

Abstract
Background
Arachidonic acid (ARA, C20:4, n-6), which belongs to the omega-6 series of polyunsaturated fatty acids and has a variety of biological activities, is commercially produced in Mortierella alpina. Dissolved oxygen or oxygen utilization efficiency is a critical factor for Mortierella alpina growth and arachidonic acid production in large-scale fermentation. Overexpression of the Vitreoscilla hemoglobin gene is thought to significantly increase the oxygen utilization efficiency of the cells.
Results
An optimized Vitreoscilla hemoglobin (VHb) gene was introduced into Mortierella alpina via Agrobacterium tumefaciens-mediated transformation. Compared with the parent strain, the VHb-expressing strain, termed VHb-20, grew faster under both limiting and non-limiting oxygen conditions and exhibited dramatic changes in cell morphology. Furthermore, VHb-20 produced 4- and 8-fold higher total lipid and ARA yields than those of the wild-type strain under a microaerobic environment. Furthermore, ARA production of VHb-20 was also 1.6-fold higher than that of the wild type under normal conditions. The results demonstrated that DO utilization was significantly increased by expressing the VHb gene in Mortierella alpina.
Conclusions
The expression of VHb enhances ARA and lipid production under both lower and normal dissolved oxygen conditions. This study provides a novel strategy and an engineered strain for the cost-efficient production of ARA.
Electronic supplementary material
The online version of this article (doi:10.1186/s12896-017-0388-8) contains supplementary material, which is available to authorized users.
Keywords: Arachidonic acid, Mortierella Alpina, Hemoglobin, Dissolved oxygen, Fermentation
Background
Arachidonic acid (ARA, 5,8,11,14-cis-eicosatetraenoic acid), belonging to the omega-6 series of polyunsaturated fatty acids (PUFAs), is not only a structural component of the cell membrane but also the biogenetic precursor of prostaglandins, leukotrienes, thromboxanes, and other eicosanoid hormones [1, 2]. Furthermore, ARA is widely used in health food, pharmacology, agriculture, cosmetics and other industries [3, 4]. Mortierella alpina, which has a high ARA content, is considered one of the best ARA-producing strains and has been used in industrial applications [5].
The growth of M. alpina is closely related to the dissolved oxygen, especially in high cell density fermentation. The fast growth of the mycelium can result in an apparent increase in broth viscosity. The mycelium can agglomerate easily and further decrease the efficiency of oxygen mass transfer [6, 7]. Therefore, improving the concentration of dissolved oxygen in the fermentation process or increasing the utilization of dissolved oxygen is key to improving the yield of ARA.
At present, there are various methods to improve the oxygen supply, for example, by increasing the stirring speed or ventilation, or by adding surfactants to increase the concentration of dissolved oxygen in the fermentation. However, these methods require either special equipment or high energy consumption [8, 9]. It has been shown that the expression of the bacterial (Vitreoscilla) hemoglobin (VHb) is an effective method to solve the problem of oxygen supply in the fermentation process because VHb can improve the efficiency of intracellular oxygen transport [10, 11]. VHb is an oxygen-binding protein produced by Vitreoscilla, which allows the Vitreoscilla (aerobic bacteria) to survive under limited oxygen conditions. Khosla and Bailey first cloned the vgb gene and successfully expressed it in E. coli [12].They demonstrated that the expression of VHb could promote cell growth, increase protein synthesis capacity under limited oxygen conditions [13, 14]. According to the characteristics of VHb, it has a good application, which has been successfully engineered into different bacteria, fungi, as well as some plants and animals, to improve cell growth and to increase the expression of exogenous protein and metabolite production [15, 16].
The metabolic engineering method also has been applied to improve the ARA production. For example, Hao et al. [17] studied overexpression of ME2 and G6PD2 in M. alpina, increasing the supply of intracellular NADPH, with a 1.7-fold increase in total fatty acid and a 1.5-fold increase in arachidonic acid (ARA) content. In 2005, Takeno et al. [18] improved of the fatty acid composition of M. alpina 1S-4, through RNA interference with Δ12 desaturase gene expression. But ARA biosynthesis is multi-step, subject to multiple factors. The modification of one or two genes is not easy to produce transformants with the desired trait. Overexpression of VHb could increase the intensity of respiratory chain and increase the supply of intracellular ATP, which may affect the expression of intracellular multi-gene expression and promote the formation of ARA.
To the best of our knowledge, VHb has not been used in M. alpina to date. In this study, we investigate the effect of VHb on growth and ARA production in Mortierella alpina and explored the mechanism of VHb in ARA synthesis.
