the RNase protection assay, control oocytes showed two major bands that represent the full-length probe protected by the intact B1 mRNA (Fig. 7A). After injection of oligonucleotide B1-3 most cyclin B1 mRNA was cleaved within 30 min, giving rise to two shorter 5′ and a longer 3′ fragment of the protected probe. Immunoblotting with anti-cyclin B1 antibodies showed that an hour after injection of the oligonucleotide, cyclin B1 was essentially undetectable (Fig. 7B). This disappearance is due to the rapid turnover of cyclin B1 that occurs after entry into meiosis I (21). General inhibition of protein synthesis by cycloheximide (CHX) also caused the disappearance of cyclin B1, but in contrast to the oligonucleotide-injected oocytes, which showed no ill effects, the CHX-treated oocytes rapidly degenerated.
Figure 7.
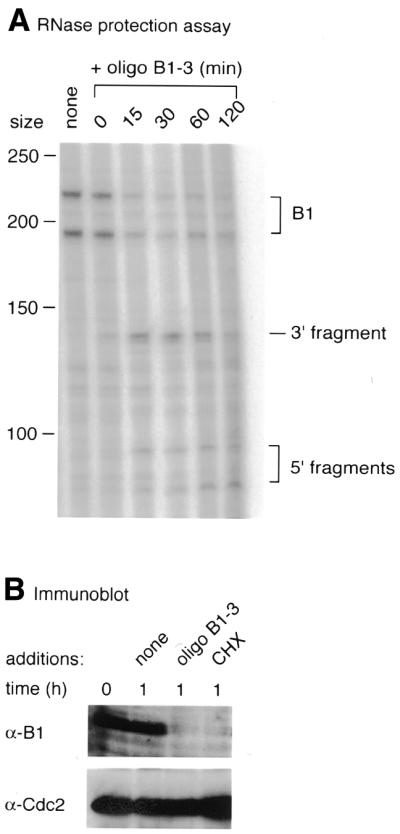
Time course of cleavage of cyclin B1 mRNA and effects on the levels of cyclin B1 protein in response to oligonucleotide B1-3. (A) Cleavage of cyclin B1 mRNA as measured by RNase protection mapping at the indicated times after injection of the DNA into progesterone-treated Xenopus oocytes. (B) Immunoblots of cyclin B1 and Cdc2 (loading control) in progesterone-treated oocytes at the time of GVBD (0) or 1 h later after injection of water or oligonucleotide solution as indicated or addition of 100 µg/ml cycloheximide (CHX) to the medium.
DISCUSSION
The aim of the studies described here was to test the validity of hybridisation of naked RNA to oligonucleotides for choosing antisense oligonucleotides that work in living cells. We roughly mapped the accessible positions of the mRNAs for Xenopus cyclins B1, B4 and B5 by mixing synthetic 5′-end-labelled transcripts with a random 12mer oligonucleotide library and RNase H, which located several accessible sites in the first 100 nt of all three mRNAs. Hybridisation of uniformly labelled transcripts to scanning arrays covering the first 120 nt of the transcripts revealed three or four regions available for heteroduplex formation in each RNA. It was not possible to use RNase H mapping alone to identify the precise sequence of the accessible site or to obtain quantitative measures of the relative accessibility of the various sites. And because it is necessary to use end-labelled transcripts for this mapping, RNase H-mediated cleavage of stronger upstream sites in the mRNA may mask accessible sites located further downstream. In contrast, hybridisation to arrays locates all accessible sites and provides precise, reproducible and quantitative measures of the accessibility of different regions of the transcript. Nevertheless, RNase H/dN12 mapping provides a useful first guide to identify accessible regions on an mRNA that can then be scanned in detail by an array, and we have found a good correlation between the methods for other targets (data not shown).
Binding affinities of oligonucleotides correlate with antisense efficiency
To test the correlation between binding of oligonucleotides on arrays and their ability to inhibit protein synthesis we first used a cell-free system derived from Xenopus egg extracts, in which it is a relatively simple matter to vary the concentration of added antisense oligonucleotides and the time of interaction before measuring the effects on translation of total and targeted mRNA populations.
One of the most serious practical problems we encountered was non-specific inhibition of protein synthesis due to impurities in the oligonucleotides, which could be minimised but never abolished by purification. For this reason it was important to keep the concentration of oligonucleotides as low as possible. We found that 0.5–1 µM gave good results in the case of the most active oligonucleotides, which represents an ∼1000-fold excess over the concentration of the cyclin mRNAs in Xenopus egg translation mix (22). Using this assay system we found a very good correlation between the hybridisation intensities of oligonucleotides on the scanning arrays and their capacity to ablate the targeted mRNA and thereby inhibit synthesis of the cognate protein.
