

Abstract
The Achillea millefolium L. (Yarrow) is a common herb which is widely being used, worldwide. Achillea is being used for treatment of many disorders since centuries. It is considered safe for supplemental use and flavonoids such as kaempferol, luteolin and apigenin are of main constituents present in Achillea. Most of both antioxidant and anti-inflammatory properties of this herb have been attributed to its flavonoid content. Oxidative and inflammatory processes play important roles in pathogenesis of neurodegenerative diseases. Present review was aimed to review the latest literature evidences regarding application of Achillea and/or its three main flavonoid constituents on epilepsy, Alzheimer’s disease, multiple sclerosis, Parkinson’s disease and stroke.
Keywords: Achillea millefolium, Flavonoids, Medicinal, Neurodegenerative – disease, Plants
Introduction
Achillea millefolium L. (yarrow), as a medicinal plant which grows worldwide, has been used for treatment of diseases and injuries since ancient historical era. Most often is called yarrow. Achillea by people and belongs to the family of Asteraceae and contains more than 100 species (1). Achillea has erect stems, with 30–60 cm height, herbaceous, perennial herb, simple or branched from the base, fern-like feathery leaves and pink-white as well as dense flowers (Figure 1) (2).
Figure 1.

Achillea millefolium L.
For many years, Achillea has popularly been used for treatment of ulcers and gastrointestinal disorders. Regardingly, recent studies have reported some other medicinal properties for this herb, including antioxidant (3), anti-inflammatory (4), antibacterial (5) and anti-hepatotoxic (6) effects.
A wide variety of bioactive components, including amino acids, fatty acids, salicylic and succinic acids (7, 8), ascorbic acid, folic acid (9), caffeic acid (10) and flavonoids (9) are found in Achillea. Among them, flavonoids (kaempferol: 3,5,7-Trihydroxy-2-(4-hydroxyphenyl)-4H-chromen-4-one, luteolin: 3’,4’, 5,7-tetrahydroxyflavone and apigenin: 4’,5,7-trihydro-xyflavone) have attracted more attentions in recent years and most of antioxidant and anti-inflammatory properties of this herb attributed to them (Figure 2).
Figure 2.
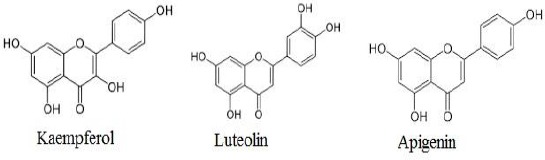
Chemical structure of main flavonoids of Achillea millefolium L.
Regarding the extensive body of literature on the critical role of oxidative stress in neuronal damage induced by neurodegenerative diseases (such as Parkinson’s disease and Alzheimer’s disease) and also considering the rich flavonoid content of Achillea, as aforementioned, in present article, we reviewed the studies which have used Achillea and/or its three main flavonoid constituents on nervous system related diseases (Table 1).
Table 1.
Summary of in vivo and in vitro studies of Achillea millefolium L. and its flavonoid contents on neurodegenerative diseases
| Herb and its flavonoid contents Disease | Achillea millefolium | Apigenin | Luteolin | Kaempferol |
|---|---|---|---|---|
| Multiple Sclerosis | Attenuate disease severity, inflammatory responses and demyelinating lesions in EAE (11) | Reduces the progression and relapse in EAE (12) | Suppresses behavioral deficits, prevented relapse and reduced inflammation in EAE (13), improve maturation of OPCs (14), delay the recovery of behavioral deficits in EAE (15) | |
| Alzheimer’s disease | Inhibits activated microglial cells and neuronal death in animal models of AD (16, 17), protect copper-mediated β-amyloid neurotoxicity in AD cell model (18), ameliorate AD-associated learning and memory deficits and relieve Aβ plaque formation (19) | Inhibits Aβ-induced neuronal death in mouse cortical cultures (20, 21), inhibits memory impairment, degeneration of cholinergic neurons and amyloid deposition (22, 23), the protective effect in traumatic brain injury induced AD (24) | Inhibits formation, extension, and destabilization of beta-amyloid fibrils in Escherichia coli (25), inhibits Aβ-induced toxicity in mouse-derived hippocampal neuronal cells (HT22) (26, 27), revers memory impairment in mice model of AD (28) | |
| Parkinson’s disease | Improves motility disorders and muscle tone in a rat (29) | Inhibited the disaggregation of alpha-synuclein oligomers (30), improves the locomotor and muscular activity and protected dopaminergic neurons (31) | Protects neural pheochromocytoma (PC12) and glial C6 cells (32), protect dopaminergic neurons from oxidant and inflammatory insults in PC12 cell line (33), improved the locomotor and muscular activities (31) | Prevents oxidative stress-induced cell death in rat PC12 and primary neuronal cells (34), improves motor coordination, raise striatal dopamine level and prevent the loss of dopaminergic neurons in mice (35) |
| Stroke | Reduction in infarct volume, neurological disorders and sensorimotor disorder in rat (36) | Inhibition of activated murine microglia cell line (37), neuroprotective effects on ROS-insulted primary neurons (38) | Lowered the risk of stroke (39), reduce serum levels of inflammatory markers (40), neuroprotective effect on ROS-insulted primary neurons (41), decreases infarct volume and suppresses edema in rat (42, 43), improve neurological status, cognitive abilities in human (44) | Lowered the risk of stroke (39), prevents ischemic brain injury and neuroinflammation in rats (45, 46) |
| Epilepsy | Reduces the seizure severity in rat (47) | Reduces the latency in the onset of picrotoxin-induced convulsions and inhibitory effects on GABA activated Cl(-) currents in cerebellar granule cells (48), antiepileptic effect and inhibition of Kainic acid-induced cytotoxicity in neurons (49) | Suppresses seizure intensity and duration (50) | Anti-epileptic effects (51), anticonvulsant effects (52) |
Experimental autoimmune encephalomyelitis (EAE), reactive oxygen species (ROS), oligodendrocyte precursor cells (OPCs), Alzheimer’s disease (AD)
Multiple Sclerosis
Multiple Sclerosis (MS) is a neuroinflammatory disorder with autoimmune base, characterized by a relapsing-remitting demyelinating phenomenon (53, 54).
Inflammatory and oxidant pathways (via modulation of immune system) are thought to be involved in pathogenesis of the disease (55). The experimental autoimmune encephalitis (EAE) is a well-established animal model of MS in which, in addition to demyelinating lesions and inflammatory reactions (56, 57), cognitive disturbances are also evident within EAE albeit (58) not in the early phase of this model (56, 59). Herbal medicine as an alternative or complementary therapy protocol for MS has been shown to enhance the efficacy of current treatments (60-62).
