

Abstract
Growth of the yeast vacuolar protein-sorting mutant vps5Δ affected in the endosome-to-Golgi retromer complex was more sensitive to Mg2+-limiting conditions than was the growth of the wild-type (WT) strain. This sensitivity was enhanced at acidic pH. The vps5Δ strain was also sensitive to Al3+, known to inhibit Mg2+ uptake in yeast cells. In contrast, it was found to be resistant to Ni2+ and Co2+, two cytotoxic analogs of Mg2+. Resistance to Ni2+ did not seem to result from the alteration of plasma-membrane transport properties because mutant and WT cells displayed similar Ni2+ uptake. After plasma-membrane permeabilization, intracellular Ni2+ uptake in vps5Δ cells was 3-fold higher than in WT cells, which is consistent with the implication of the vacuole in the observed phenotypes. In reconstituted vacuolar vesicles prepared from vps5Δ, the rates of H+ exchange with Ni2+, Co2+, and Mg2+ were increased (relative to WT) by 170%, 130%, and 50%, respectively. The rates of H+ exchange with Ca2+, Cd2+, and K+ were similar in both strains, as were α-mannosidase and H+-ATPase activities, and SDS/PAGE patterns of vacuolar proteins. Among 14 other vacuolar protein-sorting mutants tested, only the 8 mutants affected in the recycling of trans-Golgi network membrane proteins shared the same Ni2+ resistance phenotype as vps5Δ. It is proposed that a trans-Golgi network Mg2+/H+ exchanger, mislocalized to vps5Δ vacuole, could be responsible for the phenotypes observed in vivo and in vitro.
Protein sorting pathways from the trans-Golgi network (TGN) to the vacuole have been extensively studied through a large number of yeast mutants (reviewed in refs. 1 and 2). To date, two main intracellular routes have been identified, exemplified by the respective targeting pathways of carboxypeptidase Y (CPY) and alkaline phosphatase (ALP). CPY travels through a prevacuolar/endosomal compartment (PVC) and subsequently onto the vacuole, whereas ALP bypasses the PVC to reach the same organelle. Numerous dedicated proteins localized in transport vesicles or target membranes take part in this process, and the recycling of some of these proteins is important for the efficiency of some trafficking steps. In particular, a membrane-coat complex, designated as the retromer complex, is essential for the PVC-to-Golgi retrograde vesicular transport (3). The retromer complex assembles from two subcomplexes composed of Vps35p/Vps29p/Vps26p and Vps5p/Vps17p, respectively. The first subcomplex is thought to select cargo for retrieval, whereas the second would promote vesicle formation (3–5). Mutation of VPS5, a nonessential gene, causes the secretion of CPY (6, 7). Indeed, Vps5p is required for the PVC-to-Golgi retrieval of Vps10p, a membrane protein that ensures binding and Golgi-to-PVC transport of CPY. Consequently, Vps10p is misrouted to the vacuole membrane in vps5 mutants, whereas CPY accumulates in the Golgi and is then secreted.
In this study, a vps5Δ strain is shown to display new growth phenotypes toward Mg2+-limiting media or media containing the Mg2+ cytotoxic analogs Ni2+ and Co2+. Together with in vivo assays of Ni2+ uptake and in vitro assays of vacuolar C2+ (divalent cation)/H+ exchange, our data suggest that the phenotypes originate from an enhanced vacuolar compartmentalization of C2+ mediated by a Mg2+/H+ exchange mechanism.
Materials and Methods
Strains, Media, and Spot Assays.
The haploid Saccharomyces cerevisiae wild-type (WT) strain Hansen BY4741 (ref. 8; genotype: MATa his3-1 leu2-0 met15-0 ura3-0) was obtained from the American Type Culture Collection, and the vps mutants were obtained from Research Genetics (Huntsville, AL). Yeast strains were propagated in standard yeast nitrogen base (YNB) minimal medium (9). To screen heavy metal resistance phenotypes, medium was prepared from a YNB powder containing ammonium sulfate but no C2+ (Bio 101). The YNB powder was complemented with glucose and Ca2+, Cu2+, Fe2+, Mg2+, Mn2+, and Zn2+ up to standard concentrations (9), except for Mg2+, which was adjusted to 100 μM. A supplemental mixture (CSM, Bio 101) was added according to manufacturer's instructions, and agar was used as the gelling agent (14 g⋅liter−1). After autoclaving, heavy metals to which resistance was tested were added in the medium at the desired concentrations. Sensitivity to Mg2+-limiting conditions was studied on a similar YNB minimal medium, except that agar was replaced by a low-Mg2+-containing agarose (12 g⋅liter−1; Low EEO, Sigma). This medium did not allow yeast growth without Mg2+ supplementation (data not shown). Magnesium was added at the indicated concentrations after autoclaving. The pH of YNB minimal media was 4.3. When required, the pH was adjusted before autoclaving with HCl to pH 3.0, with 50 mM succinate/KOH to pH 4.3 or 5.6, or with 50 mM Hepes/KOH to pH 7.5.
Spot assays were performed with strains grown up to stationary phase in standard YNB minimal medium (OD600 = 6). Ten microliters of diluted cultures (OD600 = 0.02) was laid on plates containing the selective media, and growth phenotypes were documented daily with a video imaging system. A wide range of ion conditions was tested; only representative conditions were chosen for the figures presented below. For Ni2+ uptake or tonoplast isolation, strains were grown to mid-logarithmic phase (OD600 ≈ 3) in standard YNB minimal medium. Cells were harvested by centrifugation at 4,400 × g for 5 min. Pellets were washed at room temperature in water and resuspended in the buffers indicated below.