Methods
Strains, plasmids and growth conditions
Wild-type Mortierella alpina ATCC 32222 was obtained from the American Type Culture Collection (ATCC, Manassas, VA, USA). The Agrobacterium tumefaciens strain C58C1 and plasmid pBIG4MRHrev were gifts from Yasuyuki Kubo (Kyoto Prefectural University, Japan). The strains, plasmids and primers used in this study are listed in Additional file 1: Table S1 and Table S2, respectively. The composition of the LB-Mg agar, minimal medium (MM) and inducing medium (IM), which were used for the culture, transformation and infection of Agrobacterium tumefaciens, respectively, have been described previously [19]. GY medium (20 g L−1 glucose, 10 g L−1yeast extract and 20 g L−1agar) was used for the transformation and screening of Mortierella alpina. M. alpina strains were grown on PDA medium (potatoes 200 g L−1, glucose 20 g L−1, and agar 20 g L−1) slants at 25 °C. The seed culture medium contained 30 g L−1 glucose, 6 g L−1 yeast extract, 3 g L−1 KH2PO4, 2.8 g L−1 NaNO3, and 0.5 g L−1 MgSO4•7H2O. The medium for shake-flask culture contained 80 g L−1 glucose, 11 g L−1 yeast extract, 3.8 g L−1 KH2PO4, 3.5 g L−1 NaNO3, and 0.5 g L−1 MgSO4•7H2O. The medium for bioreactor fermentation contained 30 g L−1 glucose, 20 g L−1 yeast extract, 4 g L−1 KH2PO4, 3.8 g L−1 NaNO3, and 0.6 g L−1 MgSO4•7H2O. The Escherichia coli strains were grown in LB medium (10 g L−1 tryptone, 5 g L−1 yeast extract, and 10 g L−1 NaCl) at 37 °C.
Codon analysis and optimization
The codon preference table was found by analyzing the codon preference of the gene of M. alpina by the online software Codon Usage Database (http://www.kazusa.or.jp/codon/). The vgb gene (GenBank accession no.M30794.1) of Vitreoscilla was optimized by software Optimizer (http://genomes.urv.cat/OPTIMIZER/) based on the codon preference table. The codon-optimized nucleotide sequence of vgb was synthesized by GenScript (Nanjing) Co., Ltd. (Jiangsu, China).
Vector construction
The hygromycinB phosphotransferase gene (HPH) expression cassette containing the hisH4.1 promoter, the hpt resistance gene and the trpC terminator was amplified with primer pair P1/T1 from the vector pD4 [20]. This HPH expression cassette was inserted into pMD19-T to yield the plasmid pMD19T-HPH. The 18S sequence, which was amplified from M. alpina ATCC 32222 cDNA with primer pair 18S-F/18S-R, was ligated to the pMD19T-HPH vector, and the resulting construct was named pMD19T-HPH-18S. The carboxin resistance gene (CBXB) [21] was amplified with the primer pair CBXB-F / CBXB-R and used to replace the hpt gene, thus forming plasmid pMD19T-CBXB-18S. The CBXB expression cassette was amplified from plasmid pMD19T-CBXB-18S with primer pair P2/T2, digested with restriction endonucleases ClaI and EcoRI, and then ligated to T-DNA binary vector pBIG4MRHrev. The resulting plasmid was named pBIG-CBXB. The HisH4.1 promoter and trpC terminator were first amplified with primer sets P3-F/P3-R and T4-F/T4-R from the vector pBIG-CBXB, thus yielding fragments of 1.1 kb and 0.7 kb, respectively.
The optimized vgb sequence was inserted into pUC57 to create the plasmid pUC57-vgb (Additional file 1: Table S1). The primer pair VHb-F/VHb-R was used to amplify the optimized vgb gene from the vector pUC57-vgb, forming a 441 bp fragment. The vgb gene cassette containing the hisH4.1 promoter, the vgb gene and the trpC terminator was assembled by overlap PCR. The obtained 2.1 kb vgb gene cassette was digested with EcoRI and SmaI and then ligated to pBIG-CBXB, thus forming the plasmid pBIG-CBXB-VHb (Additional file 1: Figure S2).
Agrobacterium tumefaciens-mediated transformation of M. alpina
The transformation method was optimized on the basis of previously described Agrobacterium tumefaciens-mediated transformation (ATMT) methods [19, 22]. M. alpina spores were collected from cultures growing on PDA agar medium, and the suspension was filtered through three layers of lens paper. A. tumefaciens C58C1 was electrotransformed with the plasmid pBIG-CBXB-VHb. The positive transformants were confirmed by PCR, and then a single colony containing plasmid pBIG-CBXB-VHb was selected to grow overnight in 5 mL of liquid LB-Mg medium containing 50 μg mL−1 kanamycin and 50 μg mL−1 rifampin at 28 °C and 200 rpm. The bacterial cells were collected by centrifugation at 5000 rpm for 3 min at room temperature and washed once with fresh IM, and then fresh IM was used to adjust the OD to 0.2-0.3. After being incubated at 28 °C until an OD660 of 0.8 was reached, 100 μL of the Agrobacterium culture was mixed with 100 μL of M. alpina spore suspension (108 mL−1) and then spread onto cellophane membranes, placed on co-cultivation medium (IM with 1.5% agar) and incubated at 23 °C for 2 days. After co-cultivation, the cellophane membranes were transferred to GY plates containing 150 μg mL−1 carboxin, 50 μg mL−1 spectinomycin and 50 μg mL−1 cefotaxime to inhibit the growth of Agrobacterium. The plates were incubated at 25 °C until CBXB-resistant colonies became visible. The transformed candidates were transferred to GY agar plates containing 150 μg mL−1 carboxin. To obtain stable transformants, this procedure was repeated three times, and all experiments were conducted in triplicate.