A recent study showed that oral administration of aqueous extract of Achillea (40, 200 and 400 mg/kg) attenuated the disease severity in combination with inflammatory responses, and demyelinating lesions in EAE model of MS in male C57BL/6 mice (11). An ongoing author-designed clinical trial is now assessing the effects of A. millefolium aqueous extract on MS patients by the authors of this manuscript.
In addition, for luteolin; Hendriks and colleagues reported that both oral and intraperitoneal administration of luteolin (100 mg/kg) in Lewis rat model at EAE suppressed behavioral deficits, prevented relapse and reduced inflammation, as well as axonal damage (13). Furthermost, it was recently reported that luteolin (10 μM) could improve maturation of oligodendrocyte precursor cells (OPCs) in tissue culture. These cells serve as myelin sheet producer around neurons in mice EAE model (14). However, in a different study it was reported that oral administration of luteolin (10 mg) delayed the recovery of behavioral defects rather than any reduction in disease severity in EAE female SJL/J mice (15). Further studies are requisite to be undertaken to clarify the effect(s) of luteolin in improvement of MS related damages.
For apigenin; it was reported that oral and intraperitoneal (IP) administration of apigenin (40 mg/kg of body weight) reduces the progression and relapse in two mouse models of MS (female C57/BL/6 and SJL/J mice) by modulating the immune system responses (12).
Anxiety disorders are prevalent and disabling conditions are usually accompanied by other neurological disorders, such as MS (63). There exist some reports in favor of sedative effects of Achillea and its flavonoid components; for example, injection of Achillea extract (100 mg/kg) in Wistar rats by intraperitoneal showed more sedative and anxiolytic effects than diazepam (2 mg/kg) (64). Further, chronic gavage (300-600 mg/kg) (65) or thereby being intraperitonealy injected (8, 10 or 12.0 mg/kg), Achillea extract showed anti-anxiety effects in albino Swiss male mice (66). Apigenin modulates the GABAergic system and show anxiolytic effect in rodents (67). Grundmann et al. demonstrated anxiolytic activity for kaempferol (0.02 to 1.0 mg/kg IP) and they also showed that this effect was partially antagonized by flumazenil (68). Despite the low affinity of luteolin for the GABAA receptors, it has anxiolytic-like effects. Coleta and coworkers showed that luteolin (0.1–50 mg/kg IP) increased exploration in the open areas and increased the number of entries in the open areas of the elevated plus maze, suggesting an anxiolytic-like effect in male adult Swiss mice (69). Also in another study Crupi colleagues investigated the effects of luteolin (1 mg/kg IP) in an animal model of anxiety/depression (129Sv/Ev mice) and reported similar results in elevated plus maze (70).
Alzheimer’s disease
The Alzheimer’s disease (AD) is described as one of the main subtypes of neurodegenerative diseases. The prominent hallmark of AD includes formation of Amyloid β (Aβ) plaques, degeneration of cholinergic neurons and neuronal cell death (71).
The effects of Achillea flavonoids have been previously evaluated in AD. A beneficial effect of luteolin on AD complications has drawn attentions during last two years. Cell culture studies demonstrated that luteolin (30, 27, 2.7 and 0.27 μM) inhibits Aβ-induced neuronal death in cultured mouse cortical cells and it had strong scavenging activities (20, 21). In line with these data, experimental studies using animal models of AD (male Wistar rats) demonstrated that luteolin (10 and 20 mg/kg) is able to inhibit memory impairment, prohibits degeneration of cholinergic neurons and amyloid deposition (22, 23). Brain injuries following trauma are recognized as a major risk factors for AD, and interestingly, the protective effect of luteolin (1.25, 2.5, 5 and 10 mg/kg/day) on AD complications was also found in traumatic brain injury-induced AD in Tg2576 mice (24). Considering beneficial effects of luteolin in animal models and in vitro studies, designing of clinical trial studies by luteolin is highly recommended.
Studies on the effects of apigenin in AD have longer history. In earlier studies it was reported that apigenin (25 μM and 50 μM) can inhibit activated microglial cells, which may contribute for creation of gliosis and neuronal death in AD-induced animal models (16, 17). It was also reported that apigenin (25 μM) could protect copper-mediated β-amyloid neurotoxicity in AD cell model (copper dyshomeostasis
is evident within AD-affected brain) (18). In, a later study, using animal model of AD (APP/PS1 mice) demonstrated that apigenin in a dose of 40 mg/kg could ameliorate AD-associated learning and memory deficits and relieve as well as formation of Aβ plaques (19).
The kaempferol is a polyphenol compound that was reported to inhibit formation, extension, and destabilization of beta-amyloid fibrils at a dose of 30 μl in Escherichia coli (E. coli). As a result, the fibril Aβ plaque was remodeled to non-toxic oligomers and fibrils (25, 72). In cell culture studies, it was demonstrated that kaempferol (25 mM) inhibit Aβ- induced toxicity in mouse-originated hippocampal neuronal cells (HT22) (26, 27). In vivo studies showed that kaempferol (10, 20 and 40 mg/kg) reversed memory impairment in mouse model of AD (28).
Parkinson’s disease
Parkinson’s disease (PD) is a progressive neuro-degenerative disorder characterized by injuries to dopaminergic neurons of the basal ganglia. Accumulating evidences have suggested that both inflammation and oxidative stress are involved in PD pathogenesis (73).
It was reported that gavage of A. millefolium aqueous extract (1.4 and 2.8 mg/kg) can improve motility disorders and muscle tone in rat models of PD (29).
Some studies have investigated the effects of Achillea flavonoids on PD. In a study, Wruck et al. reported that luteolin (3.13, 6.25, 12.5, 25 or 50 μM) was able to protect neural pheochromocytoma (PC12) and glial C6 cells against oxidative-medieted inturies in a rat cell line (32). Later, evidences demonstrated that luteolin (100 μM) could prohibit dopaminergic neurons death, raised by oxidant and inflammatory insults (PC12 cell line) (33, 74, 75). In accordance with in vitro results, in vivo studies also showed that gavage of luteolin (10 and 20 mg/kg) to mice model of PD ameliorated the locomotor and muscular activities and protected dopaminergic neurons (31). Interestingly, co-administration of luteolin with palmitoylethanolamide by a ratio of 1:10 potentiated the protective effect of luteolin on dopaminergic neurons in, in vivo models of PD (76).