Vacuole Isolation, Biochemical Assays.
According to Roberts et al. (10), yeast cells were resuspended in Zymolyase-containing buffer to be converted to spheroplasts, and after osmotic lysis in the presence of a yeast protease inhibitor mixture (Sigma), vacuoles were purified by flotation on a discontinuous Ficoll gradient. After elimination of lipid particles (11), vacuoles were fragmented, and tonoplast vesicles were pelleted and resuspended in 2 mM Tris⋅HCl, pH 7.5/250 mM sorbitol/2 mM DTT/20% (vol/vol) glycerol and stored in liquid nitrogen. V-ATPase and α-mannosidase activities were measured as described by Lichko et al. (12) and Roberts et al. (10). SDS/PAGE of tonoplast proteins was performed according to Laemmli (13). Gels were silver-stained (14) and analyzed with imagemaster image analysis software (Amersham Pharmacia). Cell fractionation experiments using differential centrifugation to separate P150 and P13 membrane fractions from the cytosolic fraction (S150) were performed essentially as described (15). First, the purity of vacuolar membrane preparations was assessed from the ATPase activities of plasma membrane and mitochondria, according to Landolt-Marticorena et al. (16). Second, Western blotting was performed by using monoclonal antibodies (Molecular Probes) against Vph1p (dilution 1:2,000), Pep12p (1:600), Dpm1p (1:400), CoxIIIp (1:1,000), and Vps10p (1:400). Proteins were immunodetected by using a chemiluminescent assay (Aurora, ICN). Protein concentrations were determined by the Schaffner and Weissman method (17), using BSA as a standard.
C2+/H+Exchange in Vitro.
Tonoplast proteins were inserted into soybean liposomes essentially as
described (18), except for the following. The detergent octyl glucoside
was eliminated by using Sephadex G-50 minicolumns (Amersham Pharmacia)
(19). Reconstitution was performed at a lipid/protein ratio of 30
(wt/wt). The reconstitution buffer contained 5 mM EDTA
(Na+-free), adjusted to pH 5.5 with
1,3-bis[tris(hydroxymethyl)methylamino]propane and then adjusted
to pH 7.0 with NH4OH, 0.4 M sorbitol, and
glycerol to a final concentration of 20% (vol/vol). Reconstituted
tonoplast vesicles were diluted 200-fold in a stirred cuvette (1 ml)
containing a NH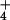 -free assay buffer (5 mM LiHepes, pH
7.5/0.4 M sorbitol) and the fluorescent permeant pH probe
9-amino-6-chloro-2-methoxyacridine (ACMA) at 1 μM. The
NH
-free assay buffer (5 mM LiHepes, pH
7.5/0.4 M sorbitol) and the fluorescent permeant pH probe
9-amino-6-chloro-2-methoxyacridine (ACMA) at 1 μM. The
NH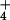 dilution resulted in an acid-loading of the
vesicles, caused by the outward diffusion of NH3
(20). Transmembrane ΔpH was monitored by the fluorescence quenching
of ACMA measured at 415/485 nm. Thereafter, quasi-infinite inward
gradients of the indicated C2+ were imposed by
adding C2+ to the outside.
C2+/H+ exchangers used
this gradient to generate divalent influxes coupled to
H+ efflux, estimated by the initial rate of
dissipation of the ACMA quenching (determined from the derivation of
the kinetics). This initial rate, named
VH+, was linear with the protein
concentration (data not shown) and is expressed in
%⋅min−1 per mg of protein.
dilution resulted in an acid-loading of the
vesicles, caused by the outward diffusion of NH3
(20). Transmembrane ΔpH was monitored by the fluorescence quenching
of ACMA measured at 415/485 nm. Thereafter, quasi-infinite inward
gradients of the indicated C2+ were imposed by
adding C2+ to the outside.
C2+/H+ exchangers used
this gradient to generate divalent influxes coupled to
H+ efflux, estimated by the initial rate of
dissipation of the ACMA quenching (determined from the derivation of
the kinetics). This initial rate, named
VH+, was linear with the protein
concentration (data not shown) and is expressed in
%⋅min−1 per mg of protein.
63Ni2+ Uptake.
The yeast pellet was resuspended (final OD600 = 4) in 5 mM Tris⋅succinate, pH 4.3/2% (wt/vol) glucose/50 mM KCl. This suspension (900 μl) was incubated for 30 min at 30°C under constant agitation (80 rpm). Nickel uptake was initiated by the addition of 63Ni (6.14 MBq⋅mmol−1 63NiCl2, DuPont) at 3.5 μM final concentration and 250 μM NiCl2. The uptake was stopped at different times by the filtration of 100-μl aliquots on 0.45-μm pore nitrocellulose membranes (Millipore). Filters were washed three times with 10 ml of 40 mM NiCl2; radioactivity retained on filters was measured by using a liquid scintillation analyzer. 63Ni2+ contents were expressed in mmol per mg−1 dry weight. 63Ni2+ uptake experiments on permeabilized yeast cells were performed at pH 6.9 as described (21, 22) and in the same conditions as above.