PCR verification of the transformants
PCR was used to identify whether the mycelium was successfully transformed. Genomic DNA of M. alpina strains was prepared by a method described previously [23]. The genomic DNAs of the transformants were used as PCR templates to confirm the integration of the plasmid pBIG-CBXB-VHb by using the vgb gene specific primers VHb-F and VHb-R (Additional file 1: Table S2). Wild type M. alpina and the plasmid pBIG-CBXB-VHb were used as negative and positive PCR controls, respectively. The PCR products were run on a 1.0% agarose gel and stained with ethidium bromide.
Analytical methods for dry mycelial weight, total lipid and arachidonic acid production
Cell dry weight determination
In total, 50 mL cell suspensions were harvested by filtration at room temperature. The cell pellet was washed twice with distilled water and dried to constant weight at 50 °C. The dry cell weight was determined by weight analysis and represented by dry cell weight (DCW).
Lipid extraction and fatty acid composition analysis
Fatty acid extraction and methylation were carried out according to the previously described methods [24]. Total lipids were extracted by using approximately 100 mg of mycelia (dry weight). Fatty acid methyl esters (FAMEs) were obtained by reacting the lipids at 85 °C for 2.5 h in the presence of 2% sulfuric acid/methanol (v/v). FAMEs were extracted in hexane and analyzed by gas chromatography (Agilent Technologies, 7890B) with an HP-INNOWAX capillary column (30 m by 0.25 mm, 0.25 μm film thickness). The oven temperature was set at 100 °C for 1 min, was raised to 250 °C at a rate of 15 °C per minute and then was held at 250 °C for 5 min. The split ratio was 1:19, and the carrier gas was nitrogen. The injection volume was 1 μL. Peak detection used a flame ionization detector (FID). The temperature of the flame ionization port and injection port was 280 °C.
Shake-flask culture and bioreactor scale fermentation
To study the effect of VHb on M. alpina, especially under the oxygen-restricted conditions, we compared the DCW, total lipid content, and arachidonic acid production between the transformant VHb-20 and the wild-type strain in shake flasks and a bioreactor under dO2 limiting and non-limiting conditions. First, the VHb-20 transformants and wild type were inoculated into six 250 mL shake flasks with baffles. Three shake flasks contained 50 mL medium (normal, dO2 non-limiting condition), and three contained 150 mL medium (dO2 restriction). They were shaken at 25 °C and 200 rpm for 7 days. Meanwhile, bioreactor cultures were grown in 5-L Biostat B plus bioreactors (Sartorius Stedim Biotech, Germany) with 3 L of medium. The rate of ventilation and agitation during growth in the bioreactor was set to non-limiting oxygen (1.7 vvm, 200 rpm) and limiting oxygen (0.8 vvm, 100 rpm) conditions. The transformant and wild-type cultures were grown for 7 days at 25 °C, pH 6.5. The glucose concentration was maintained at approximately 20 g L−1 by feeding the 80% (w / v) glucose solution, which was measured with a SBA-40D Biosensor (Institute of Biology, Shandong Academy of Sciences, China). Twenty milliliter samples were taken at 24 h intervals in all experiments. The entire fermentation process used bioprocess-automated software for monitoring and control.
Quantitative real-time PCR (qRT-PCR) validation
The expression levels of some desaturase genes, such as Δ9, Δ12, Δ6, and Δ5 desaturase, were further quantified by real-time PCR to validate the effects of VHb expression on the biosynthesis of unsaturated fatty acids. The primer sequences (Additional file 1: Table S3) were designed according to the genome sequencing data (NCBI BioProjects PRJNA41211). Total RNA was extracted using a thermo Scientific GeneJET RNA Purification kit (No. K0731). After DNA degradation by DNase, 2 μg of total RNA was reverse transcribed using a TIANScript reverse transcription Kit (Tiangen) in a 20 μl reaction volume.
qRT-PCR was performed in a LightCycler R480 Real-time Detection System (Roche). A melting curve analysis of the amplification products was performed to confirm the specificity of amplification. To quantify the transcription of each gene, the copy number was determined by generating a standard curve by using a serial 10-fold dilution of the targeted PCR product inserted into the pMD™ 19-T vector (TaKaRa Bio Group). For sample normalization, 18S rRNA was used as an internal standard. All the reactions were performed in triplicate, and the data were normalized by using the average values for the internal standard.
