

Abstract
Purpose of review
This study aims to discuss possible reasons why research to date has not forged direct links between light at night, acute melatonin suppression or circadian disruption, and risks for disease.
Recent Findings
Data suggest that irregular light–dark patterns or light exposures at the wrong circadian time can lead to circadian disruption and disease risks. However, there remains an urgent need to: (1) specify light stimulus in terms of circadian rather than visual response; (2) when translating research from animals to humans, consider species-specific spectral and absolute sensitivities to light; (3) relate the characteristics of photometric measurement of light at night to the operational characteristics of the circadian system; and (4) examine how humans may be experiencing too little daytime light, not just too much light at night.
Summary
To understand the health effects of light-induced circadian disruption, we need to measure and control light stimulus during the day and at night.
Keywords: Circadian rhythms, circadian phototransduction, circadian disruption, cancer in shift workers, light at night, daytime light
Introduction
Circadian rhythms in humans
Earth’s axial rotation results in exposure of all the planet’s creatures to 24-hour cycles of light and dark. Living organisms have adapted to this daily rotation by developing biological rhythms that have evolved as self-sustaining oscillations with their own unique sets of species-specific characteristics, including amplitude, phase, and period. Included among these self-sustaining oscillations are circadian rhythms, which repeat themselves at a period of approximately 24 hours.(1)
Circadian rhythms are generated endogenously, and are aligned with the environment by exogenous factors. It is widely known that, in mammals, circadian rhythms are regulated by an internal biological clock located in the suprachiasmatic nuclei (SCN) contained within the hypothalamus region of the brain.(2) The SCN are self-sustaining oscillators that maintain their daily activities for weeks when isolated and cultured. The SCN in humans have a free-running period that is slightly greater than 24 hours. Environmental cues can reset and synchronize the SCN daily, thereby ensuring that the organism’s behavioral and physiological rhythms are in synchrony with the daily rhythms in its environment. Information about the light–dark cycle, which is the main synchronizer of the SCN to the solar day, is received by the eyes’ photoreceptors and reaches the SCN via the retinohypothalamic tract (RHT). (3)
The three classes of photoreceptors in the retina (i.e., rods, cones, and intrinsically photosensitive retinal ganglion cells [ipRGCs]) participate in circadian phototransduction, which is the process used by the retina to convert light signals into electrical signals for the circadian system. In the 1980s, Ebihara and Tsuji showed that circadian entrainment was achieved in retinal-degenerated mice.(4) Soon thereafter, Foster et al. demonstrated that mice with severely degenerated classical photoreceptors exhibited normal circadian clock resetting by light.(5) These studies suggested that the retina contained a then-unidentified, novel class of photoreceptor. In 2002, Berson et al. identified the ipRGCs and demonstrated that they had a peak spectral sensitivity close to 484 nm, suggesting that melanopsin was the photopigment driving the ipRGCs’ photosensitivity.(6, 7)
Although ipRGCs are central to circadian phototransduction, the classical photoreceptors (i.e., rods and cones) also contribute to this response, as shown in studies using rodents.(8) Melanopsin-deficient mice, for example, can still be entrained to light–dark cycles and are still able to show a phase shifting response to bright white light, although these responses are attenuated by approximately 40 %.(9) Melanopsin-knockout mice, moreover, had a 45 % attenuation in phase shifting response to two higher irradiances of 480 nm light and an even greater attenuation at a lower irradiance.(10) Hattar et al. showed that genetically manipulated mice lacking melanopsin, coupled with a rod/cone system that was unable to signal light, failed to show any pupil light reflex, did not entrain to the light–dark cycle, and showed no masking effect to light.(8) Collectively, these studies suggest that while ipRGCs are needed to transmit signals emanating from the retina to the SCN, these signals contain information from all three classes of photoreceptors.
Several studies have shown that, at a cellular level, the circadian oscillations existing in most human cells result from a small group of clock genes inside the cell nuclei that create interlocked transcriptional and post-translational feedback loops.(1, 11-13) These cellular oscillators are synchronized to the master clock located in the SCN. Neural and hormonal stimuli transmit time information from the master clock to these peripheral clocks. In order to accomplish this, the SCN project to neuronal targets such as the endocrine neurons (i.e., the autonomic neurons of the paraventricular nucleus [PVN] of the hypothalamus and the sub-paraventricular zone of the hypothalamus). The SCN control the release of melatonin through the autonomic nervous system of the PVN. Melatonin is a hormone produced by the pineal gland at night and under conditions of darkness, thus its nickname of “the darkness hormone.”(14) Given its close interaction with the SCN, melatonin is believed to be a timing messenger, providing the entire body with information about time of day and seasons of the year (i.e., photoperiods). Light at night (LAN) will acutely suppress melatonin and can change the timing of melatonin synthesis. Since melatonin is believed to act as the main internal synchronizer of the peripheral clocks with the SCN, and the main synchronizer of information from the master clock between the peripheral clocks, suppression of the hormone melatonin or a change in the timing of melatonin secretion can lead to circadian disruption.