Results
On Mg2+ nonlimiting solid media, the vps5Δ and WT strains displayed a similar growth capacity at any pH tested (pH 3.0 to 7.5; Fig. 1A). In contrast, on Mg2+-limiting media, the growth capacity of both strains was impaired upon medium acidification. Importantly, vps5Δ exhibited a stronger sensitivity than WT to such acidification in low-Mg2+ conditions. This phenotype is called low-MgS in the rest of this paper.
Figure 1.

Growth phenotypes of vps5Δ and WT. (A) pH-dependent phenotypes in Mg2+-limiting media. Spot assays were recorded after 5-days (200 μM Mg2+, Mg2+-nonlimiting condition) or 10-days of growth (other assays, Mg2+-limiting conditions) on an agarose/YNB minimal medium at the indicated pH and Mg2+ concentrations. (B) Tolerance to cytotoxic heavy metals. Spot assays were recorded after 6-days of growth on an agar/YNB minimal medium (pH 4.3) containing 0.1 mM Mg2+ (nonlimiting), and the following concentrations of the indicated metals: 1 mM Al3+, 0.5 mM Co2+, 0.25 mM Ni2+, and 0.25 mM Cd2+. C, control plate.
The vps5Δ strain also showed contrasting phenotypes in response to heavy metals in nonlimiting Mg2+ conditions (Fig. 1B). Relative to WT, the vps5Δ strain was resistant to Ni2+ and to Co2+ (NiR and CoR phenotypes) but sensitive to Al3+ and Cd2+ (AlS and CdS phenotypes). The kinetics of 63Ni2+ uptake by vps5Δ and WT cells were similar (Fig. 2A). However, after plasma-membrane permeabilization (21, 22), the intracellular 63Ni2+ uptake by vps5Δ cells reached a level 3-fold higher after 6 hr (Fig. 2B). Tonoplast vesicles were prepared from Ficoll gradient-purified vacuoles as indicated in Materials and Methods. The isolation yields of tonoplast proteins per g fresh weight of the two strains were similar, as well as the activities of the tonoplast enzyme markers V-ATPase and α-mannosidase (Fig. 3A). Inhibition of the total ATP hydrolysis activity by bafilomycin was about 70%, whereas inhibition by oligomycin and vanadate was less than 10%, indicating minimal contamination by mitochondrial and plasma membranes, respectively. SDS/PAGE patterns of tonoplast proteins were indistinguishable (Fig. 3B). Fig. 3C shows Western blot analyses performed on vacuolar membrane preparations and subcellular P15, P150, and S150 fractions from WT and mutant by using monoclonal antibodies directed against membrane markers of the tonoplast (Vph1p), mitochondria (CoxIIIp), endoplasmic reticulum (Dpm1p), PVC (Pep12p), and TGN (Vps10p). These data indicate that the vacuolar preparations are highly enriched in vacuolar membrane proteins, which is in agreement with previous studies (11). Contaminations with endoplasmic reticulum and PVC were minor and comparable for both strains, whereas contaminations with mitochondria and TGN were not detectable.
Figure 2.

63Ni2+ uptake by vps5Δ and WT cells. (A) Control cells. (B) Cells with permeabilized plasma membrane. Values for WT (●) and vps5Δ (○) are presented as mean ± SE (n = 3). The uptake is expressed as mmol of Ni2+ per mg of dry weight.
Figure 3.

Biochemical properties of tonoplast isolated from vps5Δ and WT. (A) Isolation yield, hydrolytic activity of V-ATPase and α-mannosidase expressed in mg of protein per g of fresh weight, μmol of ATP hydrolyzed per hr per mg of protein, and nmol of nitrophenol per min per mg of protein, respectively. These activities were not significantly different between the two strains (Student's t tests, data not shown). (B) In SDS/PAGE patterns of tonoplast proteins, 62 bands were detected in both patterns with image analysis software. (C) Western blot analysis of vacuole membrane (Vac) and subcellular P15, P150, and S150 fractions from WT and vps5Δ strains. For a given antibody, equal amounts of fraction proteins were loaded. Monoclonal antibodies were directed against membrane markers of the tonoplast (Vph1p), PVC (Pep12p), endoplasmic reticulum (Dpm1p), mitochondria (CoxIIIp), and TGN (Vps10p).
Initial trials (not shown) showed that the filling kinetics of native
tonoplast vesicles with C2+ in exchange with
H+ were too fast to determine the initial rate of
the exchange (VH+). Therefore, the latter
was assayed on reconstituted vesicles to slow down the transport
reaction (23). We showed that a similar reconstitution of a plant
tonoplast Mg2+/H+
exchanger allowed for the recovery of the same affinity for
Mg2+ and sensitivity to inhibitors as at the
native membrane level (18). VH+ was
measured after an initial acid-loading step was performed as described
in Materials and Methods. The initial fluorescence quenching
of the pH probe ACMA could be totally dissipated by
NH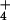 addition. This result ascertained the
instantaneous establishment of a stable transmembrane ΔpH upon
acid-loading (Fig. 4). In control
liposomes, the addition of Ni2+ (or other cations
used in this study) to the outside caused a negligible dissipation of
ΔpH, as compared with that observed with reconstituted tonoplast
vesicles (Fig. 4). Relative to WT, a 2.7-fold higher initial rate of
Ni2+/H+ exchange was
observed with reconstituted vesicles from vps5Δ.
addition. This result ascertained the
instantaneous establishment of a stable transmembrane ΔpH upon
acid-loading (Fig. 4). In control
liposomes, the addition of Ni2+ (or other cations
used in this study) to the outside caused a negligible dissipation of
ΔpH, as compared with that observed with reconstituted tonoplast
vesicles (Fig. 4). Relative to WT, a 2.7-fold higher initial rate of
Ni2+/H+ exchange was
observed with reconstituted vesicles from vps5Δ.