Calculation and statistical analysis
All results are given as the mean ± standard error of the mean (SEM), and significant differences (P < 0.05, P < 0.01) between means were compared by the T-test.
Results
The stabilized expression of VHb in M. alpina
To express VHb efficiently in M. alpina, we designed and optimized the vgb gene according to the codon preference in M. alpina. The optimized vgb gene sequence is shown in Additional file 1: Figure S1.
We first tested the sensitivity of M. alpina ATCC 32222 to carboxin and another series of antibiotics (Additional file 1: Table S4). The strain did not grow in the presence of 150 μg mL−1 carboxin but showed high resistance to other antibiotics. Therefore, the carboxin resistance gene (CBXB) was used as the selection marker [21, 25] at a screening concentration of 150 μg mL−1.
The plasmid pBIG-CBXB-VHb was transformed into M. alpina ATCC 32222 by ATMT. All clones were verified by PCR using genomic DNA as templates. To select the high-yield ARA strain, 14 correct clones verified by PCR were randomly selected to check ARA production in limiting and non-limiting oxygen conditions. After 7 d of cultivation, a positive clone exhibiting the highest ARA production (VHb-20) was identified. Thus, VHb-20 strain was chosen for all further experiments. The vgb gene expression cassette inserted in clone VHb-20 was confirmed by genomic PCR amplification (see Fig. 1a and b).
Fig. 1.
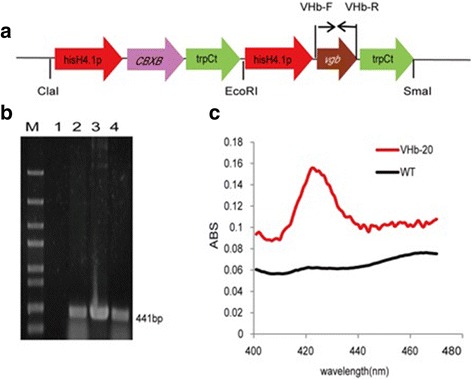
Transformant verification. a Scheme for expression of vgb in Mortierella alpina ATCC 32222; b PCR analysis of transformants of Mortierella alpina. The PCR product corresponding to VHb is 441 bp (Lanes: M, 5000 bp marker; 1, negative control, wild-type strain ATCC 32222; 2, transformant VHb-20; 3, positive control, plasmid pBIG-CBXB-VHb; 4, transformant VHb-10); c CO binding spectra of cell extracts from original strain ATCC 32222 and transformant VHb-20
To confirm the functional expression of VHb in VHb-20, the carbon monoxide (CO) difference spectra were measured, and the wild-type strain was used as a control. VHb-20 showed a typical VHb CO-binding absorption peak at 419 nm, whereas this absorption peak was not observed in the control (Fig. 1c). This result indicated that the VHb expressed in VHb-20 was biologically active. Afterwards, VHb-20 was continuously inoculated and cultivated on carboxin-free PDA solid medium, and the integrated vgb gene in the genome was monitored by PCR. The result indicated that the recombinant strain was genetically stable after 5 sub-cultivation cycles.
Effects of VHb expression on the growth and ARA production of M. alpina
Both VHb-20 and the wild-type strain were cultivated in 250 mL shake flasks under limiting and non-limiting oxygen conditions, and their growth patterns and ARA production were investigated. The results showed that the VHb-20 strain grew faster than the wild-type strain and formed more homogeneous mycelial pellets under both conditions. In addition, the hyphae of M. alpina ATCC 32222 were looser.
Under aerobic conditions (cultivated in a 250-mL baffled flask containing 50 mL of medium at 25 °C and 200 rpm), the glucose consumption rates and dry weight of VHb-20 and wild-type strains were not obviously different after cultivation for 7 days (Fig. 2a and b), but the total lipid yield of strain VHb-20 (7.97 g L−1) was 3.15-fold higher than that of the wild-type strain (2.53 g L−1; Fig. 2c). In addition, the ARA production of the strain VHb-20 was 3.62 g L−1, which was 5.10 times higher than that of the wild-type strain (0.71 g L−1; Fig. 2d). To obtain a microaerobic condition with a lower level of dissolved oxygen, the liquid medium in the flask was increased from 50 mL to 150 mL. Under these conditions, the VHb-20 strain had a higher glucose consumption rate and growth rate compared with that of the control strain (Fig. 2a and b), and lipid and ARA production of VHb-20 was also higher by 4.03 folds (4.64 g L −1 vs. 1.15 g L−1) and 8.36 folds (1.84 g L −1 vs. 0.22 g L −1), respectively (Fig. 2c, and d).
Fig. 2.