In vitro studies indicated that apigenin (100 μM) inhibited the disaggregation of alpha-synuclein oligomers (77) and lipid membrane damage was also prevented by this protein (30). During recent years, studies demonstrated that alpha-synuclein protein plays a fundamental part in pathophysiology of PD. As a matter of fact, increased expression and/or pathologically altered forms of this protein could degenerate nigrostriatal dopaminergic neurons (78). Results of a new in vivo study revealed that gavage of apigenin (5, 10 and 20 mg/kg) to mice model of PD has improved the locomotor and muscular activity along with protection of dopaminergic neurons (31).
There also exist positive reports regarding the effect of kaempferol (20 mM) on PD. It has been shown that this compound was able to prevent oxidative stress-induced cell death in rat PC12 and primary neuronal cells, in in vitro cell culture studies (34) and enhances mitochondrial turnover by autophagy (79). Interestingly, it appears that the antioxidant properties of kaempferol are mediated through DJ-1 dependent pathways (34). DJ-1 is a causative gene product that plays a role in anti-oxidative stress, and its dysfunctional state would result in the onset of PD (80, 81). In line with the in vitro studies, Li and Pu, reported that kaempferol (25, 50 and 100 mg/kg) ameliorated motor coordination, elevated striatal dopamine level and prohibited the loss of dopaminergic neurons in male C57BL/6 mice model of PD (35).
In conclusion, both Achillea and its three flavonoid contents have shown anti-parkinson properties. Altogether this may evolve a new avenue to the treatment of neurodegenerative disorders or at least, partially be considered as a promising target for reduction of harmful clinical symptoms.
Stroke
It was reported that oral administration of the aqueous extract of Achillea (50, 500 mg/kg) in female rats subjected to ovariectomy, caused a significantly reduced the infarct volume, neurological disorders and sensorimotor disorder in middle cerebral artery occlusion in animal models of stroke (36).
Studies about the possible beneficial effects of luteolin in stroke were initially come from two interesting human studies. Marniemi et al. showed that high dietary intake of the two flavonoids such as, luteolin and kaempferol (as supplemental nutrient at least once a week) has attenuated risk of stroke (39). Holt and colleagues also reported that serum levels of inflammatory markers were inversely associated with the dietary input of luteolin (40). Cell culture studies demonstrated that luteolin (there by used as an aqueous solution of 500 ml) has exerted a neuroprotective effect on reactive oxygen species (ROS)-insulted primary neurons (41). Accordingly, in vivo studies showed that luteolin (5 and 20 mg/kg) alleviates neurological deficit, decreases infarct volume and suppresses edema in rat stroke models (41-43). A recently published clinical trial cohort study demonstrated that the stroke patients treated with co-ultramicronized palmitoylethanolamide (PEA)/luteolin (contained co-ultramicronized 700 mg PEA and 70 mg luteolin, in microgranular form) preparation for 60 days, have improved neurological status, cognitive abilities and independence in daily living activities (44).
Studies about the effects of apigenin on stroke emerged to the literature since eight years ago.
Firstly, Ha and colleagues reported that apigenin (1, 5, and 10 mM) had anti-inflammatory effect through inhibition of activated murine microglia cell line (BV-2 cells). They also demonstrated that this flavonoid decreased the infarct volume in an animal model of stroke (37). Interestingly, it is likely that stroke also induces astrocytes impairment and apigenin (10, 30, or 50 μM) could inhibit these pathologic state in astrocytes which were obtained from stroke-prone
spontaneously hypertensive rats (especially the expression of adhesion molecules) (82). Recent evidences, also reported that apigenin (1, 10, 20 and 40 μM) had neuroprotective effects against reactive oxygen species (ROS)-induced insalts in primary neurons (in cultured PC12 Cells) (38). Taken together, these results may suggest that apigenin has protective effects on both glial and neuronal cells in stroke.
Two various in vivo studies demonstrated that further acute ischemia administration of kaempferol (injection, at a dose of 10–15 μM/l of blood) prevented ischemic brain injuries as well as neuroinflammation in rats (45, 46).
Epilepsy
Epilepsy is a frequent, seriously neurological disorder, worldwide. Oxidative processes and free radicals both play pivotal roles in pathogenesis of epilepsy.
It has been established that hydro-alcoholic extract of Achillea (100, 200 or 400 mg/kg) was able to decrease seizure severity in animal models (male Wistar rat epilepsy model) (47).
Reports about the effect of luteolin in epilepsy are controversial. As an instance Zhen et al. studied the effect of luteolin in pentylenetetrazole induced epileptic male Sprague–Dawley rats and that oral administration of luteolin (50 or 100 mg/kg/day) suppressed the seizure intensity and duration in this animal model of epilepsy (50). Interestingly, the anticonvulsant effects of luteolin may be due to its inhibitory effects on neuronal and oxidative stress damages which are evident in chronic seizure activities (50). Alternatively, it has been shown that luteolin (3 and 10 mg/kg) can bind the benzodiaze-pine site on the GABAA receptors on the surface of BV-2 microglia-like cells (83). However, in an earlier study, Shaikh and colleagues examined the effect of luteolin in both acute and chronic animal models of seizure. In this set of discrepant data they failed to find any indications that luteolin (3 and 10 mg/kg) is pro- or anti-convulsant neither in chronic nor in acute mouse models (CD1 mice) of seizure (84). These discrepant findings may be due to various doses of luteolin used by two different studies (50 - 100 mg/kg in first study vs. 3-10 mg/kg in last study).
The effect of luteolin on epilepsy was as similar as apigenin disputing. In an early study, it was reported that apigenin (there by was IP injected in a 25 and 50 mg/kg dose) reduced the latency in the onset of picrotoxin-induced convulsions. This effect was attributed to the inhibitory effects of apigenin on GABA activated Cl(-) currents on cerebellar granule cells (48). Later, Campbell and co-workers demons-trated a novel second order modulatory action of apigenin on GABA recombinant human α1β2γ2L GABAA receptors. Actually, they showed that apigenin (1 μM) could enhance GABA responses to diazepam (85). In accordance to these results, it has been proposed that apigenin might also potentiate the effect of the endogenous benzodiazepine-like agents like endozepines which can be used clinically to control convulsions (85). A recent study demonstrated a protective effect for apigenin (at 25 and 50 mg/kg dose) in Kainic acid-induced seizure in male ICR in mouse model. This antiepileptic effect of apigenin is associated with the inhibition of Kainic acid-induced cytotoxicity in neurons (49).