Figure 4.

Activity of Ni2+/H+ exchange in reconstituted
tonoplast vesicles from vps5Δ and WT.
Ammonium-containing vesicles were diluted 200-fold in the assay buffer
free of NH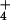 , causing vesicle acid-loading and instant
quenching of the permeant pH probe ACMA. Fluorescence recovery after
the addition of 0.5 mM Ni2+ to reconstituted tonoplast
vesicles, not observed on control liposomes, ascertained the
facilitated exchange activity. The final addition of
NH
, causing vesicle acid-loading and instant
quenching of the permeant pH probe ACMA. Fluorescence recovery after
the addition of 0.5 mM Ni2+ to reconstituted tonoplast
vesicles, not observed on control liposomes, ascertained the
facilitated exchange activity. The final addition of
NH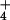 (15 mM) to the outside allowed the total
dissipation of the pH gradient. Solid and dotted lines represent the
mean ACMA fluorescence in arbitrary units (au) ± SE
(n = 5), respectively.
(15 mM) to the outside allowed the total
dissipation of the pH gradient. Solid and dotted lines represent the
mean ACMA fluorescence in arbitrary units (au) ± SE
(n = 5), respectively.
Selectivity of C2+/proton exchange of reconstituted tonoplast vesicles from vps5Δ and WT was compared (Fig. 5). In vps5Δ vesicles, VH+ for Mg2+, Co2+, or Ni2+ increased by 50%, 130%, and 170%, respectively, compared with WT. In contrast, VH+ with Ca2+, Cd2+, or K+ was similar for both strains.
Figure 5.

Selectivity of cation/proton exchange across reconstituted tonoplast vesicles from vps5Δ or WT. Cation concentration was 0.5 mM, except for Cd2+, which was 0.2 mM. Exchange rates (VH+) are presented as mean VH+ ± SE (n = 3), except for Mg2+ (n = 5). Exchange rates were significantly different between vps5Δ and WT for Mg2+, Co2+, and Ni2+, but not for Cd2+, Ca2+, and K+ (Student's t tests, data not shown).
To determine the kinetic parameters of the Mg2+/H+ exchange, VH+ was measured at various Mg2+ concentrations (Fig. 6A). Scatchard plots were linear (Fig. 6B), indicating that the facilitated exchange reaction could be fitted to the classical Michaelian model. The enhanced VH+ in vps5Δ-reconstituted tonoplast vesicles resulted from an increase of the Vmax parameter relative to that of WT, whereas Km was found at about 1 M for both strains.
Figure 6.

Mg2+/H+ exchange rate of reconstituted tonoplast vesicles from vps5Δ (○) or WT (●) as a function of Mg2+ concentration. (A) Exchange rates (VH+) were determined from experimental kinetics illustrated in Fig. 4 and described in Materials and Methods. Values are presented as mean VH+ ± SE (n = 5). (B) Scatchard plot: Km values derived from linear regression of data in A are 0.96 ± 0.02 mM and 0.92 ± 0.01 mM for vps5Δ and WT strains, respectively; Vmax are 4.53 ± 0.36 and 2.93 ± 0.26%⋅min−1 per mg of protein.
Finally, Ni2+ growth phenotypes of 14 other yeast mutants of the vacuolar protein sorting pathway were determined (Fig. 7). Eight of these mutants also were shown to be defective in the retention of TGN membrane proteins: two other retromer complex mutants vps17Δ and vps29Δ (3), vps54Δ (24), vps1Δ (25), vps8Δ (26), tlg2Δ (27), vps13/soi1Δ (15), and grd19Δ (28). All these mutants displayed a NiR phenotype. In contrast, six other mutants, for which no alteration of this TGN retention process was described, showed a NiS phenotype: vps39Δ (29), vps41Δ (30), vam3Δ (31), vam7Δ (32), vps18Δ (33), and apl6Δ (34).
Figure 7.

Ni2+ growth phenotypes of yeast mutants of the vacuolar protein-sorting pathway. Mutants were grown on agar/YNB minimal medium containing 0.1 mM Mg2+, with (+) or without (−) 0.2 mM Ni2+. Phenotypes were recorded after 4 days. Class (34) and name of each mutant are indicated. ? indicates class not determined or not available in the literature. Among these mutants, only Ni2+-resistant mutants were found to be defective in the retention of Golgi membrane proteins. We refer the reader to the Yeast Proteome Database (http://www.proteome.com) for further information about the mutants and the corresponding genes (including alternative names and functional details).
Discussion
The vps5Δ Mutant Strain Displays an Enhanced Vacuolar Mg2+ (Ni2+, Co2+)/H+Exchange Activity.
The vps5Δ strain belongs to class B vps mutants (35) and displays fragmented vacuoles (refs. 6 and 7; data not shown). However, no gross alteration of functional properties of the vps5Δ vacuole could be observed at the cellular level (35, 36). The present study confirms this conclusion at the biochemical level: the isolated vacuole membranes of WT and vps5Δ exhibit the same SDS/PAGE protein profiles, α-mannosidase and V-ATPase activities, and H+ exchange activities with K+, Ca2+, and Cd2+.