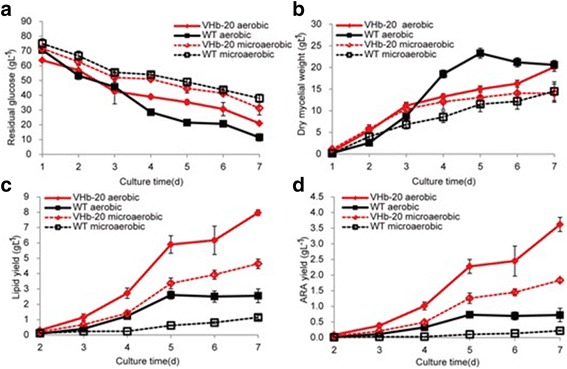
Comparison of growth profiles in shake flasks under aerobic and microaerobic conditions. a Residual glucose; b Dry mycelial weight; c Total lipid content; (d) Arachidonic acid production; The data are the mean of three repeats
Effect of VHb expression on the composition of fatty acids in M. alpina
The concentration of dissolved oxygen affects not only cell growth but also the synthesis of fatty acids. Therefore, we further compared the fatty acid composition and content of VHb-20 and the wild-type strain under aerobic or microaerobic conditions. VHb-20 and the wild-type strain produced similar types of fatty acids (Fig. 3), but the proportion of various fatty acids in the lipids had changed significantly (Table 1).The results showed that the proportion of palmitic acid (C16: 0), stearic acid (C18: 0) and oleic acid (C18: 1) in the total fatty acids of wild type strains was 15.82%, 17.98% and 17.58%, significantly higher than other fatty acids. Especially in the limited oxygen condition, the accumulation of oleic acid (C18: 1) is very huge, the content in the total fatty acids as high as 39.61%, resulting in ARA content was only 19.27%. However, the contents of palmitic acid (C16: 0) and oleic acid (C18: 1) in VHb-20 strain were significantly decreased, and the ARA content was increased to 45.68% and 38.51%, under the aerobic or microaerobic conditions respectively, which were nearly 1.7 folds and 2.0 folds as high as that of the control.
Fig. 3.
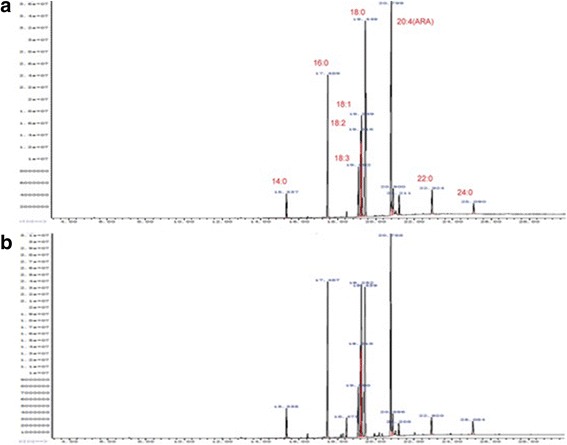
GC-MS analysis of fatty acids composition from wild type and VHb-20. a VHb-20 in shake flasks with free air diffusion; b Wild type in shake flasks with free air diffusion
Table 1.
Fatty Acid Compositions of wild type and VHb-20 in shake flasks
| Fatty acid | Content (% total fatty acids) | |||
|---|---|---|---|---|
| WT(aerobic) | VHb-20(aerobic) | WT(microaerobic) | VHb-20(microaerobic) | |
| 14:0 | 3.22 ± 0.14 | 0.73 ± 0.02** | 2.84 ± 0.04 | 1.46 ± 0.17** |
| 16:0 | 15.82 ± 0.33 | 9.72 ± 0.36** | 13.35 ± 0.10 | 11.51 ± 0.24** |
| 18:0 | 17.98 ± 0.49 | 19.63 ± 0.73** | 13.34 ± 0.95 | 23.19 ± 0.48** |
| 18:1 | 17.58 ± 0.58 | 6.92 ± 0.55** | 39.61 ± 2.26 | 8.34 ± 0.24** |
| 18:2 | 7.43 ± 0.44 | 5.82 ± 0.22** | 2.87 ± 0.35 | 5.94 ± 0.56** |
| 18:3 | 3.37 ± 0.11 | 3.74 ± 0.08* | 2.27 ± 0.38 | 3.71 ± 0.22* |
| 20:3 | 3.43 ± 0.06 | 2.84 ± 0.19* | 3.71 ± 0.08 | 2.38 ± 0.26** |
| 20:4(ARA) | 27.30 ± 0.77 | 45.68 ± 1.09** | 19.27 ± 1.18 | 38.51 ± 0.44** |
**p < 0.01,*p < 0.05
Effects of VHb expression on the expression of desaturase genes
The biosynthesis of unsaturated fatty acids in cell is catalyzed by desaturases, which require oxygen as a substrate [3]. Therefore, the expression of desaturase genes, including Δ9 desaturase, Δ12 desaturase, Δ6 desaturase, Δ5 desaturase, was investigated to verify the effects of VHb expression on the biosynthesis of unsaturated fatty acids at the molecular level. All the expression levels of these desaturase genes were higher in VHb-20, as compared with the wild type strain, in both aerobic and microaerobic conditions (Fig. 4a). From the results, Δ9 desaturase, which is the first enzyme in unsaturated fatty acid biosynthesis, was most sensitive to the expression of VHb, which resulted 3.2 and 4.2-fold higher expression of the Δ9 desaturase gene under aerobic and microaerobic conditions, respectively. A lesser effect was observed in the increased expression of Δ12 and Δ6 desaturase, which were only 1.4-fold higher in VHb-20, and 1.6-fold higher expression of Δ5 desaturase was observed under aerobic conditions. However, under microaerobic conditions, the expression level of Δ12 and Δ6 desaturase was increased by 2.3 and 8.7 times higher than that of the control strain. In agreement with the differences in the levels of these desaturases, the biosynthesis of unsaturated fatty acids was augmented, and the ARA production of VHb-20 was increased to 3.62 g L−1 and 1.84 g L −1 under the aerobic and microaerobic conditions (Fig. 4b), respectively.