Juan Song et al. showed the anti-epileptic effects of kaempferol (8 and 30 μM) in a genetic model of epilepsy in Drosophila (51). The anti-epileptic activity of kaempferol may be due to its effects on inhibition of DNA topoisomerase I enzyme. This could be especially beneficial for brain tumor patients who are suffering from epilepsy. Nugroho et al. showed the anticonvul-sant effects of kaempferol (per os (p.o) injection at 100 and 200 mg/kg dose) in extract of Aster glehni on pentylenetetrazole-induced seizure in ICR mouse model (52).
Conclusion
There is an extensive body of evidences on beneficial effects of Achillea and its main flavonoids on nervous system related disorders within the database. However, almost all of performed studies have focused on in vitro and animal preparations. Also, almost all of these in vivo and in vitro reports showed beneficial preventive or therapeutic properties of Achillea and/or its flavonoids on epilepsy, Alzheimer’ diseases, multiple sclerosis, Parkinson’ diseases and stroke (Figure 3). Considering little side effects of Achillea and its derivative flavonoids (86) on main brain functions (87, 88), as well as their availability for supplemental use, one can suggest to use them in neurodegenerative disorders.
Figure 3.
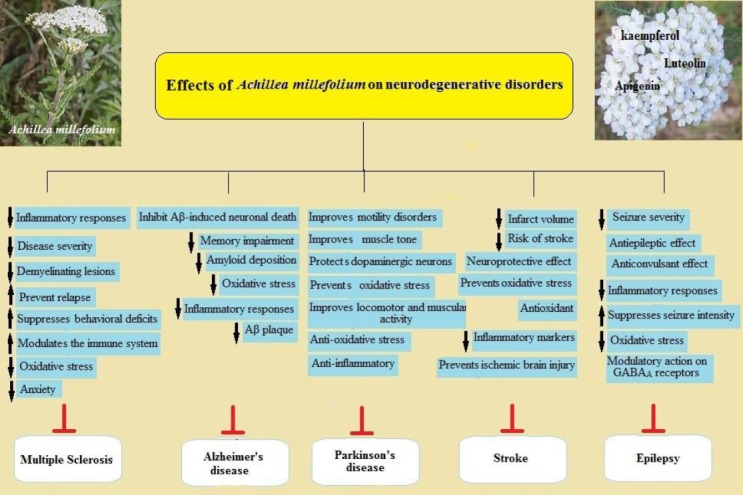
Schematic description for protective mechanisms of Achillea millefolium L. in ameliorating neurodegenerative diseases
As tried our best, we could not find any further clinical trial study about the effects of Achillea and/or its three flavonoids in nervous system related disorders. Future studies are required to address a possible preventative/therapeutic role for Achillea and/or its flavonoids on neurodegenerative diseases with focusing on human subjects.
Conflict of interest
The authors have declared that no conflict of interest exists.
References
- 1.Applequist WL, Moerman DE. Yarrow (Achillea millefolium L.): a neglected panacea? A review of ethnobotany, bioactivity, and biomedical research. Econ Bot. 2011;65:209–225. [Google Scholar]
- 2.Kowal T, Pic S. Produktywnośćgatunku Achillea millefolium L, w warunkach naturalnych [Productivity of the species Achillea millejolium L. in natural habitats] Acta Agrobot. 2015;32:91–100. [Google Scholar]
- 3.Stojanović G, Radulović N, Hashimoto T, Palić R. In vitro antimicrobial activity of extracts of four Achillea species: The composition of Achillea clavennae L.(Asteraceae) extract. J Ethnopharmacol. 2005;101:185–190. doi: 10.1016/j.jep.2005.04.026. [DOI] [PubMed] [Google Scholar]
- 4.Benedek B, Kopp B, Melzig MF. Achillea millefolium L. sl–Is the anti-inflammatory activity mediated by protease inhibition? J Ethnopharmacol. 2007;113:312–317. doi: 10.1016/j.jep.2007.06.014. [DOI] [PubMed] [Google Scholar]
- 5.Mahady GB, Pendland SL, Stoia A, Hamill FA, Fabricant D, Dietz BM, et al. In vitro susceptibility of Helicobacter pylori to botanical extracts used traditionally for the treatment of gastrointestinal disorders. Phytother Res. 2005;19:988–991. doi: 10.1002/ptr.1776. [DOI] [PubMed] [Google Scholar]
- 6.Lemmens-Gruber R, Marchart E, Rawnduzi P, Engel N, Benedek B, Kopp B. Investigation of the spasmolytic activity of the flavonoid fraction of Achillea millefolium sl on isolated guinea-pig ilea. Arzneimittelforschung. 2005;56:582–588. doi: 10.1055/s-0031-1296755. [DOI] [PubMed] [Google Scholar]
- 7.Ivanov Ch L. Y. Composition of Achillea millefolium. III. Composition of the acidic, water-insoluble part of the alcoholic extract. God Vissh Khimikotekhnol Inst Sofia. 1967;14:61–72. [Google Scholar]
- 8.Ivanov Ch L. Y. Composition of Achillea millefolium. V. Composition and structure of the components of neutral fraction insoluble in the aqueous part of the alcoholic extract. God Vissh Khimikotekhnol Inst Sofia. 1967;14:73–101. [Google Scholar]
- 9.Chandler R, Hooper S, Harvey MJ. Ethnobotany and phytochemistry of yarrow, Achillea millefolium, Compositae. Econ Bot. 1982;36:203–223. [Google Scholar]
- 10.Falk A, Smolenski S, Bauer L, Bell C. Isolation and identification of three new flavones from Achillea millefolium L. J Pharm Sci. 1975;64:1838–1842. doi: 10.1002/jps.2600641119. [DOI] [PubMed] [Google Scholar]
- 11.Vazirinejad R, Ayoobi F, Arababadi MK, Eftekharian MM, Darekordi A, Goudarzvand M, et al. Effect of aqueous extract of Achillea millefolium on the development of experimental autoimmune encephalomyelitis in C57BL/6 mice. Indian J Pharm. 2014;46:303. doi: 10.4103/0253-7613.132168. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 12.Ginwala R, McTish E, Raman C, Singh N, Nagarkatti M, Nagarkatti P, et al. Apigenin, a Natural Flavonoid, Attenuates EAE Severity Through the Modulation of Dendritic Cell and Other Immune Cell Functions. J Neuroimmune Pharmacol. 2016;11:36–47. doi: 10.1007/s11481-015-9617-x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 13.Hendriks JJ, Alblas J, van der Pol SM, van Tol EA, Dijkstra CD, de Vries HE. Flavonoids influence monocytic GTPase activity and are protective in experimental allergic encephalitis. J Exp Med. 2004;200:1667–1672. doi: 10.1084/jem.20040819. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 14.Barbierato M, Facci L, Marinelli C, Zusso M, Argentini C, Skaper SD, et al. Co-ultramicronized Palmitoylethanolamide/Luteolin Promotes the Maturation of Oligodendrocyte Precursor Cells. Sci Rep. 2015;5:16676. doi: 10.1038/srep16676. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 15.Verbeek R, van Tol EA, van Noort JM. Oral flavonoids delay recovery from experimental autoimmune encephalomyelitis in SJL mice. Biochem Pharmacol. 2005;70:220–228. doi: 10.1016/j.bcp.2005.04.041. [DOI] [PubMed] [Google Scholar]