Maintaining the sequential and phase-relation ordering of the various circadian rhythms, from the molecular level to the behavioral level, is crucial for coordinated function throughout the human body. A person whose rhythms are synchronized to diurnal activity by the light–dark cycle and social activities must undergo phase readjustment when forced to adapt to a new light–dark cycle (or sleep–wake cycle in the case of night-shift workers). The SCN and peripheral clocks will eventually readapt to the new light–dark cycle, but several days are required to accomplish a complete phase shift. More importantly, the time required for each of the different peripheral clocks to re-entrain to the new light–dark cycle is variable, as some clock genes respond faster to the changes than others.(15) As a result, an internal desynchronization occurs between the SCN, which more quickly readjust to the new light–dark cycle, and the peripheral clocks. At the same time, a desynchronization also occurs between the various peripheral clocks. This internal desynchronization (or circadian disruption) affects the entire body, including sleep, alertness, digestion, and physical performance. This internal desynchronization is also believed to affect cell metabolism and proliferation, and has been shown to be associated with increased cancer risks. (16)
Circadian disruption and LAN
It has been postulated that electric LAN acts as an endocrine disruptor, either by acutely suppressing the hormone melatonin or by inducing circadian disruption, and therefore is linked to increased incidence of diseases in modern society. In fact, animal studies performed to date strongly suggest that both acute melatonin suppression by LAN and circadian disruption resulting from irregular light–dark patterns are associated with an increased rate of tumor growth and increased mortality.(17-20)
Shift work is used as a surrogate for overexposure to LAN resulting in melatonin suppression or circadian disruption. Indeed, a large number of epidemiological studies suggest an association between working rotating night shifts for 20–30 years and cancer risks, especially for breast and colorectal cancer. Notably, however, a few studies have failed to confirm this association (reviewed in Kolstad and Kamdar et al.(21, 22)). Given the limited human evidence, in combination with sufficient evidence gathered from experimental animals, in 2007 the International Agency for Research on Cancer (IARC) classified “shift work that involves circadian disruption” as a probable human carcinogen, Group 2A. (23) It should be noted that since the release of the IARC’s report in 2007, additional published studies have added support to an epidemiological link between shift work and cancer risks. (24-29)
Another mounting concern in contemporary society is that the use of self-luminous displays in the evening might be affecting melatonin levels, and consequently, sleep quantity and quality. Gradisar et al. showed that nine out of 10 survey participants reported using a technological device in the hour prior to bedtime.(30) While televisions were the most popular devices, the researchers also found that individuals under 30 years of age were more likely to use cellular phones than were those over 30 years of age. Not surprisingly, young adults, especially those who reported using interactive technological devices (e.g., cellular phones, tablets, video games), reported having significantly later sleep patterns, greater difficulty falling asleep, and less “refreshing” sleep. These results are consistent with recent studies showing that the use of self-luminous displays prior to bedtime can acutely suppress melatonin production, delay sleep onset, and decrease sleep duration, especially in young adults and adolescents.(31-34) As discussed below, however, one’s photic history (i.e., prior light exposures) also must be taken into account when determining whether light will have a detrimental effect on sleep. Rångtell et al. demonstrated, for example, that participants who used tablets during the evening and were exposed to >500 lux at the eye of white light for 6.5 hours during the daytime experienced effects that were not significantly different from the effects of reading a physical book.(35) Specifically, the participants’ evening use of tablets did not suppress melatonin, nor did it affect sleep quality and quantity.
Despite growing support of the notion that LAN is an endocrine disruptor, the research to date has not firmly established direct links between LAN, acute melatonin suppression or circadian disruption, and risks for disease (especially cancer in shift workers). The main goal of this review is to discuss several key points to address and explain why such links have not been established.
Key point 1: Light stimulus must be specified
There are five important characteristics of light for both the human visual and circadian systems: quantity, spectrum, timing, duration, and distribution. However, the ideal characteristics for vision are quite different from those that are most effective for the circadian system.(36, 37) Briefly, the quantity of polychromatic (white) light that is required to activate the circadian system is significantly greater than that which is required to activate the visual system (measured through acute melatonin suppression or phase shift of the dim light melatonin onset [DLMO]). The spectral sensitivity of the circadian system, again measured as acute melatonin suppression, peaks at short wavelengths (i.e., close to 460 nm), (38, 39) while the visual system is most sensitive to the middle-wavelength portion of the visible spectrum.(39) The photopic luminous efficiency function (V(λ)) is used as part of the fundamental definition of light to convert radiometric quantities to photometric quantities.(40) Although the V(λ) function, with a peak spectral response at 555 nm, is nearly universally used to characterize and measure all visually effective optical radiation, it is actually based upon the spectral sensitivity of the foveal cones (i.e., long- and middle-wavelength cones) in the central 2-degree visual field.
As described above, all classes of photoreceptors contribute to circadian phototransduction. All other spectral response functions, including those of the circadian system, are not formally recognized, so there is no official definition of light as it might impact the circadian system. In 2014, Lucas et al. proposed a toolbox that would allow researchers to report the effective irradiance experienced by each of the rod, cone, and ipRGCs photoreceptors involved in non-visual responses. (41) While this toolbox is helpful for equating the stimulus–response relationships employed in different studies, as well as for relating research findings to lighting conditions in the field, it provides no indication of the circadian system’s response to light stimulus. In other words, reporting how light stimulus excites ipRGC photoreceptors, though useful for researchers, does not provide any insight into how that stimulus will suppress production of the hormone melatonin.
In 2005, however, Rea et al. proposed a mathematical model of human circadian phototransduction (42, 43) based on fundamental knowledge of retinal neurophysiology and neuroanatomy and on published action spectrum data for acute melatonin suppression.(38, 39) Using this model, the density of light measured at the cornea is converted into circadian light (CLA), which is comparable to conventional photopic illuminance (i.e., the amount of luminous flux incident upon a given surface) but weighted by the spectral sensitivity of the human circadian system as measured by acute melatonin suppression after a 1-hour exposure. The CLA value can then be used to determine circadian stimulus (CS), which reflects the effectiveness of the spectrally weighted density of light incident at the eye from no suppression, or threshold (CS = 0.1), to saturation (CS = 0.7). Therefore, unlike the toolbox proposed by Lucas et al., the mathematical model proposed by Rea et al. allows researchers to predict the response of one outcome of the circadian system, namely, acute melatonin suppression, after 1-hour exposure to a specific light level and light spectrum. (41-43) Figure 1 shows the modeled spectral sensitivity of the human circadian system at a light level (300 scotopic lux at the cornea) needed to determine CLA at that light level, and Figure 2 shows the absolute sensitivity of the human circadian system plotted as a function of CLA. For reference, corresponding values for photopic illuminance, CLA, and CS for two common light sources (incandescent and daylight) are shown in Figure 2.
Figure 1.

The modeled spectral sensitivity of the human circadian system to narrowband light sources (dashed line) at a corneal illuminance of 300 scotopic lux; data points are the relative nocturnal melatonin suppression values published by Brainard et al. (open diamonds) and by Thapan et al. (open squares). [38, 39] Also shown are the modeled spectral sensitivity functions for two types of broad-band, polychromatic light sources, warm (solid gray line) and cool (solid black line). When a light source causes the blue versus yellow spectral opponent channel (b-y) to signal “yellow” (i.e., for a warm light source), the spectral sensitivity is defined in terms of the photopigment melanopsin. When the b-y channel signals “blue” (i.e., for a cool light source), the spectral sensitivity is defined in terms of melanopsin plus the short-wavelength sensitivity (S)-cone. Adapted from Chronobiology International.
Figure 2.

The modeled absolute sensitivity of the human circadian system based upon nocturnal melatonin suppression. Circadian stimulus (CS) (right ordinate) is directly proportional to measured nocturnal melatonin suppression (left ordinate), and both measures are plotted as a function of CLA, the corneal illuminance weighted by the spectral sensitivity functions in Fig. 1. Shown for reference are the ranges of indoor and outdoor light levels found in actual applications together with several daylight (dashed arrows) and incandescent (dotted arrows) corneal (photopic) illuminance levels. Note that for the same photopic illuminance, the daylight spectrum has a higher value of CS than the incandescent spectrum because daylight has relatively more shortwavelength energy to which the human circadian system is maximally sensitive. Adapted from Chronobiology International.