Fig. 4.
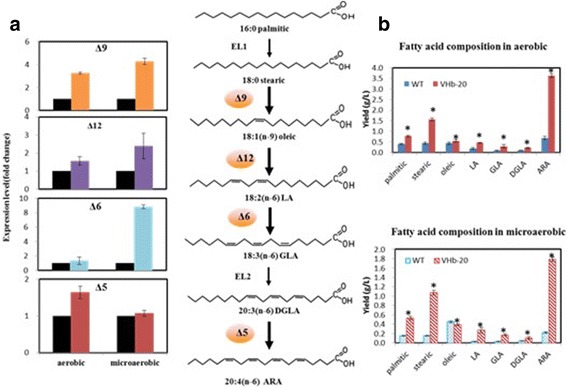
The changes of the expression of desaturases (Δ9 desaturase, Δ12 desaturase, Δ6 desaturase, Δ5 desaturase) and ARA production between the WT and VHb-20 strains under aerobic and microaerobic conditions. a Changes in the expression of desaturases, Black: WT; Colored: VHb-20 strain; b Changes in unsaturated fatty acid production (*p < 0.05)
Confirmation of the effects of VHb expression on cell growth and ARA production by using bioreactor fermentation
Bioreactor cultures were grown to confirm the influence of VHb expression on cell growth and lipid production under both non-limiting (1.7vvm, 200 rpm) and limiting (0.8vvm, 100 rpm) dO2 conditions. From a morphological viewpoint (Fig. 5), VHb-20 grew faster and formed more uniform mycelial pellets than did the wild-type strain M. alpina ATCC 32222. In hypoxic conditions, the VHb-20 strain grew faster than the wild-type strain and reached the maximum biomass (12.90% more than the wild type) sooner (Fig. 6a). In addition, the culture of the wild-type strain clumped heavily and became difficult to stir after 3 days of cultivation, whereas uniform mycelial pellets were observed in the VHb-20 culture, and the culture broth maintained good fluid properties through 7 days of fermentation.
Fig. 5.
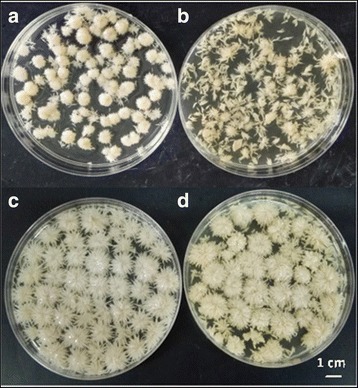
Morphology of hyphae in 5 L fermenter. a VHb-20 transformant under aerobic conditions; b wild type under vaerobic conditions; c VHb-20 transformant under microaerobic conditions; d wild type under microaerobic conditions
Fig. 6.
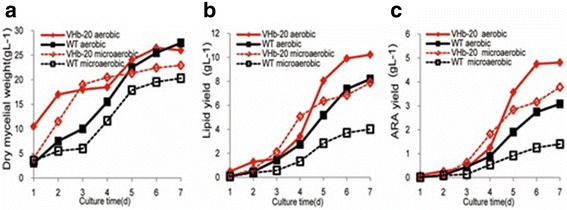
Comparison of growth profiles in a 5-L fermenter under aerobic and microaerobic conditions. a Dry mycelial weight; b Total lipid content; c Arachidonic acid production
In agreement with the results of the shake flask assay, VHb expression also enhanced lipid and ARA biosynthesis in the bioreactor experiments, especially under limiting oxygen conditions. There was almost no difference in the lipid content of the DCW of VHb-20 in the two conditions (39.30% vs 34.40% for the normal and limiting oxygen conditions, respectively), but in the control, the lipid content was 34% lower in limiting oxygen conditions (29.80% vs 19.80%) at the end of fermentation. The maximum lipid yield of VHb-20 strain was 10.21 g L−1 and 7.89 g L −1, which was nearly 1.25 and 1.96 fold higher than that of the original strain under the two aerobic conditions (Fig. 6b).