- 16.Elsisi NS, Darling-Reed S, Lee EY, Oriaku ET, Soliman KF. Ibuprofen and apigenin induce apoptosis and cell cycle arrest in activated microglia. Neurosci Lett. 2005;375:91–96. doi: 10.1016/j.neulet.2004.10.087. [DOI] [PubMed] [Google Scholar]
- 17.Rezai-Zadeh K, Ehrhart J, Bai Y, Sanberg PR, Bickford P, Tan J, et al. Apigenin and luteolin modulate microglial activation via inhibition of STAT1-induced CD40 expression. J Neuroinflamm. 2008;5:41. doi: 10.1186/1742-2094-5-41. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 18.Zhao L, Wang JL, Wang YR, Fa XZ. Apigenin attenuates copper-mediated beta-amyloid neurotoxicity through antioxidation, mitochondrion protection and MAPK signal inactivation in an AD cell model. Brain Res. 2013;1492:33–45. doi: 10.1016/j.brainres.2012.11.019. [DOI] [PubMed] [Google Scholar]
- 19.Zhao L, Wang JL, Liu R, Li XX, Li JF, Zhang L. Neuroprotective, anti-amyloidogenic and neurotrophic effects of apigenin in an Alzheimer’s disease mouse model. Molecules. 2013;18:9949–9965. doi: 10.3390/molecules18089949. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 20.Choi SM, Kim BC, Cho YH, Choi KH, Chang J, Park MS, et al. Effects of Flavonoid Compounds on beta-amyloid-peptide-induced Neuronal Death in Cultured Mouse Cortical Neurons. Chonnam Med J. 2014;50:45–51. doi: 10.4068/cmj.2014.50.2.45. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 21.Paterniti I, Cordaro M, Campolo M, Siracusa R, Cornelius C, Navarra M, et al. Neuroprotection by association of palmitoylethanolamide with luteolin in experimental Alzheimer’s disease models: the control of neuroinflammation. CNS Neurol Disord Drug Targets. 2014;13:1530–1541. doi: 10.2174/1871527313666140806124322. [DOI] [PubMed] [Google Scholar]
- 22.Yu TX, Zhang P, Guan Y, Wang M, Zhen MQ. Protective effects of luteolin against cognitive impairment induced by infusion of Abeta peptide in rats. Int J Clin Exp Pathol. 2015;8:6740–6747. [PMC free article] [PubMed] [Google Scholar]
- 23.Wang H, Wang H, Cheng H, Che Z. Ameliorating effect of luteolin on memory impairment in an Alzheimer’s disease model. Mol Med Rep. 2016;13:4215–4220. doi: 10.3892/mmr.2016.5052. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 24.Sawmiller D, Li S, Shahaduzzaman M, Smith AJ, Obregon D, Giunta B, et al. Luteolin reduces Alzheimer’s disease pathologies induced by traumatic brain injury. Int J Mol Sci. 2014;15:895–904. doi: 10.3390/ijms15010895. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 25.Sharoar MG, Thapa A, Shahnawaz M, Ramasamy VS, Woo ER, Shin SY, et al. Keampferol-3-O-rhamnoside abrogates amyloid beta toxicity by modulating monomers and remodeling oligomers and fibrils to non-toxic aggregates. J Biomed Sci. 2012;19:104. doi: 10.1186/1423-0127-19-104. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 26.Yang EJ, Kim GS, Jun M, Song KS. Kaempferol attenuates the glutamate-induced oxidative stress in mouse-derived hippocampal neuronal HT22 cells. Food Funct. 2014;5:1395–1402. doi: 10.1039/c4fo00068d. [DOI] [PubMed] [Google Scholar]
- 27.Roth A, Schaffner W, Hertel C. Phytoestrogen kaempferol (3,4’,5,7-tetrahydroxyflavone) protects PC12 and T47D cells from beta-amyloid-induced toxicity. J Neurosci Res. 1999;57:399–404. [PubMed] [Google Scholar]
- 28.Kim JK, Choi SJ, Cho HY, Hwang HJ, Kim YJ, Lim ST, et al. Protective effects of kaempferol (3,4’,5,7-tetrahydroxyflavone) against amyloid beta peptide (Abeta)-induced neurotoxicity in ICR mice. Biosci Biotechnol Biochem. 2010;74:397–401. doi: 10.1271/bbb.90585. [DOI] [PubMed] [Google Scholar]
- 29.Akramian Fard M, Moghadam Ahmadi A, Ayoobi F, Nakisa H, Hadadian Z, Shabani M, et al. Effects of Achillea millefolium aqueous extract in a Parkinsons diaease model induced by intra-cerebralventricular injection of 6-hydroxydopamine in male rats. Quarterly J Sabzevar Univ Med Sci. 2015:22. [Google Scholar]
- 30.Caruana M, Neuner J, Hogen T, Schmidt F, Kamp F, Scerri C, et al. Polyphenolic compounds are novel protective agents against lipid membrane damage by alpha-synuclein aggregates in vitro. Biochim Biophys Acta. 2012;1818:2502–2510. doi: 10.1016/j.bbamem.2012.05.019. [DOI] [PubMed] [Google Scholar]
- 31.Patil SP, Jain PD, Sancheti JS, Ghumatkar PJ, Tambe R, Sathaye S. Neuroprotective and neurotrophic effects of Apigenin and Luteolin in MPTP induced parkinsonism in mice. Neuropharmacology. 2014;86:192–202. doi: 10.1016/j.neuropharm.2014.07.012. [DOI] [PubMed] [Google Scholar]
- 32.Wruck CJ, Claussen M, Fuhrmann G, Romer L, Schulz A, Pufe T, et al. Luteolin protects rat PC12 and C6 cells against MPP+induced toxicity via an ERK dependent Keap1-Nrf2-ARE pathway. J Neural Transm Suppl. 2007;72:57–67. doi: 10.1007/978-3-211-73574-9_9. [DOI] [PubMed] [Google Scholar]