A detailed analysis of the cyclin B5 5′-end reinforced the initial observations. Ten oligonucleotides that showed high hybridisation intensities on the array proved to be efficient antisense reagents, whereas five oligonucleotides that failed to score significantly on the array left cyclin B5 synthesis unaffected. Titrations of a series of oligonucleotides showed that their hybridisation efficiency closely reflected their antisense potency.
Similar results were obtained using intact Xenopus oocytes, although we found that higher concentrations (typically we tested 5, 10 and 20 µM) were required to give complete ablation of cyclin synthesis, probably because diffusion is required to deliver the reagent (cell-free reactions can be mixed efficiently). At these concentrations the purified oligonucleotides caused no detectable non-specific inhibition of protein synthesis in the intact oocytes, whose translation machinery is probably more robust than that of the cell-free system (data not shown). We compared the activity of four 15mers against B5 and confirmed that their relative hybridisation values reflected their potency to cut the cyclin B5 mRNA in oocytes, presumably by an RNase H-mediated mechanism (2). Oligonucleotide B5-2 was predicted by the array to be the best antisense reagent and it efficiently cleaved most of the cyclin B5 mRNA after 60 min at a concentration of 5 µM (25 ng/oocyte). Most published studies have used 100 ng of antisense oligonucleotide per oocyte (23,24).
These results suggest that the in vitro transcripts used have a similar, if not identical, secondary structure to the endogenous cyclin mRNAs in the cell. We also conclude that the intramolecular folding of mRNA forms the main obstacle to efficient hybridisation of antisense oligonucleotides. Oligonucleotide scanning arrays are thus useful tools for structural predictions and for selection of the most potent antisense reagents for a given mRNA. We have taken advantage of this method to re-investigate the question of the specific roles of the four B-type cyclins in oocyte maturation and the early cell cycles of fertilised Xenopus eggs and will report these findings elsewhere (H.Hochegger, A.Klotzbücher, J.Kirk, M.Howell, R.le Guellec, K.Fletcher, T.Duncan, M.Sohail and T.Hunt., submitted for publication). It is extremely important to choose highly active and specific oligonucleotides, especially in the case of the B-type cyclins, where there are four different mRNAs to ablate simultaneously and non-specific inhibition of protein synthesis seriously confuses the results.
Partial complementarity with accessible sites in mRNAs may trigger non-specific inhibition
What is the optimal length of an antisense nucleotide? Several considerations have to be taken into account to answer this question. An oligonucleotide should be long enough [∼15mer in human cells (5)] to have a unique complementary target sequence in the expressed gene pool and it would be tempting to suppose that longer sequences would become ever more specific. This appears to be a misleading assumption and most workers have used antisense oligonucleotides of ∼20 residues long. A 20mer has six unique 15 nt binding sites in the expressed gene pool (for an oligonucleotide of length n there are n – S + 1 Smers). Furthermore, since a 7 nt homology between an oligonucleotide and RNA is sufficient to recruit RNase H in human cells, a 20mer has 14 7 nt sequences, each one capable of mediating RNase H cutting. Increasing the length of the oligonucleotide thus carries the paradoxical risk of increasing the chance of non-specificity.
The ability of an antisense reagent to inhibit only the intended target is difficult to predict or even to find experimentally due to the lack of an available method to examine global effects. Since only short regions of homology between mRNA and an oligonuleotide, such as 4 bp in vitro (25) and 7 bp in cells (10), are sufficient to recruit RNase H activity, it is highly unlikely that any antisense oligonucleotide working by an RNase H-dependent mechanism would be completely specific for one mRNA.
We tested the incidence of non-specific inhibition by the antisense oligonucleotides in two ways. First, we investigated their effects on total protein translation by one-dimensional polyacrylamide gel electrophoresis. Despite the fact that resolution of proteins by this method was limited, it was possible to detect bands that were specifically inhibited. For some reason, non-specific inhibition was usually more serious with the most efficient antisense oligonucleotides. Many of the oligonucleotides inhibited the synthesis of several proteins and the pattern of inhibition varied among the various oligonucleotides, suggesting that this inhibition was likely to be sequence-specific. High molecular weight bands appeared to be most vulnerable to inhibition. We also encountered problems with impurities in the oligonucleotide preparations, but these could usually be overcome by careful purification of the nucleic acids. In general, the most efficient way to minimise these problems was to use as little oligonucleotide as possible.