In contrast, we found that the vacuole membrane isolated from vps5Δ displays a higher H+ exchange activity with Mg2+, Ni2+, and Co2+. Ni2+ and Co2+ are two Mg2+ analogs commonly used to trace Mg2+ transport (37–42). They are not required for normal yeast growth and are usually omitted from standard growth media. Moreover, significant H+ exchange activities with these cytotoxic heavy metals were observed only at high concentrations (0.1 to 1 mM, data not shown), indicating that vacuolar Co2+/H+ and Ni2+/H+ exchanges are unlikely to play a physiological role in yeast in standard growth conditions. Conversely, the affinity of the H+ exchange with Mg2+ seems physiologically sound because it displayed a Km for Mg2+ at about 1 mM, close to the reported activity of Mg2+ in the yeast cytosol (43). Moreover, the yeast vacuole has been described as an essential Mg2+ reservoir (44). The present work provides direct evidence for the activity of a Mg2+/H+ transport system at the tonoplast of S. cerevisiae. The plant vacuolar protein AtMHX was proposed to be an Mg2+/H+ exchanger (45). Nevertheless, no clear homologue could be detected in yeast.
C2+/H+ exchangers have been shown to promote a net vacuolar uptake of C2+, driven by the exergonic efflux of H+ out of acidic vacuoles in yeast cells (46–48). Thus, the rate of vacuolar uptake by C2+/H+ exchangers is expected to depend on the size of the ΔpH. In the present study, WT and vps5Δ exhibited similar vacuolar H+-ATPase activities. Furthermore, their growth capacity at pH 7.5, known to depend on the presence of a functional vacuolar H+-ATPase (49, 50), was similar on standard media. In agreement with published data (6, 35), the vacuole should be energized by similar ΔpH in both strains.
In conclusion, the vacuole membrane of the vps5Δ strain was found to exhibit similar biochemical and functional features compared with those of the WT strain, except for an enhanced Mg2+ (Ni2+, Co2+)/H+ exchange activity. The latter is expected to mediate a higher vacuolar uptake of C2+ in mutant yeast cells. Consistently, Ni2+ uptake was 3-fold higher in vps5Δ cells than in WT cells after plasma-membrane permeabilization. By comparison, Ni2+ uptakes by nonpermeabilized mutant cells and WT cells measured in short-term experiments were the same, indicating that both strains would display similar transport properties at the plasma-membrane level.
The vps5Δ Mutant Displays New Growth Phenotypes Likely Related to its Enhanced Vacuolar Mg2+ (Ni2+, Co2+)/H+Activity.
In the present study, a new selectable growth phenotype is presented for mutants altered in the recycling of Golgi membrane proteins: all mutants tested were resistant to Ni2+. In addition, the vps5Δ strain was resistant to Co2+, but sensitive to Cd2+, Al3+, and Mg2+-limiting media.
The vacuole has been described as a major site for Cd2+ detoxification (51, 52). As mentioned earlier, vps5Δ displays a fragmented vacuome composed of a large number of minivacuoles. Unexpectedly, the amounts of tonoplast isolated per g fresh weight of vps5Δ and WT were the same, suggesting that vacuomes of both strains might share the same tonoplast surface area despite their different morphologies. With the same areas but smaller vacuoles, a reduction of the vacuome volume might be expected in mutant cells, which would supposedly be detrimental for metal tolerance. Consistently, the CdS phenotype of vps5Δ was shared by vps41Δ (data not shown) and vps54Δ (53), two class B mutants showing fragmented vacuoles (30, 35, 53). In addition, vps18Δ, a class C mutant lacking any structure resembling a normal vacuole (35), also displays a CdS phenotype (54). Thus, a reduction of the vacuome volume might be responsible for the CdS phenotype of vps5Δ.
Along the same lines, the sensitivity of vps41Δ and vps18Δ to Ni2+ could be related as well to such a reduction of the vacuome volume. Indeed, the vacuole has been described also as a major site for Ni2+ detoxification (48, 55, 56). Because class B mutants, showing fragmented vacuoles, displayed either NiS (vps41Δ, vps39Δ, vam3Δ, vam7Δ) or NiR (vps5Δ, vps17Δ, vps54Δ) phenotypes (Fig. 7), the assumed reduction in size of their vacuome cannot account for the phenotype of NiR class B mutants. Therefore, we propose that the NiR (and CoR) phenotypes of vps5Δ would result from the marked enhancement of its vacuolar Ni2+ (Co2+)/H+ exchange activity. This transport activity would improve Ni2+ (Co2+) detoxification by increasing their vacuolar compartmentalization in vps5Δ (as shown for Ni2+, Fig. 2).
In contrast, vacuolar sequestration of Mg2+ by Mg2+/H+ exchange should be responsible for both WT and vps5Δ growth impairments observed at acidic pH in Mg2+-limiting media (Fig. 1A). Importantly, acidic conditions alone were not responsible for this effect because the latter was not observed on media containing nonlimiting Mg2+ concentrations. On the other hand, the lower the pH in the medium, the lower the pH in the vacuole (57), and thus, the higher the ΔpH energizing vacuolar ion uptake by cation/proton exchangers. In the present study, an increase of the vacuolar Mg2+ uptake is expected at acidic pH. In this condition, the increase of the vacuolar uptake would exceed the low cellular Mg2+ uptake occurring in low-Mg2+ media, thereby impairing cytosolic Mg2+ homeostasis. This result would explain the decrease of yeast growth at acidic pH in low-Mg2+ (but not in nonlimiting Mg2+) media, depicted horizontally in Fig. 1A. At a given pH, i.e., at a constant vacuolar ΔpH, cellular Mg2+ uptake would decrease below vacuolar uptake upon Mg2+ deprivation. This imbalance would also impair Mg2+ cytosolic homeostasis and would explain the yeast growth decrease depicted vertically in Fig. 1A.