Under aerobic conditions, VHb-20 had a higher ARA yield by 1.56-fold, from a value of 3.08 g L−1 to 4.81 g L−1, compared with the original strain M. alpina ATCC 32222. At the end of fermentation, the ARA yield of VHb-20 was 2.70 times (3.78 g L−1 vs 1.40 g L−1) higher than that of the wild type under microaerobic conditions (Fig. 6c).
Discussion
ARA is an essential polyunsaturated fatty acid in human nutrition that has broad applications in many industries [26]. Although M. alpina is considered to be a good production strain for high level of ARA production, the low percentage of ARA in total lipids results in a production bottleneck [27]. In this study, we demonstrated that expression of Vitreoscilla hemoglobin significantly improves ARA and total lipid production in M. alpina.
Heterologous expression of VHb in M. alpina increased cell growth as well as oxygen uptake under non-limiting and limiting dO2 conditions. Furthermore, VHb expression shortened the lag phase for the growth of M. alpina and the appearance of ARA. A similar observation has previously been reported in S. occidentalis, in which alpha-amylase appears earlier in the culture medium of VHb-expressing cells [28], and the result is also consistent with the expression of VHb in Gluconobacter oxydans [29] and Phellinus igniarius [8]. Importantly, the morphology of the VHb-20 strain changed significantly. The mutant VHb-20 stain formed small homogeneous globules, whereas the wild-type strain formed loose mycelia (Fig. 5). Previous experiments have shown that in late fermentation stages, the wild-type strain readily agglomerates, thus resulting in decreased dissolved oxygen. In contrast, mycelial pellets increased the fermentation fluidity, which is more conducive to dissolved oxygen and accumulation of lipids and ARA [7]. Moreover, the growth pattern of M. alpina has a large effect on ARA production [30]. These results are also consistent with findings by Higashiyama et al. [31] indicating that high concentrations of dissolved oxygen are necessary for obtaining smooth pellets, and the number of spherical fluffy pellets and the dissolved oxygen concentration are positively correlated.
Dissolved oxygen levels affect not only cell growth but also the synthesis of fatty acids in M. alpina. Therefore, the VHb-20 and wild-type strains were analyzed for fatty acid and ARA content under normal culture conditions and dissolved oxygen-limiting conditions. Studies have shown that the expression of VHb increases the yield of ARA and total lipids, especially under oxygen-limiting conditions. It has been reported previously that VHb enhances ATP production and respiratory activity [32]. Evidence has shown that VHb improves the efficiency of oxidative phosphorylation of cells by regulating the activity of the relevant respiratory oxidase, thereby changing the central carbon metabolic pathway, and ultimately promoting cell growth and protein expression under hypoxic conditions [33, 34]. In addition, VHb also has peroxidase activity, and many heterologous hosts protect against oxidative invasion by expressing this gene, in agreement with the hypothesis that VHb is a carrier of oxygen in the cell [12, 35]. On the basis of these observations, VHb expression may promote fatty acid synthesis and increase total lipid accumulation, owing to the enhancement of respiratory activity and ATP supply.
In addition, another prominent finding was that VHb expression significantly changed the fatty acid composition, particularly by decreasing the ratio of C16:0 and C18:1 and increasing the ratio of C20:4 (ARA; Table 1). The biosynthesis of ARA strongly depends on oxygen accessibility, and some specific catalytic functions of various fatty acid desaturases and elongation enzymes involved in ARA synthesis in M. alpina have been elucidated [36, 37]. The oxygen requirement for desaturation was higher than that for cell growth and total lipid production. Therefore, the increase in ARA content in VHb-20 may be a result of the overall increase of the fatty acid desaturase caused by the presence of heterologous hemoglobin. This speculation is consistent with the expression level of desaturases that we determined by qRT-PCR. It can be seen from Fig. 4 that four kinds of desaturases in the ARA metabolic pathway are Δ9-, Δ12-, Δ6-, Δ5- desaturases respectively, and their expression levels are significantly up-regulated under both the aerobic and microaerobic conditions. Moreover, these desaturases were the key enzymes catalyzed the palmitic acid (C16: 0), stearic acid (C18: 0) and Oleic acid (C18: 1) converted to ARA. Therefore, we hypothesized that the increase of ARA production of M. alpina is probably due to the increase of the expression of desaturase and the improved desaturase activities in ARA metabolic pathway and the promotion of metabolic flow to ARA synthesis. More detailed mechanisms need further study.