Light stimulus is not, however, only about absolute and spectral sensitivities. Timing also matters. Operation of the visual system does not depend significantly on the timing of light exposure; it responds well to a light stimulus at any time of the day or night. Depending on the timing of the exposure, light can phase advance or phase delay the biological clock.(44) Thus, the same light stimulus may be beneficial when given at one time of day and detrimental when given at another time of day.
In addition, while the visual system responds to a light stimulus very quickly (< 1 second), the duration of light exposure that is required to affect the circadian system can take much longer. To achieve measurable melatonin suppression from exposure to a moderate amount of light in young adults, for example, the required duration of light exposure is at least 5–10 minutes. (45,46) Recent studies have shown that the circadian system will respond to brief flashes of light, as long as those flashes are delivered continuously for at least 1 hour. (47, 48)
For the visual system, spatial light distribution is critical (e.g., when reading black letters on white paper), while the circadian system does not respond to spatial patterns. One study showed that light reaching the lower retina is more effective at suppressing melatonin than light reaching the upper retina, (49) while another claims that the nasal part of the retina is more sensitive to light for the circadian system than the peripheral retina. (50) Regardless, it is the total light exposure reaching the retina that is effective for eliciting a circadian response.
The short-term history of light exposure affects the sensitivity of the circadian system to light. For example, the higher the exposure to light during the day (e.g., 4 hours per day for 1 week of exposure to outdoor light), the lower the sensitivity of the circadian system to LAN (as measured by nocturnal melatonin suppression). (51) Recently, Figueiro showed that for a lighting intervention that is designed to either phase advance or phase delay the circadian system, phase shifting predictions can only be made if the total light exposures during waking hours—not just those from the lighting intervention—are taken into consideration. (52)
When specifying the stimulus, it is necessary to account for all of the lighting characteristics discussed above; in other words, the dose (i.e., quantity, spectrum, timing, duration) must be specified. Wood et al. (31) showed that a 1-hour exposure to iPads did not significantly suppress melatonin, but a 2-hour exposure did (i.e., an average 3 % suppression was observed after a 1-hour exposure and an average 23 % suppression was observed after a 2-hour exposure). Age also appears to matter in this regard. Recently, Figueiro and Overington showed that for the same circadian stimulus, teenagers (aged 15-17 years) suppressed more melatonin after exposure to self-luminous displays (on average 25 % after a 1-hour exposure) than did middle-aged adults and college students (on average 3 % after a 1-hour exposure). (53) These results are consistent with the findings of Crowley et al., who also showed that pre-pubertal children are more sensitive to evening light for melatonin suppression than post-pubertal adolescents. (54) This increase in sensitivity to light combined with increasing use of self-luminous displays in the evening may have long-term consequences, given that early-life environmental exposures may affect behavior and physiology in adulthood. (55)
In summary, when determining whether a lighting system is effective at activating the circadian system, the amount, spectrum, timing, duration, and total light exposure (or photic history) must be specified.
Key point 2: Species-specific spectral and absolute sensitivities to light must be considered
To date, many of the studies relating LAN and health risks have used nocturnal rodents. (56-58) One important point to keep in mind, however, is that the spectral and absolute sensitivities to light are species-specific. Animal work, therefore, can be only be translated to human models once these spectral and absolute sensitivities to light are taken into account.
In terms of absolute sensitivities, Figure 3, which is based on the review of circadian responses to light by Bullough et al., shows that there is a very large difference in the absolute sensitivities of the circadian systems of nocturnal rodents and humans. (59) The ratio for the thresholds for melatonin suppression and for circadian phase shifting in nocturnal rodents and in humans is between 3,000:1 and 10,000:1, both for polychromatic white light and narrow-band spectra light presented near the respective peaks of their spectral sensitivities. It should be noted that while diurnal rodents are similarly sensitive to light as humans, these species are rarely used as models for studies investigating the negative effects of LAN on health.
Figure 3.
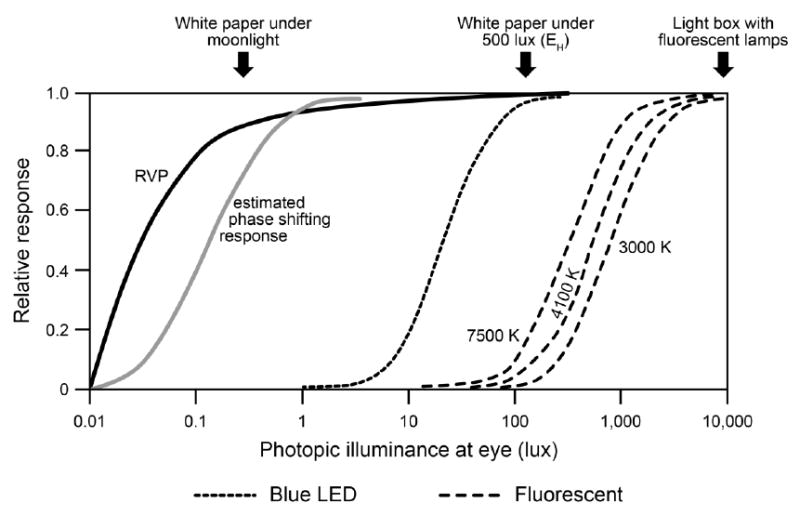
Relative visual performance (RVP, which is a measure of visual acuity) and estimated melatonin suppression after 1-hour exposure to 470-nm-peaking (blue) light emitting diode (LED) and fluorescent light sources (at three nominal correlated color temperatures, or CCTs) compared to the estimated phase shifting response in mice after a 30-minute light pulse as a function of photopic illuminance at the eye. The sensitivity of the nocturnal rodent circadian system is much greater than the human circadian system. It should be noted that a 7500 K CCT emits more short-wavelength content than a 3000 K light source, and is therefore, more effective at activating the circadian system. CCT is not, however, a good metric to compare effectiveness of various light sources for the circadian system and users should ask for the spectral power distribution of the light source rather than simply the CCT values. Photopic illuminance at the eye is shown because it is used by many researchers and end users.
The differences between the absolute sensitivities of rodents and humans may be even greater than indicated in Figure 3 because it is common to measure irradiance on the floors of cages where the animals are housed, while human studies report measured (vertical) irradiances at the subjects’ eyes. Vertical illuminances at human eye level tend to be about 1/3 to 1/5 the magnitude of the horizontal illuminances on work surfaces reported and recommended for buildings. (36, 60) Therefore, it is not unreasonable to suppose that there can be an absolute sensitivity ratio even greater than 10,000:1 between species, as it relates to their respective living environments.