Secondly, we examined the effects of oligonucleotides designed to ablate particular B-type cyclins on the synthesis of their close relatives. We found that cases of cross-reactivity between oligonucleotides and non-targeted cyclins was usually the result of partial sequence complementarity, typically stretches of 8–10 residues that were complementary to ‘accessible regions’ of their unintended targets. These data show how complicated it is to predict the specificity of oligonucleotides. If a 7mer or 8mer overlap of a given oligonucleotide is enough to cause RNase H-mediated cleavage, any oligonucleotide is likely to have a number of unintended targets in the cell. Even so, efficient ablation of the additional targets will only occur in cases where the structure of the mRNA is accessible in the complementary region under the conditions obtaining in cytoplasm.
These considerations stress that the design of potent antisense oligonucleotides is crucial to minimise non-specificity. The experiments with frog oocytes described here well illustrate this point, for we never observed non-specific inhibition of oocyte maturation using our carefully characterised and purified oligonucleotides, which were nevertheless very effective in inhibiting cyclin synthesis. Such non-specific inhibition was a serious problem in previous studies in which ∼80% of the oocytes failed to mature, even when injected with control DNA (26). Nevertheless, non-specific ablation of mRNAs can never be completely eliminated and this study reinforces the importance of control experiments in the interpretation of biological effects caused by antisense oligonucleotides.
Relationship of target site and oligonucleotide sequences with antisense activity
Quite often the translation initiation site is targeted with antisense reagents because it is believed to be relatively free from secondary structure to allow initiation of protein synthesis (27). However, data obtained for at least two cyclins, B1 and B5, do not support this assumption. For both these cyclins, sequences around the AUG start site were found to be largely inaccessible, as shown by the hybridisation results (Fig. 2) and also by the lack of any detectable antisense activity of oligonucleotides B1-1 and B5-1, both of which covered the AUG start codon. Antisense effect of an oligonucleotide covering the start codon of cyclin B4 was not investigated but, due to the lack of any detectable signal in this region (Fig. 2), this was also likely to be ineffective.
In order to find any noticeable features that may be present in effective antisense oligonucleotides, such as the presence of sequence motifs (28) or abundance of certain nucleotides (29,30), we used a set of nine oligonucleotides (B1-2, B1-3, B1-4, B4-2, B4-3, B4-4, B5-a, B5-e and B5-g) that covered a total of 192 nt sequence. Seven of these were non-overlapping, but two, B1-2 and B1-3, have a 5 nt overlap (Table 1). None of these (or any other oligonucleotides used here) had the motif TCCC described by Tu et al. (28), but an unrelated 4 nt sequence, TTTC, occurred six times and two other 4 residue sequences, TCCA and CTTT, five times. Matveeva et al. (31) suggested a positive correlation between the presence or absence of certain motifs (CCAC, TCCC, ACTC, GCCA and CTCT) and the antisense efficiency of the oligonucleotides. They also suggested that the likelihood of activity of oligonucleotides was sequence motif content-dependent. However, we were unable to detect such a correlation in our set of oligonucleotides. For example, the motif ACTC was present in one ‘good’ oligonucleotide (B5-g) but also in two ‘bad’ ones (B1-1 and B5-1). The motif GCCA was present in one good oligonucleotide (B1-4) but also in three bad ones (B5-1, B5-b and B5-h). The oligonucleotide B5-g has the motif CCAC occurring four times and ACTC once, but this accumulation of positive motifs did not make it the best antisense oligonucleotide in our tests. It is also important to note that Matveeva et al. (31) did not find the TCCC motif in a significant number of efficient oligonucleotides.
The ‘good’ antisense oligonucleotides were also not particularly high in either G or C nucleotides, as has previously been reported (29,30), but most had a relatively high content of T residues. Further empirical studies are required to settle these issues.
mfold was unable to predict optimal binding sites
Computer-predicted folded structures are often used as a starting point to design antisense oligonucleotides (32–34). A general strategy is to target single-stranded regions in the predicted structures. However, computational methods to predict secondary structure of mRNAs are not sufficiently developed and it is not clear that single-stranded regions are indeed more accessible than stem–loops (35). We analysed the structure at the binding sites of 12 15mer oligonucleotides targeted to the three frog B-type cyclins using the program mfold, but were unable to discern a pattern that could allow a reliable distinction to be made between accessible and inaccessible sites, as previously concluded by others (7,8).
Acknowledgments
ACKNOWLEDGEMENTS
The authors would like to thank Lydie Baudouin, Stephan Geley and Hiro Mahbubani for their help and advice and the ICRF Biological Resource Unit for providing rabbit polyclonal antisera. M.S. was funded by the Medical Research Council, UK and H.H. by the Boehringer Ingelheim Fonds.