- 33.Lin YP, Chen TY, Tseng HW, Lee MH, Chen ST. Chemical and biological evaluation of nephrocizin in protecting nerve growth factor-differentiated PC12 cells by 6-hydroxydopamine-induced neurotoxicity. Phytochemistry. 2012;84:102–115. doi: 10.1016/j.phytochem.2012.07.020. [DOI] [PubMed] [Google Scholar]
- 34.Qu W, Fan L, Kim YC, Ishikawa S, Iguchi-Ariga SM, Pu XP, et al. Kaempferol derivatives prevent oxidative stress-induced cell death in a DJ-1-dependent manner. J Pharmacol Sci. 2009;110:191–200. doi: 10.1254/jphs.09045fp. [DOI] [PubMed] [Google Scholar]
- 35.Li S, Pu XP. Neuroprotective effect of kaempferol against a 1-methyl-4-phenyl-1,2,3,6-tetrahydropyridine-induced mouse model of Parkinson’s disease. Biol Pharm Bull. 2011;34:1291–1296. doi: 10.1248/bpb.34.1291. [DOI] [PubMed] [Google Scholar]
- 36.Imani E, Esmaili A, Alimohammadi R, Ehsani V, Shamsizadeh A, Mobini M, et al. Effects of Achillea.millefolium on the Consequences of Stroke in Ovarectomized Rats. J Shahid Sadoughi Univ Med Sci. 2015;22:1725–1736. [Google Scholar]
- 37.Ha SK, Lee P, Park JA, Oh HR, Lee SY, Park JH, et al. Apigenin inhibits the production of NO and PGE2 in microglia and inhibits neuronal cell death in a middle cerebral artery occlusion-induced focal ischemia mice model. Neurochem Int. 2008;52:878–886. doi: 10.1016/j.neuint.2007.10.005. [DOI] [PubMed] [Google Scholar]
- 38.Guo H, Kong S, Chen W, Dai Z, Lin T, Su J, et al. Apigenin mediated protection of OGD-evoked neuron-like injury in differentiated PC12 cells. Neurochem Res. 2014;39:2197–2210. doi: 10.1007/s11064-014-1421-0. [DOI] [PubMed] [Google Scholar]
- 39.Marniemi J, Alanen E, Impivaara O, Seppanen R, Hakala P, Rajala T, et al. Dietary and serum vitamins and minerals as predictors of myocardial infarction and stroke in elderly subjects. Nutr Metab Cardiovasc Dis. 2005;15:188–197. doi: 10.1016/j.numecd.2005.01.001. [DOI] [PubMed] [Google Scholar]
- 40.Holt EM, Steffen LM, Moran A, Basu S, Steinberger J, Ross JA, et al. Fruit and vegetable consumption and its relation to markers of inflammation and oxidative stress in adolescents. J Am Diet Assoc. 2009;109:414–421. doi: 10.1016/j.jada.2008.11.036. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 41.Zhao G, Zang S-Y, Jiang Z-H, Chen Y-Y, Ji X-H, Lu B-F, et al. Postischemic administration of liposome-encapsulated luteolin prevents against ischemia-reperfusion injury in a rat middle cerebral artery occlusion model. J Nutr Biochem. 2011;22:929–936. doi: 10.1016/j.jnutbio.2010.07.014. [DOI] [PubMed] [Google Scholar]
- 42.Qiao H, Dong L, Zhang X, Zhu C, Zhang X, Wang L, et al. Protective effect of luteolin in experimental ischemic stroke: upregulated SOD1, CAT, Bcl-2 and claudin-5, down-regulated MDA and Bax expression. Neurochem Res. 2012;37:2014–2024. doi: 10.1007/s11064-012-0822-1. [DOI] [PubMed] [Google Scholar]
- 43.Qiao H, Zhang X, Zhu C, Dong L, Wang L, Zhang X, et al. Luteolin downregulates TLR4, TLR5, NF-κB and p-p38MAPK expression, upregulates the p-ERK expression, and protects rat brains against focal ischemia. Brain Res. 2012;1448:71–81. doi: 10.1016/j.brainres.2012.02.003. [DOI] [PubMed] [Google Scholar]
- 44.Caltagirone C, Cisari C, Schievano C, Di Paola R, Cordaro M, Bruschetta G, et al. Co-ultramicronized Palmitoylethanolamide/Luteolin in the Treatment of Cerebral Ischemia: from Rodent to Man. Transl Stroke Res. 2016;7:54–69. doi: 10.1007/s12975-015-0440-8. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 45.Lopez-Sanchez C, Martin-Romero FJ, Sun F, Luis L, Samhan-Arias AK, Garcia-Martinez V, et al. Blood micromolar concentrations of kaempferol afford protection against ischemia/reperfusion-induced damage in rat brain. Brain Res. 2007;1182:123–137. doi: 10.1016/j.brainres.2007.08.087. [DOI] [PubMed] [Google Scholar]
- 46.Yu L, Chen C, Wang LF, Kuang X, Liu K, Zhang H, et al. Neuroprotective effect of kaempferol glycosides against brain injury and neuroinflammation by inhibiting the activation of NF-kappaB and STAT3 in transient focal stroke. PLoS One. 2013;8:e55839. doi: 10.1371/journal.pone.0055839. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 47.Hosseini M, Harandizadeh F, Niazamand S, Soukhtanloo M, Mahmoudabady M. Antioxidant effect of Achillea wilhelmsii extract on pentylenetetrazole (seizure model)-induced oxidative brain damage in Wistar rats. Indian J Physiol Pharmacol. 2013;57:418–424. [PubMed] [Google Scholar]
- 48.Avallone R, Zanoli P, Puia G, Kleinschnitz M, Schreier P, Baraldi M. Pharmacological profile of apigenin, a flavonoid isolated from Matricaria chamomilla. Biochem Pharm. 2000;59:1387–1394. doi: 10.1016/s0006-2952(00)00264-1. [DOI] [PubMed] [Google Scholar]
- 49.Han J-Y, Ahn S-Y, Kim C-S, Yoo S-K, Kim S-K, Kim H-C, et al. Protection of apigenin against kainate-induced excitotoxicity by anti-oxidative effects. Biol Pharm Bull. 2012;35:1440–1446. doi: 10.1248/bpb.b110686. [DOI] [PubMed] [Google Scholar]
- 50.Zhen J-L, Chang Y-N, Qu Z-Z, Fu T, Liu J-Q, Wang W-P. Luteolin rescues pentylenetetrazole-induced cognitive impairment in epileptic rats by reducing oxidative stress and activating PKA/CREB/BDNF signaling. Epilepsy Behav. 2016;57:177–184. doi: 10.1016/j.yebeh.2016.02.001. [DOI] [PubMed] [Google Scholar]