In terms of spectral sensitivity, it is known from previous studies that the mouse retina contains rods, two cone types (i.e., a UV-cone peaking close to 365 nm, and an M-cone peaking close to 510 nm), (61) and ipRGCs (peaking close to 480 nm). (6) The overall peak spectral sensitivity for phase shifting of wheel running activity is close to 500 nm in mice, (59) while the spectral sensitivity of the human circadian system, as measured by acute melatonin suppression and phase shifting of DLMO, is close to 460 nm. (38, 39, 62)
Moreover, in addition to spectral and absolute sensitivities, the duration of light exposure that is needed to elicit a criterion circadian response (e.g., nocturnal melatonin suppression) differs markedly between nocturnal rodents and humans; the time course of melatonin suppression is much faster in rodents than in humans. (46, 63-65) It is imperative, therefore, that light stimulus from studies employing nocturnal animals be scaled to better represent the spectral and absolute sensitivities of the human circadian system before translating the findings of those studies from nocturnal animals to humans.
Key point 3: Light stimulus must be measured with calibrated instruments
Epidemiological studies have shown a statistical association between LAN, as characterized by satellite photometry or self-reports of room brightness, and breast cancer incidence. (66-69) It is important, however, that the characteristics of photometric measurement of LAN be related to the operational characteristics of the circadian system (described above). The photopic luminous efficiency function, V(λ), which most commercially available photometers employ, does not represent the spectral response of the human circadian system, which is maximally sensitive to short wavelengths (i.e., blue light). (39, 42, 70-72)
Underscoring the importance of using calibrated measurements of LAN, especially when attempting to use satellite photometry as a surrogate exposure to LAN, Figure 4 shows the spectral sensitivity of the human circadian system for narrow-band spectra based on data from Brainard et al. (38) and Thapan et al. (39) The peak spectral sensitivity of the human circadian system as measured by acute melatonin suppression is approximately 460 nm, with limited sensitivity at long wavelengths (>580 nm), where the human visual system (as measured by visual acuity) is still very sensitive. Given that some of the epidemiological studies used the photometric values derived from U.S. Air Force Defense Meteorological Satellite Program Operation Linescan System (DMSP-OLS) images to characterize LAN, and related those to breast cancer incidence (66-69), Figure 4 also shows the spectral response characteristics of the photomultiplier tube (PMT) used by the U.S. Air Force. This spectral response function is based upon what is termed the F16 response characteristic, (73) which extends from about 500 nm into the invisible, infrared region of the electromagnetic spectrum and is typical of those from earlier satellite PMTs (F12, F14, and F15). To more closely match sensitivity range of the PMT to the visible portion of the electromagnetic spectrum, an astronomical V-band filter (74) is commonly used with raw satellite PMT data. This filter provides photometric values more comparable to those based upon V(λ), the spectral weighting function used in commercial photometry. As illustrated in Figure 4, the V-band spectral weighting function matches the V(λ) response closely. As can also be seen in Figure 4, photometric measurements based upon: (1) the V(λ), (2) the V-band filter, and certainly (3) the raw PMT data will not accurately characterize the effectiveness of optical radiation incident on the human retina for circadian system stimulation, which peaks at short wavelengths.
Figure 4.

Spectral response characteristics relevant to the discussion of LAN. V(λ), shown with the dashed line, is the photopic luminous efficiency curve used in commercial photometry, closely mimicking the spectral sensitivity of the human fovea. The data points from Brainard et al. and Thapan et al. represent the spectral sensitivity of the human circadian system to narrowband spectra measured in terms of their effectiveness to each suppress the synthesis of melatonin at night. (38,39) The solid line represents the predictions from Rea et al. for the spectral sensitivity of the human circadian system to these narrow band spectra. (42) The spectral sensitivity of the satellite PMT currently used in the DMSP-OLS (F16) and those data filtered with an astronomical V-band filter (74) are shown as dot-dashed and dotted lines, respectively. The maximum value from all spectral response characteristics has been normalized to unity. Used with permission from Chronobiology International [75]
In terms of measuring exposure to LAN in shift workers and how it may affect melatonin levels, only a few studies to date have measured light exposure and melatonin levels in the field. As reviewed by Hunter and Figueiro, three studies conducted in real-world settings showed that light levels at work were below what is known to be required for the activation of the circadian system (< 80 lux). (76-80) Despite the low light levels, even after controlling for potential confounders, night-shift workers had lower melatonin (or its metabolite) concentrations than day-shift workers, and their peak levels tended to occur later than in day-shift workers. Moreover, the greater the number of consecutive nights worked, the greater the reduction in melatonin concentrations. These results suggest that LAN experienced by shift workers may not be acutely suppressing melatonin. Instead, the data suggest that misalignment of circadian rhythms may lead to the observed lower overall melatonin amplitude, in turn suggesting that circadian disruption may be the underlying cause for the observed health risks associated with shift work.
These poorly resolved, conflicting results point to an immediate need for commercializing light meters and developing measuring tools that are calibrated to measure circadian-effective light. Field data of circadian light exposures will further our knowledge and help scientists forge the direct link between LAN and health risks. Shift work doubtlessly results in circadian disruption and is associated with health risks, but the direct link between LAN and health risks will not be established until more scientific rigor is applied to the specification and measurement of the light stimulus.
Key point 4: Daytime light exposures must also be considered
Light and dark are the primary regulators of circadian rhythms, and light–dark exposures differ quantitatively between societies and eras. A century ago, prior to the widespread use of electric lighting, people in the United States were exposed to very bright days and very dark nights. This is probably true today only for those who live in agrarian societies or perform some form of outdoor work. With increasing frequency, however, people throughout the world are living in urban or suburban built environments and, in terms of circadian regulation, are likely to experience extended dim days and nights.
In fact, data obtained using an instrument designed to measure both conventional photopic illuminance and circadian light exposure at the eye throughout the waking day showed that people working inside the built environment experience much lower light levels compared to those living in an agrarian society. (81, 82) People working indoors are exposed to dim, extended and aperiodic circadian light, whereas those working outdoors are likely to be exposed to a robust light–dark cycle that is ideal for regulating the circadian system. These differences are probably even more pronounced during winter, when the duration of daylight becomes shorter. The disruption of the circadian cycle by dim, extended, aperiodic light exposure that is now probably experienced by most people in modern societies may very well have a direct influence on diseases associated with circadian disruption.
Boubekri et al. showed that people working in offices without access to windows reported poorer sleep quality, shorter sleep duration, and more-frequent sleep disturbances as assessed by self-reports and actigraphy recordings. (83) Recently, Figueiro et al. also showed that compared to receiving low levels of circadian-effective light in the morning, office workers receiving high levels of circadian-effective light in the morning experienced reduced sleep onset latency (especially in winter months), increased measures of circadian entrainment (phasor magnitude), and improved measures of sleep quality and mood. (84) While much of the discussion about light as an endocrine disruptor has centered on exposure to LAN, these recent studies underscore the importance of daytime light exposure for affecting circadian entrainment and nighttime sleep.