In this context, the enhanced vacuolar Mg2+/H+ activity of vps5Δ, relative to that of WT, should be responsible for the higher sensitivity of the mutant to the conditions described above. The AlS phenotype of vps5Δ could be interpreted along the same line, because Al3+ toxicity was shown to result from the Al3+-inhibition of Mg2+ cellular uptake in yeast (42, 58), thereby mimicking Mg2+-limiting conditions. Finally, it should be noted that the low-MgS growth phenotype of vps5Δ is clearly observable at acidic pH relative to neutral pH, which is consistent with the implication of the vacuole energization in this phenotype.
Missorting of a TGN Mg2+/H+ Exchanger to the Vacuole Membrane of vps5Δ: A Molecular Hypothesis.
One straightforward interpretation of the increase of vacuolar C2+/H+ exchange activity in vps5Δ would be that the endogenous vacuolar exchanger is more active in mutant cells. However, deregulation of this exchanger (resulting, for example, in an increase of the exchanger density at the membrane surface or of the proportion of exchangers in the active state) is expected to increase the Vmax of the transport reaction without modification of its selectivity. Compared with WT, vps5Δ exhibited an increase of the Vmax, but also a marked alteration of ion selectivity: the selectivity sequence was Mg2+ = Co2+ > Ni2+ for WT and Co2+ > Mg2+ = Ni2+ for vps5Δ (Fig. 5). In addition, yeast vps5 mutants, like vps17 and vps29 mutants of the retromer complex and other mutants also altered in the recycling of Golgi membrane proteins (vps54Δ, vps1Δ, vps8Δ, tlg2Δ, vps13Δ, and grd19Δ), have been reported to missort various membrane proteins from the TGN (proteinase Kex2p, Vps10p, or dipeptidyl aminopeptidase A) to the tonoplast (3, 15, 24–28). In our study, all these mutants displayed a NiR phenotype, whereas other vacuolar protein sorting mutants not affected in this recycling process displayed a NiS phenotype. In this context, it may be hypothesized that vps5Δ, and possibly other mutants in retention of Golgi membrane proteins, would also missort a TGN Mg2+/H+ exchanger to the vacuole membrane of mutant cells. It is noteworthy that the literature data indicate that TGN and endosomal compartments sustain high ΔpH generated by the V-ATPase (59, 60).
In conclusion, our data argue in favor of the coexistence of two kinds of exchangers with distinct selectivities at the vacuolar membrane of vps5Δ (and possibly of other Golgi retention-defective mutants). Validation of this working hypothesis will require the molecular identification of yeast Mg2+/H+ exchangers. In wild-type cells, the TGN Mg2+/H+ exchanger would be recycled between the PVC and the TGN by the retromer complex and could be involved in the pH regulation of the TGN or Golgi-derived vesicles. Such a regulation is thought to be important for the sorting of secretory cargo and the retrieval of components of the biosynthetic pathway (59, 61). Recently, it has been shown that the VPS44 gene actually encodes the Nhx1p Na+/H+ exchanger of the PVC and that it is required for endosomal protein trafficking (62). It is noteworthy that neither Na+/H+ nor Ca2+/H+ exchanges were found to be involved in the regulation of the acidification of TGN vesicles (59).
Acknowledgments
We thank C. Grignon, P. Doumas, and T. Tranbarger for critical reading of the manuscript. We thank J.-P. Grouzis and J. Rigaud for their expertise in vacuole isolation and biochemical analyses, and M. Enjuto for initial help on yeast culture handling. G.B. was supported by a Fellowship from the Ministère de l'Education Nationale, de la Recherche, et de la Technologie.
Abbreviations
- TGN
trans-Golgi network
- CPY
carboxypeptidase Y
- PVC
prevacuolar compartment
- C2+
divalent cation
- WT
wild type
- ACMA
9-amino-6-chloro-2-methoxyacridine
- R and S
resistant and sensitive
Footnotes
This paper was submitted directly (Track II) to the PNAS office.