- 51.Song J, Parker L, Hormozi L, Tanouye MA. DNA topoisomerase I inhibitors ameliorate seizure-like behaviors and paralysis in a Drosophila model of epilepsy. Neuroscience. 2008;156:722–728. doi: 10.1016/j.neuroscience.2008.07.024. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 52.Nugroho A, Kim MH, Choi J, Choi JS, Jung WT, Lee KT, et al. Phytochemical studies of the phenolic substances in Aster glehni extract and its sedative and anticonvulsant activity. Arch Pharm Res. 2012;35:423–430. doi: 10.1007/s12272-012-0304-7. [DOI] [PubMed] [Google Scholar]
- 53.Khorramdelazad H, Bagheri V, Hassanshahi G, Zeinali M, Vakilian A. New insights into the role of stromal cell-derived factor 1 (SDF-1/CXCL12) in the pathophysiology of multiple sclerosis. J Neuroimmunol. 2016;290:70–75. doi: 10.1016/j.jneuroim.2015.11.021. [DOI] [PubMed] [Google Scholar]
- 54.Noroozi Karimabad M, Khanamani Falahati-Pour S, Hassanshahi G. Significant Role(s) of CXCL12 and the SDF-1 3’A Genetic Variant in the Pathogenesis of Multiple Sclerosis. Neuroimmunomodulation. 2016;23:197–208. doi: 10.1159/000449427. [DOI] [PubMed] [Google Scholar]
- 55.Vazirinejad R, Ahmadi Z, Kazemi Arababadi M, Hassanshahi G, Kennedy D. The biological functions, structure and sources of CXCL10 and its outstanding part in the pathophysiology of multiple sclerosis. Neuroimmunomodulation. 2014;21:322–330. doi: 10.1159/000357780. [DOI] [PubMed] [Google Scholar]
- 56.Fatemi I, Shamsizadeh A, Ayoobi F, Taghipour Z, Sanati MH, Roohbakhsh A, et al. Role of orexin-A in experimental autoimmune encephalomyelitis. J Neuroimmunol. 2016;291:101–109. doi: 10.1016/j.jneuroim.2016.01.001. [DOI] [PubMed] [Google Scholar]
- 57.Fatemi I, Shamsizadeh A, Roohbakhsh A, Ayoobi F, Sanati MH, Motevalian M. Increased mRNA Level of Orexin1 and 2 Receptors Following Induction of Experimental Autoimmune Encephalomyelitis in Mice. Iran J Allergy Asthma Immunol. 2016;15:20. [PubMed] [Google Scholar]
- 58.Hollinger KR, Alt J, Riehm AM, Slusher BS, Kaplin AI. Dose-dependent inhibition of GCPII to prevent and treat cognitive impairment in the EAE model of multiple sclerosis. Brain Res. 2016;1635:105–112. doi: 10.1016/j.brainres.2016.01.035. [DOI] [PubMed] [Google Scholar]
- 59.Ayoobi F, Fatemi I, Roohbakhsh A, Shamsizadeh A. Tactile learning within the early phase of experimental autoimmune encephalomyelitis in mice. Neurophysiology. 2013;45:306–311. [Google Scholar]
- 60.Jafarzadeh A, Mohammadi-Kordkhayli M, Ahangar-Parvin R, Azizi V, Khoramdel-Azad H, Shamsizadeh A, et al. Ginger extracts influence the expression of IL-27 and IL-33 in the central nervous system in experimental autoimmune encephalomye-litis and ameliorates the clinical symptoms of disease. J Neuroimmunology. 2014;276:80–88. doi: 10.1016/j.jneuroim.2014.08.614. [DOI] [PubMed] [Google Scholar]
- 61.Jafarzadeh A, Ahangar-Parvin R, Azizi S, Ayobi F, Taghipour Z, Shamsizadeh A, et al. Evaluation of the Effect of Vitamin D3 and Ginger Extract on the Clinical Symptoms and the Severity of Inflammation in EAE. J Rafsanjan Univ Med Sci. 2015;13:683–694. [Google Scholar]
- 62.Jafarzadeh A, Azizi S-V, Nemati M, Khoramdel-Azad H, Shamsizadeh A, Ayoobi F, et al. Ginger Extract Reduces the Expression of IL-17 and IL-23 in the Seraand Central Nervous System of EAEMice. Iran J Immunol. 2015;12:288. doi: 10.22034/iji.2015.16757. [DOI] [PubMed] [Google Scholar]
- 63.Association AP. Diagnostic and Statistical Manual of Mental Disorders: DSM-5: ManMag. 2003 [Google Scholar]
- 64.Rezaie Ali Changhiz. A Study of sedation, pre-anesthetic and anti-anxiety effects of polar, semi-polar and non-polar fractions of yarrow (Achillea millefolium) extract compared with Diazepam in rats. Life Sci J. 2013:10. [Google Scholar]
- 65.Irinéia Paulina Barettaa, Regiane Américo Felizardoa, Vanessa Fávero Bimbatoa, Gonc¸ M, alves Jorge dos Santosa, Candida Aparecida Leite Kassuyab, et al. Anxiolytic-like effects of acute and chronic treatment with Achillea millefolium L. extract. J Ethnopharmacol. 2012:46–54. doi: 10.1016/j.jep.2011.11.047. [DOI] [PubMed] [Google Scholar]
- 66.Molina-Hernandez M, Tellez-Alcantara NP, Diaz MA, Perez Garcia J, Olivera Lopez JI, Jaramillo MT. Anticonflict actions of aqueous extracts of flowers of A millefolium L. vary according to the estrous cycle phases in Wistar rats. Phytother Res. 2004;18:915–920. doi: 10.1002/ptr.1578. [DOI] [PubMed] [Google Scholar]
- 67.Herrera-Ruiz M, Román-Ramos R, Zamilpa A, Tortoriello J, Jiménez-Ferrer JE. Flavonoids from Tilia americana with anxiolytic activity in plus-maze test. J Ethnopharmacol. 2008;118:312–317. doi: 10.1016/j.jep.2008.04.019. [DOI] [PubMed] [Google Scholar]
- 68.Grundmann O, Nakajima J-I, Kamata K, Seo S, Butterweck V. Kaempferol from the leaves of Apocynum venetum possesses anxiolytic activities in the elevated plus maze test in mice. Phytomedicine. 2009;16:295–302. doi: 10.1016/j.phymed.2008.12.020. [DOI] [PubMed] [Google Scholar]