Conclusions
The importance of the connection between LAN, circadian disruption, and health risks will undoubtedly motivate researchers conducting future longitudinal studies of circadian disruption to devise better protocols for measuring and specifying LAN exposure. One key point which needs to be considered is that personal light exposures should be measured using calibrated devices that measure light as it affects the circadian system rather than the visual system (photopic light). Although in practical terms it is unlikely that those suffering from circadian disruption (e.g., shift workers) could wear calibrated light meters for long periods, studies nonetheless could be designed to sample light conditions and measure circadian entrainment at significant mileposts. This approach would permit more accurate extrapolation of circadian disruption’s effects on health outcomes. Another alternative would be for researchers to calibrate subjective scales using personal, calibrated sensors in a smaller group of subjects prior to administering questionnaires to a larger group of people. This approach would certainly help epidemiologists and would allow for larger data sets to be collected.
Clearly, a more sophisticated understanding of the influences of light and dark on circadian entrainment and disruption must be encouraged to emerge. There is now clear evidence that a strong light–dark cycle robustly drives circadian rhythms at the behavioral and at the cellular levels. (85) It is likely that people living in industrialized societies, however, do not experience a strong light–dark cycle and instead receive too little light during the day and too much light in the evening. The better we understand and control the stimulus, the more likely we are to understand the health effects of light-induced circadian disruption. We cannot move into what Turek calls “the new frontier for medicine: circadian medicine” without first being able to measure and specify circadian-effective light. (1)
Acknowledgments
This article was written with the support of a grant from the National Institute for Occupational Safety and Health, No. R01OH010668.
Footnotes
Conflict of Interest
Mariana G. Figueiro has nothing to disclose.
References
Papers of particular interest have been highlighted Important (•).
- 1.Turek FW. Circadian clocks: Not your grandfather’s clock. Science. 2016;354(6315):992–3. doi: 10.1126/science.aal2613. [DOI] [PubMed] [Google Scholar]
- 2.Stephan FK, Zucker I. Circadian rhythms in drinking behavior and locomotor activity of rats are eliminated by hypothalmaic lesions. Proc Natl Acad Sci USA. 1972;69:1583–6. doi: 10.1073/pnas.69.6.1583. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 3.Klein DC, Moore RY, Reppert SM. Suprachiasmatic nucleus: the mind’s clock. New York, NY: Oxford University Press; 1991. [Google Scholar]
- 4.Ebihara S, Tsuji K. Entrainment of the circadian activity rhythm to the light cycle: effective light intensity for a Zeitgeber in the retinal degenerate C3H mouse and the normal C57BL mouse. Physiol Behav. 1980;24(3):523–7. doi: 10.1016/0031-9384(80)90246-2. [DOI] [PubMed] [Google Scholar]
- 5.Foster R, Provencio I, Hudson D. Circadian photoreception in the retinally degenerate mouse (rd/rd) J Comp Physiol. 1991;169(1):39–50. doi: 10.1007/BF00198171. [DOI] [PubMed] [Google Scholar]
- 6.Berson D, Dunn F, Takao M. Phototransduction by retinal ganglion cells that set the circadian clock. Science. 2002;295(5557):1070–3. doi: 10.1126/science.1067262. [DOI] [PubMed] [Google Scholar]
- 7.Provencio I, Rodriguez IR, Jiang G, Hayes WP, Moreira EF, Rollag MD. A novel human opsin in the inner retina. J Neurosci. 2000;20(2):600–5. doi: 10.1523/JNEUROSCI.20-02-00600.2000. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 8.Hattar S, Lucas RJ, Mrosovsky N, Thompson SH, Douglas RH, Hankins MW, et al. Melanopsin and rod-cone photoreceptive systems account for all major accessory visual functions in mice. Nature. 2003;424:75–81. doi: 10.1038/nature01761. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 9.Ruby N, Brennan T, Xie X. Role of melanopsin in circadian responses to light. Science. 2002;298:2211–3. doi: 10.1126/science.1076701. [DOI] [PubMed] [Google Scholar]
- 10.Panda S, Provencio I, Tu DC, Pires SS, Rollag MD, Castrucci AM, et al. Melanopsin is required for non-image-forming photic responses in blind mice. Science. 2003;301(5632):525–7. doi: 10.1126/science.1086179. [DOI] [PubMed] [Google Scholar]
- 11.Panda S, Hogenesch JB. It’s all in the timing: many clocks, many outputs. J Biol Rhythms. 2004;19:374–87. doi: 10.1177/0748730404269008. [DOI] [PubMed] [Google Scholar]
- 12.Partch CL, Green CB, Takahashi JS. Molecular architecture of the mammalian circadian clock. Trends Cell Biol. 2014;24(2):90–9. doi: 10.1016/j.tcb.2013.07.002. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 13.Lim C, Allada R. Emerging roles for post-transcriptional regulation in circadian clocks. Nat Neurosci. 2013;16(11):1544–50. doi: 10.1038/nn.3543. [DOI] [PubMed] [Google Scholar]
- 14.Arendt J. Melatonin and the mammalian pineal gland. London, UK: Chapman & Hall; 1994. [Google Scholar]
- 15.Haus E, Smolensky M. Biological clocks and shift work: circadian dysregulation and potential long-term effects. Cancer Causes Control. 2006;17(4):489–500. doi: 10.1007/s10552-005-9015-4. [DOI] [PubMed] [Google Scholar]
- 16.Sahar S, Sassone-Corsi P. Metabolism and cancer: the circadian clock connection. Nat Rev Cancer. 2009;9(12):886–96. doi: 10.1038/nrc2747. [DOI] [PubMed] [Google Scholar]
- 17.Blask DE, Dauchy RT, Sauer LA. Putting cancer to sleep at night: the neuroendocrine/circadian melatonin signal. Endocrine. 2005;27(2):179–88. doi: 10.1385/ENDO:27:2:179. [DOI] [PubMed] [Google Scholar]
- 18.Filipski E, King VM, Li X, Granda TG, Mormont M-C, Claustrat B, et al. Disruption of circadian coordination accelerates malignant growth in mice. Pathol Biol (Paris) 2003;51(4):216–9. doi: 10.1016/s0369-8114(03)00034-8. [DOI] [PubMed] [Google Scholar]