References
- 1.Bryant N J, Stevens T H. Microbiol Mol Biol Rev. 1998;62:230–247. doi: 10.1128/mmbr.62.1.230-247.1998. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 2.Lemmon S K, Traub L M. Curr Opin Cell Biol. 2000;12:457–466. doi: 10.1016/s0955-0674(00)00117-4. [DOI] [PubMed] [Google Scholar]
- 3.Seaman M N, McCaffery J M, Emr S D. J Cell Biol. 1998;142:665–681. doi: 10.1083/jcb.142.3.665. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 4.Nothwehr S F, Bruinsma P, Strawn L A. Mol Biol Cell. 1999;10:875–890. doi: 10.1091/mbc.10.4.875. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 5.Nothwehr S F, Ha S A, Bruinsma P. J Cell Biol. 2000;151:297–310. doi: 10.1083/jcb.151.2.297. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 6.Nothwehr S F, Hindes A E. J Cell Sci. 1997;110:1063–1072. doi: 10.1242/jcs.110.9.1063. [DOI] [PubMed] [Google Scholar]
- 7.Horazdovsky B F, Davies B A, Seaman M N, McLaughlin S A, Yoon S, Emr S D. Mol Biol Cell. 1997;8:1529–1541. doi: 10.1091/mbc.8.8.1529. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 8.Brachmann C B, Davies A, Cost G J, Caputo E, Li J, Hieter P, Boeke J D. Yeast. 1998;14:115–132. doi: 10.1002/(SICI)1097-0061(19980130)14:2<115::AID-YEA204>3.0.CO;2-2. [DOI] [PubMed] [Google Scholar]
- 9.Sherman F. Methods Enzymol. 1991;194:3–21. doi: 10.1016/0076-6879(91)94004-v. [DOI] [PubMed] [Google Scholar]
- 10.Roberts C J, Raymond C K, Yamashiro C T, Stevens T H. Methods Enzymol. 1991;194:644–661. doi: 10.1016/0076-6879(91)94047-g. [DOI] [PubMed] [Google Scholar]
- 11.Zinser E, Daum G. Yeast. 1995;11:493–536. doi: 10.1002/yea.320110602. [DOI] [PubMed] [Google Scholar]
- 12.Lichko L P, Okorokov L A. FEBS Lett. 1984;174:233–237. doi: 10.1016/0014-5793(84)81164-3. [DOI] [PubMed] [Google Scholar]
- 13.Laemmli U K. Nature (London) 1970;227:680–685. doi: 10.1038/227680a0. [DOI] [PubMed] [Google Scholar]
- 14.Oakley R B, Kirsch R D, Morris R N. Anal Biochem. 1980;105:361–363. doi: 10.1016/0003-2697(80)90470-4. [DOI] [PubMed] [Google Scholar]
- 15.Brickner J H, Fuller R S. J Cell Biol. 1997;139:23–36. doi: 10.1083/jcb.139.1.23. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 16.Landolt-Marticorena C, Williams K M, Correa J, Chen W, Manolson M F. J Biol Chem. 2000;275:15449–15457. doi: 10.1074/jbc.M000207200. [DOI] [PubMed] [Google Scholar]
- 17.Schaffner W, Weissman C. Anal Biochem. 1973;56:502–514. doi: 10.1016/0003-2697(73)90217-0. [DOI] [PubMed] [Google Scholar]
- 18.Amalou Z, Gibrat R, Trouslot P, d'Auzac J. Plant Physiol. 1994;106:79–85. doi: 10.1104/pp.106.1.79. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 19.Amalou Z, Enjuto M, Grouzis J P, Prevot J C, Jacob J L, Grignon C, Gibrat R. Plant Physiol Biochem (Paris) 1997;35:355–361. [Google Scholar]
- 20.Schumaker K S, Sze H. Plant Physiol. 1990;92:340–345. doi: 10.1104/pp.92.2.340. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 21.Ohsumi Y, Kitamoto K, Anraku Y. J Bacteriol. 1988;170:2676–2682. doi: 10.1128/jb.170.6.2676-2682.1988. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 22.Nass R, Cunningham K W, Rao R. J Biol Chem. 1997;272:26145–26152. doi: 10.1074/jbc.272.42.26145. [DOI] [PubMed] [Google Scholar]
- 23.Pouliquin P, Grouzis J-P, Gibrat R. Biophys J. 1999;76:360–373. doi: 10.1016/s0006-3495(99)77203-6. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 24.Conibear E, Stevens T H. Mol Biol Cell. 2000;11:305–323. doi: 10.1091/mbc.11.1.305. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 25.Wilsbach K, Payne G S. EMBO J. 1993;12:3049–3059. doi: 10.1002/j.1460-2075.1993.tb05974.x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 26.Chen Y J, Stevens T H. Eur J Cell Biol. 1996;70:289–297. [PubMed] [Google Scholar]
- 27.Holthuis J C, Nichols B J, Dhruvakumar S, Pelham H R. EMBO J. 1998;17:113–126. doi: 10.1093/emboj/17.1.113. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 28.Voos W, Stevens T H. J Cell Biol. 1998;140:577–590. doi: 10.1083/jcb.140.3.577. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 29.Nakamura N, Hirata A, Ohsumi Y, Wada Y. J Biol Chem. 1997;272:11344–11349. doi: 10.1074/jbc.272.17.11344. [DOI] [PubMed] [Google Scholar]
- 30.Radisky D C, Snyder W B, Emr S D, Kaplan J. Proc Natl Acad Sci USA. 1997;94:5662–5666. doi: 10.1073/pnas.94.11.5662. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 31.Darsow T, Rieder S E, Emr S D. J Cell Biol. 1997;138:517–529. doi: 10.1083/jcb.138.3.517. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 32.Ungermann C, Wickner W. EMBO J. 1998;17:3269–3276. doi: 10.1093/emboj/17.12.3269. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 33.Rieder S E, Emr S D. Mol Biol Cell. 1997;8:2307–2327. doi: 10.1091/mbc.8.11.2307. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 34.Cowles C R, Odorizzi G, Payne G S, Emr S D. Cell. 1997;91:109–118. doi: 10.1016/s0092-8674(01)80013-1. [DOI] [PubMed] [Google Scholar]