- 69.Coleta M, Campos MG, Cotrim MD, de Lima TCM, da Cunha AP. Assessment of luteolin (3′, 4′, 5, 7-tetrahydroxyflavone) neuropharmacological activity. Behav Brain Res. 2008;189:75–82. doi: 10.1016/j.bbr.2007.12.010. [DOI] [PubMed] [Google Scholar]
- 70.Crupi R, Paterniti I, Ahmad A, Campolo M, Esposito E, Cuzzocrea S. Effects of palmitoylethanol-amide and luteolin in an animal model of anxiety/depression. CNS Neurol Disord Drug Targets. 2013;12:989–1001. doi: 10.2174/18715273113129990084. [DOI] [PubMed] [Google Scholar]
- 71.Glenner GG. Reprint of “Alzheimer’s disease: Initial report of the purification and characterization of a novel cerebrovascular amyloid protein”. Biochem Biophys Res Commun. 2012;425:534–539. doi: 10.1016/j.bbrc.2012.08.020. [DOI] [PubMed] [Google Scholar]
- 72.Ono K, Yoshiike Y, Takashima A, Hasegawa K, Naiki H, Yamada M. Potent anti-amyloidogenic and fibril-destabilizing effects of polyphenols in vitro: implications for the prevention and therapeutics of Alzheimer’s disease. J Neurochem. 2003;87:172–181. doi: 10.1046/j.1471-4159.2003.01976.x. [DOI] [PubMed] [Google Scholar]
- 73.NCCfCC. Parkinson’s disease: National clinical guideline for diagnosis and management in primary and secondary care. Royal College of Physicians. 2006 [PubMed] [Google Scholar]
- 74.Guo DJ, Li F, Yu PH, Chan SW. Neuroprotective effects of luteolin against apoptosis induced by 6-hydroxydopamine on rat pheochromocytoma PC12 cells. Pharm Biol. 2013;51:190–196. doi: 10.3109/13880209.2012.716852. [DOI] [PubMed] [Google Scholar]
- 75.Hu LW, Yen JH, Shen YT, Wu KY, Wu MJ. Luteolin modulates 6-hydroxydopamine-induced transcrip-tional changes of stress response pathways in PC12 cells. PLoS One. 2014;9:e97880. doi: 10.1371/journal.pone.0097880. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 76.Siracusa R, Paterniti I, Impellizzeri D, Cordaro M, Crupi R, Navarra M, et al. The Association of Palmitoylethanolamide with Luteolin Decreases Neuroinflammation and Stimulates Autophagy in Parkinson’s Disease Model. CNS Neurol Disord Drug Targets. 2015;14:1350–1365. doi: 10.2174/1871527314666150821102823. [DOI] [PubMed] [Google Scholar]
- 77.Caruana M, Hogen T, Levin J, Hillmer A, Giese A, Vassallo N. Inhibition and disaggregation of alpha-synuclein oligomers by natural polyphenolic compounds. FEBS Lett. 2011;585:1113–1120. doi: 10.1016/j.febslet.2011.03.046. [DOI] [PubMed] [Google Scholar]
- 78.Lee VM, Trojanowski JQ. Mechanisms of Parkinson’s disease linked to pathological alpha-synuclein: new targets for drug discovery. Neuron. 2006;52:33–38. doi: 10.1016/j.neuron.2006.09.026. [DOI] [PubMed] [Google Scholar]
- 79.Filomeni G, Graziani I, De Zio D, Dini L, Centonze D, Rotilio G, et al. Neuroprotection of kaempferol by autophagy in models of rotenone-mediated acute toxicity: possible implications for Parkinson’s disease. Neurobiol Aging. 2012;33:767–785. doi: 10.1016/j.neurobiolaging.2010.05.021. [DOI] [PubMed] [Google Scholar]
- 80.Ariga H, Takahashi-Niki K, Kato I, Maita H, Niki T, Iguchi-Ariga SM. Neuroprotective function of DJ-1 in Parkinson’s disease. Oxid Med Cell Longev. 2013;2013:683920. doi: 10.1155/2013/683920. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 81.Akbari B, Gharanfoli F, Khayyat MH, Khashyarmanesh Z, Rezaee R, Karimi G. Determination of heavy metals in different honey brands from Iranian markets. Food Addit Contam Part B Surveill. 2012;5(2):105–11. doi: 10.1080/19393210.2012.664173. [DOI] [PubMed] [Google Scholar]
- 82.Yamagata K, Kitazawa T, Shinoda M, Tagawa C, Chino M, Matsufuji H. Stroke status evoked adhesion molecule genetic alterations in astrocytes isolated from stroke-prone spontaneously hypertensive rats and the apigenin inhibition of their expression. Stroke Res Treat. 2010:2010. doi: 10.4061/2010/386389. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 83.Dirscherl K, Karlstetter M, Ebert S, Kraus D, Hlawatsch J, Walczak Y, et al. Luteolin triggers global changes in the microglial transcriptome leading to a unique anti-inflammatory and neuroprotective phenotype. J Neuroinflammation. 2010;7:1742–2094. doi: 10.1186/1742-2094-7-3. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 84.Shaikh MF, Tan KN, Borges K. Anticonvulsant screening of luteolin in four mouse seizure models. Neurosci Lett. 2013;550:195–199. doi: 10.1016/j.neulet.2013.06.065. [DOI] [PubMed] [Google Scholar]
- 85.Campbell EL, Chebib M, Johnston GA. The dietary flavonoids apigenin and (-)-epigallocatechin gallate enhance the positive modulation by diazepam of the activation by GABA of recombinant GABA A receptors. Biochemical pharmacology. 2004;68:1631–1638. doi: 10.1016/j.bcp.2004.07.022. [DOI] [PubMed] [Google Scholar]
- 86.Jager AK, Saaby L. Flavonoids and the CNS. Molecules. 2011;16:1471–1485. doi: 10.3390/molecules16021471. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 87.Ayoobi Fatemeh, Roohbakhsh Ali, Allahtavakoli Mohammad, Vazirinejad Reza, Rajabi Soodeh, Ali S. Achillea millefolium Aqueous Extract Does not Impair Recognition Memory in Mice. Trop J Pharm Res. 2013:12. [Google Scholar]
- 88.Salaria E, Shamsizadeh A, Noorbakhsh SM, Ayoobi F, Sheibani V, Oryan S. Effects of Achillea millefolium Aqueous Extract on Electrophysiological Properties of Rat Barrel Cortex Neurons. 2016;10:56–59. [Google Scholar]