References
- 1.Walder R.Y. and Walder,J.A. (1988) Role of RNase H in hybrid-arrested translation by antisense oligonucleotides. Proc. Natl Acad. Sci. USA, 85, 5011–5015. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 2.Minshull J. and Hunt,T. (1986) The use of single-stranded DNA and RNase H to promote quantitative ‘hybrid arrest of translation’ of mRNA/DNA hybrids in reticulocyte lysate cell-free translations. Nucleic Acids Res., 14, 6433–6451. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 3.Zamecnik P.C., Goodchild,J., Taguchi,Y. and Sarin,P.S. (1986) Inhibition of replication and expression of human T-cell lymphotropic virus type III in cultured cells by exogenous synthetic oligonucleotides complementary to viral RNA. Proc. Natl Acad. Sci. USA, 83, 4143–4146. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 4.Branch A.D. (1998) Antisense drug discovery: can cell-free screens speed the process? Antisense Nucleic Acid Drug Dev., 8, 249–254. [DOI] [PubMed] [Google Scholar]
- 5.Woolf T.M., Melton,D.A. and Jennings,C.G. (1992) Specificity of antisense oligonucleotides in vivo. Proc. Natl Acad. Sci. USA, 89, 7305–7309. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 6.Bramlage B., Luzi,E. and Eckstein,F. (1998) Designing ribozymes for the inhibition of gene expression. Trends Biotechnol., 16, 434–438. [DOI] [PubMed] [Google Scholar]
- 7.Ho S.P., Bao,Y., Lesher,T., Malhotra,R., Ma,L.Y., Fluharty,S.J. and Sakai,R.R. (1998) Mapping of RNA accessible sites for antisense experiments with oligonucleotide libraries. Nat. Biotechnol., 16, 59–63. [DOI] [PubMed] [Google Scholar]
- 8.Milner N., Mir,K.U. and Southern,E.M. (1997) Selecting effective antisense reagents on combinatorial oligonucleotide arrays. Nat. Biotechnol., 15, 537–541. [DOI] [PubMed] [Google Scholar]
- 9.Patzel V. and Sczakiel,G. (1998) Theoretical design of antisense RNA structures substantially improves annealing kinetics and efficacy in human cells. Nat. Biotechnol., 16, 64–68. [DOI] [PubMed] [Google Scholar]
- 10.Wagner R.W., Matteucci,M.D., Grant,D., Huang,T. and Froehler,B.C. (1996) Potent and selective inhibition of gene expression by an antisense heptanucleotide. Nat. Biotechnol., 14, 840–844. [DOI] [PubMed] [Google Scholar]
- 11.Southern E.M., Case-Green,S.C., Elder,J.K., Johnson,M., Mir,K.U., Wang,L. and Williams,J.C. (1994) Arrays of complementary oligonucleotides for analysing the hybridisation behaviour of nucleic acids. Nucleic Acids Res., 22, 1368–1373. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 12.Minshull J. and Hunt,T. (1992) Antisense ablation of mRNA in frog and rabbit cell-free systems. In Murray,J.A.H. (ed.) Antisense RNA and DNA. Wiley-Liss, New York, NY, pp. 195–212.
- 13.Sohail M. and Southern,E.M. (2001) Hybridization of antisense reagents to RNA. Curr. Opin. Mol. Ther., 2, 264–271. [PubMed] [Google Scholar]
- 14.Sohail M. and Southern,E.M. (2000) Antisense arrays. Mol. Cell Biol. Res. Commun., 3, 67–72. [DOI] [PubMed] [Google Scholar]
- 15.Matson R.S., Rampal,J.B. and Coassin,P.J. (1994) Biopolymer synthesis on polypropylene supports. I. Oligonucleotides. Anal. Biochem., 217, 306–310. [Erratum, Anal. Biochem. (1994) 220, 225] [DOI] [PubMed] [Google Scholar]
- 16.Sohail M. and Southern,E.M. (2001) Using oligonucleotide scanning arrays to find effective antisense reagents. In Pampal,J.B. (ed.) Methods in Molecular Biology—Oligonucleotide Array Methods and Protocols. Humana Press, Totowa, NJ, in press. [DOI] [PubMed]
- 17.Murray A.W. (1991) Cell cycle extracts. In Kay,B.K. and Peng,H.B. (eds) Xenopus laevis: Practical Uses in Cell and Molecular Biology. Academic Press, San Diego, CA, Vol. 36, pp. 573–597.