- 19.Filipski E, Delaunay F, King VM, Wu MW, Claustrat B, Grechez-Cassiau A, et al. Effects of chronic jet lag on tumor progression in mice. Cancer Res. 2004;64(21):7879–85. doi: 10.1158/0008-5472.CAN-04-0674. [DOI] [PubMed] [Google Scholar]
- 20.Stevens RG, Blask DE, Brainard GC, Hansen J, Lockley SW, Provencio I, et al. Meeting report: the role of environmental lighting and circadian disruption in cancer and other diseases. Environ Health Perspect. 2007;115(9):1357–62. doi: 10.1289/ehp.10200. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 21.Kolstad HA. Nightshift work and risk of breast cancer and other cancers--a critical review of the epidemiologic evidence. Scand J Work Env Hea. 2008;34(1):5–22. doi: 10.5271/sjweh.1194. [DOI] [PubMed] [Google Scholar]
- 22.Kamdar BB, Tergas AI, Mateen FJ, Bhayani NH, Oh J. Night-shift work and risk of breast cancer: a systematic review and meta-analysis. Breast Cancer Res Treat. 2013;138(1):291–301. doi: 10.1007/s10549-013-2433-1. • This review of 15 studies concludes that only weak evidence supports the association between night-shift work and increased risk for breast cancer. [DOI] [PubMed] [Google Scholar]
- 23.World Health Organization (WHO) International Agency for Research on Cancer (IARC) IARC Monographs on the Evaluation of Carcinogenic Risks to Humans: Painting, Firefighting, and Shiftwork. Lyon, France: World Health Organization (WHO); 2010. [Google Scholar]
- 24.Åkerstedt T, Knutsson A, Narusyte J, Svedberg P, Kecklund G, Alexanderson K. Night work and breast cancer in women: a Swedish cohort study. BMJ open. 2015;5(4):e008127. doi: 10.1136/bmjopen-2015-008127. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 25.Bonde JP, Hansen J, Kolstad HA, Mikkelsen S, Olsen JH, Blask DE, et al. Work at night and breast cancer-report on evidence-based options for preventive actions. Scand J Work Environ Health. 2012;38(4):380–90. doi: 10.5271/sjweh.3282. [DOI] [PubMed] [Google Scholar]
- 26.Cordina-Duverger E, Koudou Y, Truong T, Arveux P, Kerbrat P, Menegaux F, et al. Night work and breast cancer risk defined by human epidermal growth factor receptor-2 (HER2) and hormone receptor status: A population-based case-control study in France. Chronobiol Int. 2016 Apr 14;:1–5. doi: 10.3109/07420528.2016.1167709. [DOI] [PubMed] [Google Scholar]
- 27.Lin Y, Nishiyama T, Kurosawa M, Tamakoshi A, Kubo T, Fujino Y, et al. Association between shift work and the risk of death from biliary tract cancer in Japanese men. BMC Cancer. 2015;15:757. doi: 10.1186/s12885-015-1722-y. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 28.Lin X, Chen W, Wei F, Ying M, Wei W, Xie X. Night-shift work increases morbidity of breast cancer and all-cause mortality: a meta-analysis of 16 prospective cohort studies. Sleep Med. 2015;16(11):1381–7. doi: 10.1016/j.sleep.2015.02.543. [DOI] [PubMed] [Google Scholar]
- 29.Papantoniou K, Castaño-Vinyals G, Espinosa A, Aragonés N, Pérez-Gómez B, Ardanaz E, et al. Breast cancer risk and night shift work in a case-control study in a Spanish population. Eur J Epidemiol. 2016;31(9):867–78. doi: 10.1007/s10654-015-0073-y. [DOI] [PubMed] [Google Scholar]
- 30.Gradisar M, Wolfson AR, Harvey AG, Hale L, Rosenberg R, Czeisler CA. The Sleep and Technology Use of Americans: Findings from the National Sleep Foundation’s 2011 Sleep in America Poll. J Clin Sleep Med. 2013;9(12):1291–9. doi: 10.5664/jcsm.3272. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 31.Wood B, Rea MS, Plitnick B, Figueiro MG. Light level and duration of exposure determine the impact of self-luminous tablets on melatonin suppression. Appl Ergon. 2013;44(2):237–40. doi: 10.1016/j.apergo.2012.07.008. [DOI] [PubMed] [Google Scholar]
- 32.Hysing M, Pallesen S, Stormark KM, Jakobsen R, Lundervold AJ, Sivertsen B. Sleep and use of electronic devices in adolescence: results from a large population-based study. BMJ open. 2015;5(1):e006748. doi: 10.1136/bmjopen-2014-006748. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 33.Gamble AL, D’Rozario AL, Bartlett DJ, Williams S, Bin YS, Grunstein RR, et al. Adolescent sleep patterns and night-time technology use: results of the Australian Broadcasting Corporation’s Big Sleep Survey. PLoS One. 2014;9(11):e111700. doi: 10.1371/journal.pone.0111700. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 34.Falbe J, Davison K, Franckle R, Ganter C, Gortmaker SL, Smith L, et al. Sleep duration, restfulness, and screens in the sleep environment. Pediatrics. 2015;135(2):e367–e75. doi: 10.1542/peds.2014-2306. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 35.Rångtell FH, Ekstrand E, Rapp L, Lagermalm A, Liethof L, Búcaro MO, et al. Two hours of evening reading on a self-luminous tablet vs. reading a physical book does not alter sleep after daytime bright light exposure. Sleep Med. 2016;23:111–8. doi: 10.1016/j.sleep.2016.06.016. [DOI] [PubMed] [Google Scholar]
- 36.Rea MS, editor. IESNA lighting handbook: reference and application. 9. New York, NY: Illuminating Engineering Society of North America; 2000. [Google Scholar]
- 37.Rea MS, Bullough JD, Figueiro MG. Phototransduction for human melatonin suppression. J Pineal Res. 2002;32(4):209–13. doi: 10.1034/j.1600-079x.2002.01881.x. [DOI] [PubMed] [Google Scholar]
- 38.Brainard GC, Hanifin JP, Greeson JM, Byrne B, Glickman G, Gerner E, et al. Action spectrum for melatonin regulation in humans: evidence for a novel circadian photoreceptor. J Neurosci. 2001;21(16):6405–12. doi: 10.1523/JNEUROSCI.21-16-06405.2001. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 39.Thapan K, Arendt J, Skene DJ. An action spectrum for melatonin suppression: evidence for a novel non-rod, non-cone photoreceptor system in humans. J Physiol. 2001;535(1):261–7. doi: 10.1111/j.1469-7793.2001.t01-1-00261.x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 40.Commission Internationale de l’Éclairage. Light as a true visual quantity: principles of measurement. Paris: Commission Internationale de l’Éclairage; 1978. [Google Scholar]