- 35.Raymond C K, Howald-Stevenson I, Vater C A, Stevens T H. Mol Biol Cell. 1992;3:1389–1402. doi: 10.1091/mbc.3.12.1389. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 36.Banta L M, Robinson J S, Klionsky D J, Emr S D. J Cell Biol. 1988;107:1369–1383. doi: 10.1083/jcb.107.4.1369. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 37.Fuhrmann G F, Rothstein A. Biochim Biophys Acta. 1968;163:325–330. doi: 10.1016/0005-2736(68)90117-x. [DOI] [PubMed] [Google Scholar]
- 38.Hmiel S P, Snavely M D, Miller C G, Maguire M E. J Bacteriol. 1986;168:1444–1450. doi: 10.1128/jb.168.3.1444-1450.1986. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 39.Joho M, Tarumi K, Inouhe M, Tohoyama H, Murayama T. Microbios. 1991;67:177–186. [PubMed] [Google Scholar]
- 40.Snavely M D, Gravina S A, Cheung T T, Miller C G, Maguire M E. J Biol Chem. 1991;266:824–829. [PubMed] [Google Scholar]
- 41.Nies D H. Plasmid. 1992;27:17–28. doi: 10.1016/0147-619x(92)90003-s. [DOI] [PubMed] [Google Scholar]
- 42.MacDiarmid C W, Gardner R C. J Biol Chem. 1998;273:1727–1732. doi: 10.1074/jbc.273.3.1727. [DOI] [PubMed] [Google Scholar]
- 43.Zhang A, Cheng T P O, Wu X Y, Altura B T, Altura B M. Cell Mol Life Sci. 1997;53:69–72. doi: 10.1007/PL00000581. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 44.Beeler T, Bruce K, Dunn T. Biochem Biophys Acta. 1997;1323:310–318. doi: 10.1016/s0005-2736(96)00199-x. [DOI] [PubMed] [Google Scholar]
- 45.Shaul O, Hilgemann D W, de-Almeida-Engler J, Van Montagu M, Inze D, Galili G. EMBO J. 1999;18:3973–3980. doi: 10.1093/emboj/18.14.3973. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 46.Okorokov L A, Kulakovskaya T V, Lichko L P, Polorotova E V. FEBS Lett. 1985;192:303–306. doi: 10.1016/0014-5793(85)80130-7. [DOI] [PubMed] [Google Scholar]
- 47.Pozos T C, Sekler I, Cyert M S. Mol Cell Biol. 1996;16:3730–3741. doi: 10.1128/mcb.16.7.3730. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 48.Nishimura K, Igarashi K, Kakinuma Y. J Bacteriol. 1998;180:1962–1964. doi: 10.1128/jb.180.7.1962-1964.1998. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 49.Nelson H, Nelson N. Proc Natl Acad Sci USA. 1990;87:3503–3507. doi: 10.1073/pnas.87.9.3503. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 50.Nelson N, Harvey W R. Physiol Rev. 1999;79:361–385. doi: 10.1152/physrev.1999.79.2.361. [DOI] [PubMed] [Google Scholar]
- 51.Ortiz D F, Kreppel L, Speiser D M, Scheel G, McDonald G, Ow D W. EMBO J. 1992;11:3491–3499. doi: 10.1002/j.1460-2075.1992.tb05431.x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 52.Li Z S, Lu Y P, Zhen R G, Szczypka M, Thiele D J, Rea P A. Proc Natl Acad Sci USA. 1997;94:42–47. doi: 10.1073/pnas.94.1.42. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 53.Conboy M J, Cyert M S. Mol Biol Cell. 2000;11:2429–2443. doi: 10.1091/mbc.11.7.2429. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 54.Clemens S, Kim E J, Neumann D, Schroeder J I. EMBO J. 1999;18:3325–3333. doi: 10.1093/emboj/18.12.3325. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 55.Joho M, Ishikawa Y, Kunikane M, Inouhe M, Tohoyama H, Murayama T. Microbios. 1992;71:149–159. [PubMed] [Google Scholar]
- 56.Ramsay L M, Gadd G M. FEMS Microbiol Lett. 1997;152:293–298. doi: 10.1111/j.1574-6968.1997.tb10442.x. [DOI] [PubMed] [Google Scholar]
- 57.Carmelo V, Santos H, Sa-Correia I. Biochim Biophys Acta. 1997;1325:63–70. doi: 10.1016/s0005-2736(96)00245-3. [DOI] [PubMed] [Google Scholar]
- 58.MacDiarmid C W, Gardner R C. Plant Physiol. 1996;112:1101–1109. doi: 10.1104/pp.112.3.1101. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 59.Demaurex N, Furuya W, D'Souza S, Bonifacino J S, Grinstein S. J Biol Chem. 1998;273:2044–2051. doi: 10.1074/jbc.273.4.2044. [DOI] [PubMed] [Google Scholar]
- 60.Nelson N, Perzov N, Cohen A, Hagai K, Padler V, Nelson H. J Exp Biol. 2000;203:89–95. doi: 10.1242/jeb.203.1.89. [DOI] [PubMed] [Google Scholar]
- 61.Mellman I. J Exp Biol. 1992;172:39–45. doi: 10.1242/jeb.172.1.39. [DOI] [PubMed] [Google Scholar]
- 62.Bowers K, Levi B P, Patel F I, Stevens T H. Mol Biol Cell. 2000;11:4277–4294. doi: 10.1091/mbc.11.12.4277. [DOI] [PMC free article] [PubMed] [Google Scholar]