- 18.Eppig J.J. and Dumont,J.N. (1976) Defined nutrient medium for the in vitro maintenance of Xenopus laevis oocytes. In Vitro, 12, 418–427. [DOI] [PubMed]
- 19.Heasman J., Holwill,S. and Wylie,C.C. (1991) Fertilization of cultured Xenopus oocytes and use in studies of maternally inherited molecules. In Kay,B.K. and Peng,H.B. (eds) Xenopus laevis: Practical Uses in Cell and Molecular Biology. Academic Press, San Diego, CA, Vol. 36, pp. 213–230. [DOI] [PubMed]
- 20.Sambrook J., Fritsch,E.F. and Maniatis,T. (1989) Molecular Cloning: A Laboratory Manual, 2nd Edn. Cold Spring Harbor Laboratory Press, Cold Spring Harbor, NY.
- 21.Thibier C., De Smedt,V., Poulhe,R., Huchon,D., Jessus,C. and Ozon,R. (1997) In vivo regulation of cytostatic activity in Xenopus metaphase II-arrested oocytes. Dev. Biol., 185, 55–66. [DOI] [PubMed] [Google Scholar]
- 22.Kobayashi H., Stewart,E., Poon,R.Y. and Hunt,T. (1994) Cyclin A and cyclin B dissociate from p34cdc2 with half-times of 4 and 15 h, respectively, regardless of the phase of the cell cycle. J. Biol. Chem., 269, 29153–29160. [PubMed] [Google Scholar]
- 23.Sagata N., Oskarsson,M., Copeland,T., Brumbaugh,J. and Vande Woude,G.F. (1988) Function of c-mos proto-oncogene product in meiotic maturation in Xenopus oocytes. Nature, 335, 519–525. [DOI] [PubMed] [Google Scholar]
- 24.Sheets M.D., Wu,M. and Wickens,M. (1995) Polyadenylation of c-mos mRNA as a control point in Xenopus meiotic maturation. Nature, 374, 511–516. [DOI] [PubMed] [Google Scholar]
- 25.Donis-Keller H. (1979) Site specific enzymatic cleavage of RNA. Nucleic Acids Res., 7, 179–192. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 26.Minshull J., Murray,A., Colman,A. and Hunt,T. (1991) Xenopus oocyte maturation does not require new cyclin synthesis. J. Cell Biol., 114, 767–772. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 27.Kozak M. (1991) Structural features in eukaryotic mRNAs that modulate the initiation of translation. J. Biol. Chem., 266, 19867–19870. [PubMed] [Google Scholar]
- 28.Tu G.C., Cao,Q.N., Zhou,F. and Israel,Y. (1998) Tetranucleotide GGGA motif in primary RNA transcripts. Novel target site for antisense design. J. Biol. Chem., 273, 25125–25131. [DOI] [PubMed] [Google Scholar]
- 29.Agrawal S. (1999) Importance of nucleotide sequence and chemical modifications of antisense oligonucleotides. Biochim. Biophys. Acta, 1489, 53–68. [DOI] [PubMed] [Google Scholar]
- 30.Smetsers T.F., Boezeman,J.B. and Mensink,E.J. (1996) Bias in nucleotide composition of antisense oligonucleotides. Antisense Nucleic Acid Drug Dev., 6, 63–67. [DOI] [PubMed] [Google Scholar]
- 31.Matveeva O.V., Tsodikov,A.D., Giddings,M., Freier,S.M., Wyatt,J.R., Spiridonov,A.N., Shabalina,S.A., Gesteland,R.F. and Atkins,J.F. (2000) Identification of sequence motifs in oligonucleotides whose presence is correlated with antisense activity. Nucleic Acids Res., 28, 2862–2865. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 32.Lehmann M.J., Patzel,V. and Sczakiel,G. (2000) Theoretical design of antisense genes with statistically increased efficacy. Nucleic Acids Res., 28, 2597–2604. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 33.Vickers T.A., Wyatt,J.R. and Freier,S.M. (2000) Effects of RNA secondary structure on cellular antisense activity. Nucleic Acids Res., 28, 1340–1347. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 34.Patzel V., Steidl,U., Kronenwett,R., Haas,R. and Sczakiel,G. (1999) A theoretical approach to select effective antisense oligodeoxyribonucleotides at high statistical probability. Nucleic Acids Res., 27, 4328–4334. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 35.Mir K.U. and Southern,E.M. (1999) Determining the influence of structure on hybridization using oligonucleotide arrays. Nat. Biotechnol., 17, 788–792. [DOI] [PubMed] [Google Scholar]