- 41.Lucas RJ, Peirson SN, Berson DM, Brown TM, Cooper HM, Czeisler CA, et al. Measuring and using light in the melanopsin age. Trends Neurosci. 2014;37(1):1–9. doi: 10.1016/j.tins.2013.10.004. • This study proposed a toolbox allowing researchers to report the effective irradiance experienced by each of the rod, cone, and ipRGCs photoreceptors involved in non-visual responses. While this toolbox is helpful for equating the stimulus–response relationships employed in different studies, it provides no indication of the circadian system’s response to light stimulus. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 42.Rea MS, Figueiro MG, Bullough JD, Bierman A. A model of phototransduction by the human circadian system. Brain Res Rev. 2005;50(2):213–28. doi: 10.1016/j.brainresrev.2005.07.002. [DOI] [PubMed] [Google Scholar]
- 43.Rea MS, Figueiro MG, Bierman A, Hamner R. Modelling the spectral sensitivity of the human circadian system. Light Res Technol. 2012;44(4):386–96. [Google Scholar]
- 44.Jewett ME, Rimmer DW, Duffy JF, Klerman EB, Kronauer RE, Czeisler CA. Human circadian pacemaker is sensitive to light throughout subjective day without evidence of transients. Am J Physiol. 1997;273(42):1800–9. doi: 10.1152/ajpregu.1997.273.5.r1800. [DOI] [PubMed] [Google Scholar]
- 45.Figueiro MG, Wood B, Plitnick B, Rea MS. The impact of light from computer monitors on melatonin levels in college students. Neuro Endocrinol Lett. 2011;32(2):158–63. [PubMed] [Google Scholar]
- 46.McIntyre IM, Norman TR, Burrows GD, Armstrong SM. Human melatonin suppression by light is intensity dependent. J Pineal Res. 1989;6(2):149–56. doi: 10.1111/j.1600-079x.1989.tb00412.x. [DOI] [PubMed] [Google Scholar]
- 47.Figueiro MG. Individually tailored light intervention through closed eyelids to promote circadian alignment and sleep health. Sleep Health. 2015;1(1):75–82. doi: 10.1016/j.sleh.2014.12.009. • This field study demonstrated the feasibility of using light through closed eyelids during sleep for promoting circadian alignment and sleep health. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 48.Zeitzer JM, Fisicaro RA, Ruby NF, Heller HC. Millisecond flashes of light phase delay the human circadian clock during sleep. J Biol Rhythms. 2014;29(5):370–6. doi: 10.1177/0748730414546532. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 49.Glickman G, Hanifin JP, Rollag MD, Wang J, Cooper H, Brainard GC. Inferior retinal light exposure is more effective than superior retinal exposure in suppressing melatonin in humans. J Biol Rhythms. 2003;18(1):71–9. doi: 10.1177/0748730402239678. [DOI] [PubMed] [Google Scholar]
- 50.Visser EK, Beersma DG, Daan S. Melatonin suppression by light in humans is maximal when the nasal part of the retina is illuminated. J Biol Rhythms. 1999;14(2):116–21. doi: 10.1177/074873099129000498. [DOI] [PubMed] [Google Scholar]
- 51.Hebert M, Martin SK, Lee C, Eastman CI. The effects of prior light history on the suppression of melatonin by light in humans. J Pineal Res. 2002;33(4):198–203. doi: 10.1034/j.1600-079x.2002.01885.x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 52.Figueiro MG. Delayed sleep phase disorder: clinical perspective with a focus on light therapy. Nat Sci Sleep. 2016;8:91. doi: 10.2147/NSS.S85849. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 53.Figueiro MG, Overington D. Self-luminous devices and melatonin suppression in adolescents. Light Res Technol. 2016;46(8):966–75. [Google Scholar]
- 54.Crowley SJ, Cain SW, Burns AC, Acebo C, Carskadon MA. Increased Sensitivity of the Circadian System to Light in Early/Mid-Puberty. J Clin Endocrinol Metab. 2015;100(11):4067–73. doi: 10.1210/jc.2015-2775. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 55.Cissé YM, Peng J, Nelson RJ. Dim light at night prior to adolescence increases adult anxiety-like behaviors. Chronobiol Int. 2016;33(10):1473–80. doi: 10.1080/07420528.2016.1221418. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 56.Aubrecht TG, Jenkins R, Nelson RJ. Dim light at night increases body mass of female mice. Chronobiol International. 2015;32(4):557–60. doi: 10.3109/07420528.2014.986682. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 57.Fonken LK, Nelson RJ. The effects of light at night on circadian clocks and metabolism. Endocr Rev. 2014;35(4):648–70. doi: 10.1210/er.2013-1051. [DOI] [PubMed] [Google Scholar]
- 58.Fonken LK, Finy MS, Walton JC, Weil ZM, Workman JL, Ross J, et al. Influence of light at night on murine anxiety- and depressive-like responses. Behav Brain Res. 2009;205(2):349–54. doi: 10.1016/j.bbr.2009.07.001. [DOI] [PubMed] [Google Scholar]
- 59.Bullough JD, Rea MS, Figueiro MG. Of mice and women: light as a circadian stimulus in breast cancer research. Cancer Causes Control. 2006;17(4):375–83. doi: 10.1007/s10552-005-0574-1. [DOI] [PubMed] [Google Scholar]
- 60.Rea MS. Keynote Address: Light Isn’t Just for Vision Anymore!. Fifth International Lighting Research Symposium on Light and Human Health; November 3–5, 2002; Orlando, FL. Electric Power Research Institute, Lighting Research Office; [Google Scholar]
- 61.Jacobs GH, Williams GA, Fenwick JA. Influence of cone pigment coexpression on spectral sensitivity and color vision in the mouse. Vision Res. 2004;44(14):1615–22. doi: 10.1016/j.visres.2004.01.016. [DOI] [PubMed] [Google Scholar]
- 62.Wright H, Lack L. Effect of light wavelength on suppression and phase delay of the melatonin rhythm. Chronobiol Int. 2001;18(5):801–8. doi: 10.1081/cbi-100107515. [DOI] [PubMed] [Google Scholar]
- 63.Reiter R. Action spectra, dose-response relationships, and temporal aspects of light’s effects on the pineal gland. Annal NY Acad Sci. 1985;453:215–30. doi: 10.1111/j.1749-6632.1985.tb11812.x. [DOI] [PubMed] [Google Scholar]
- 64.Lewy AJ, Wehr TA, Goodwin FK, Newsome DA. Light suppresses melatonin secretion in humans. Science. 1980;210(4475):1267–9. doi: 10.1126/science.7434030. [DOI] [PubMed] [Google Scholar]
- 65.McIntyre IM, Norman TR, Burrows GD, Armstrong SM. Quantal melatonin suppression by exposure to low intensity light in man. Life Sci. 1989;45(4):327–32. doi: 10.1016/0024-3205(89)90142-2. [DOI] [PubMed] [Google Scholar]
- 66.Kloog I, Haim A, Stevens RG, Barchana M, Portnov BA. Light at night co-distributes with incident breast but not lung cancer in the female population of Israel. Chronobiol Int. 2008;25:65–81. doi: 10.1080/07420520801921572. [DOI] [PubMed] [Google Scholar]
- 67.Kloog I, Stevens RG, Haim A, Portnov BA. Nighttime light level co-distributes with breast cancer incidence worldwide. Cancer Causes Control. 2010;21(12):2059–68. doi: 10.1007/s10552-010-9624-4. [DOI] [PubMed] [Google Scholar]
- 68.Kloog I, Portnov BA, Rennert HS, Haim A. Does the modern urbanized sleeping habitat pose a breast cancer risk? Chronobiol Int. 2011;28:76–80. doi: 10.3109/07420528.2010.531490. [DOI] [PubMed] [Google Scholar]
- 69.Keshet-Sitton A, Or-Chen K, Huber E, Haim A. Illuminating a risk for breast cancer: a preliminary ecological study on the association between streetlight and breast cancer. Integr Cancer Ther. 2016 doi: 10.1177/1534735416678983. [Epub ahead of print] [DOI] [PMC free article] [PubMed] [Google Scholar]
- 70.Brainard GC, Sliney D, Hanifin JP, Glickman G, Byrne B, Greeson JM, et al. Sensitivity of the human circadian system to short-wavelength (420-nm) light. J Biol Rhythms. 2008;23(5):379–86. doi: 10.1177/0748730408323089. [DOI] [PubMed] [Google Scholar]
- 71.Glickman G, Levin R, Brainard GC. Ocular input for human melatonin regulation: relevance to breast cancer. Neuro Endocrinol Lett. 2002;23(supplement 2):17–22. [PubMed] [Google Scholar]
- 72.Kozaki T, Koga S, Toda N, Noguchi H, Yasukouchi A. Effects of short wavelength control in polychromatic light sources on nocturnal melatonin secretion. Neurosci Lett. 2008;439(3):256–9. doi: 10.1016/j.neulet.2008.05.035. [DOI] [PubMed] [Google Scholar]
- 73.Ziskin D. OLS #16 acceptance test report, CDRL 066A2, digitized by Fabio Falchi. Personal communication, December 2010. F16 tabular data provided by Mr Ziskin. Cooperative Institute for Research in Environmental Sciences (CIRES), National Oceanic and Atmospheric Administration (NOAA), National Geophysical Data Center (NGDC), National Geophysical Data Center, Westinghouse Electric Corporation; 1990. [Google Scholar]
- 74.Mermilliod JC, Hauck B, Mermilliod M. The general catalogue of photometric data. [18 Jan 2017];Institut d’Astronomie de l’Universite de Lausanne, Versoix, Switzerland: observatoire de Genève. 2008 http://obswww.unige.ch/gcpd/filters/fil01.html.
- 75.Rea MS, Brons JA, Figueiro MG. Measurements of light at night (LAN) for a sample of female school teachers. Chronobiol Int. 2011;28(8):673–80. doi: 10.3109/07420528.2011.602198. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 76.Hunter CM, Figueiro MG. Measuring light at night and melatonin levels in shift workers: a review of the literature. Biol Res Nurs. 2016 doi: 10.1177/1099800417714069. In press. • This review summarizes research detailing field studies of cancer in human participants, specifically shift workers whose exposure to LAN was quantitatively assessed in some way and whose melatonin or its metabolites was quantitatively measured over a relevant period. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 77.Grundy A, Sanchez M, Richardson H, Tranmer J, Borugian M, Graham CH, et al. Light intensity exposure, sleep duration, physical activity, and biomarkers of melatonin among rotating shift nurses. Chronobiol Int. 2009;26(7):1443–61. doi: 10.3109/07420520903399987. [DOI] [PubMed] [Google Scholar]
- 78.Grundy A, Tranmer J, Richardson H, Graham CH, Aronson KJ. The influence of light at night exposure on melatonin levels among Canadian rotating shift nurses. Cancer Epidemiol Biomarkers Prev. 2011;20(11):2404–12. doi: 10.1158/1055-9965.EPI-11-0427. [DOI] [PubMed] [Google Scholar]
- 79.Dumont M, Lanctot V, Cadieux-Viau R, Paquet J. Melatonin production and light exposure of rotating night workers. Chronobiol Int. 2012;29(2):203–10. doi: 10.3109/07420528.2011.647177. [DOI] [PubMed] [Google Scholar]
- 80.Zeitzer JM, Dijk DJ, Kronauer R, Brown E, Czeisler C. Sensitivity of the human circadian pacemaker to nocturnal light: melatonin phase resetting and suppression. J Physiol. 2000;526(3):695–702. doi: 10.1111/j.1469-7793.2000.00695.x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 81.Bierman A, Klein TR, Rea MS. The Daysimeter: A device for measuring optical radiation as a stimulus for the human circadian system. Meas Sci Technol. 2005;16:2292–9. [Google Scholar]
- 82.Figueiro MG, Rea MS, Stevens RG, Rea AC. Daylight and productivity: A possible link to circadian regulation. Light and Human Health: EPRI/LRO 5th International Lighting Research Symposium; Palo Alto, CA: Electric Power Research Institute Lighting Research Office; 2002. pp. 185–93. [Google Scholar]
- 83.Boubekri M, Cheung I, Reid K, Wang C, Zee P. Impact of windows and daylight exposure on overall health and sleep quality of office workers: a case-control pilot study. J Clin Sleep Med. 2014;10(6):603–11. doi: 10.5664/jcsm.3780. • This study showed that people working in offices without access to windows reported poorer sleep quality, shorter sleep duration, and more-frequent sleep disturbances. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 84.Figueiro MG, Steverson B, Heerwagen J, Kampschroer K, Hunter CM, Gonzales K, et al. The impact of daytime light exposures on sleep and mood in office workers. Sleep Health. 2017;3(3):204–215. doi: 10.1016/j.sleh.2017.03.005. [DOI] [PubMed] [Google Scholar]
- 85.Reddy AB, Wong GK, O’Neill J, Maywood ES, Hastings MH. Circadian clocks: neural and peripheral pacemakers that impact upon the cell division cycle. Mutat Res. 2005;574(1-2):76–91. doi: 10.1016/j.mrfmmm.2005.01.024. [DOI] [PubMed] [Google Scholar]