Conclusions
In conclusion, the successful expression of VHb in M. alpina significantly increases ARA and lipid production as well as promotes cell growth and increase viability under both lower and normal dissolved oxygen conditions. Therefore, we suggest that expression of the vgb gene in M. alpina might have good prospects in high-density fermentation. However, the specific mechanisms underlying the effects of VHb in M. alpina are not clear and require further investigation.
Acknowledgements
The authors are thankful to Prof. Yasuyuki Kubo for providing the Agrobacterium tumefaciens strain C58C1 and plasmid pBIG4MRHrev. This study is a contribution to the international IMBER project.
Funding
This work is financially supported by the National Key Research and Development Program (no. 2016YFA0601400), the National Natural Science Foundation of China (no. 41306132), and the National High Technology Research and Development Program of China (no. 2014AA021701). This study is a contribution to the international IMBER project.
Availability of data and materials
The strains, plasmids, primers, optimized vgb Sequence and Schematic representation of the plasmid construction are supplied as supporting information.
Abbreviations
- ARA
Arachidonic acid
- ATMT
Agrobacterium tumefaciens-mediated transformation
- CO
Carbon monoxide
- DCW
Dry cell weight
- DGLA
Dihomo-γ-linolenic acid
- DO
Dissolved oxygen
- FAMEs
Fatty acid methyl esters
- FID
Flame ionization detector
- GLA
γ-linolenic acid
- HPH
hygromycinb phosphotransferase gene
- IM
Inducing medium
- LA
Linoleic acid
- M. alpina
Mortierella alpina
- MM
Minimal medium
- PUFAs
Polyunsaturated fatty acids
- VHb
Vitreoscilla hemoglobin
Additional file
(DOCX 263 kb)
Authors’ contributions
Huidan Zhang performed the experiments as part of her doctoral work. All this work was carried out under the supervision of Qiu Cui and Xiaojin Song. Xiaojin Song assisted in the bioreactor experiment. Qiu Cui and Yingang Feng also assisted in editing the manuscript. All the authors have read and approved the final manuscript.
Competing interest
The authors declare that they have no competing interests.
Ethics approval and consent to participate
Not applicable.
Consent for publication
Not applicable.
Publisher’s Note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Footnotes
Electronic supplementary material
The online version of this article (doi:10.1186/s12896-017-0388-8) contains supplementary material, which is available to authorized users.
Contributor Information
Huidan Zhang, Email: zhanghd@qibebt.ac.cn.
Yingang Feng, Email: fengyg@qibebt.ac.cn.
Qiu Cui, Phone: +86 532 80662706, Email: cuiqiu@qibebt.ac.cn.
Xiaojin Song, Phone: +86 532 80662705, Email: songxj@qibebt.ac.cn.
References
- 1.You JY, Peng C, Liu X, Ji XJ, Lu J, Tong Q, et al. Enzymatic hydrolysis and extraction of arachidonic acid rich lipids from Mortierella alpina. Bioresour Technol. 2011;102:6088–6094. doi: 10.1016/j.biortech.2011.01.074. [DOI] [PubMed] [Google Scholar]
- 2.Wang HC, Yang B, Hao GF, Feng Y, Chen HQ, Feng L, et al. Biochemical characterization of the tetrahydrobiopterin synthesis pathway in the oleaginous fungus Mortierella alpina. Microbiology. 2011;157:3059–3070. doi: 10.1099/mic.0.051847-0. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 3.Dedyukhina EG, Chistyakova TI, Vainshtein MB. Biosynthesis of arachidonic acid by micromycetes (review) Appl Biochem Microbiol. 2011;47:109–117. doi: 10.1134/S0003683811020037. [DOI] [PubMed] [Google Scholar]
- 4.Nie ZK, Ji XJ, Shang JS, Zhang AH, Ren LJ, Huang H. Arachidonic acid-rich oil production by Mortierella alpina with different gas distributors. Bioprocess Biosyst Eng. 2014;37:1127–1132. doi: 10.1007/s00449-013-1104-2. [DOI] [PubMed] [Google Scholar]
- 5.Wu WJ, Yan JC, Ji XJ, Zhang X, Shang JS, Sun LN, et al. Lipid characterization of an arachidonic acid-rich oil producing fungus Mortierella alpina. Chin J Chem Eng. 2015;23:1183–1187. doi: 10.1016/j.cjche.2015.04.015. [DOI] [Google Scholar]
- 6.Dedyukhina EG, Chistyakova TI, Mironov AA, Kamzolova SV, Minkevich IG, Vainshtein MB. The effect of pH, aeration, and temperature on arachidonic acid synthesis by Mortierella alpina. Appl Biochem Microbiol. 2015;51:242–248. doi: 10.1134/S0003683815020040. [DOI] [PubMed] [Google Scholar]
- 7.Higashiyama K, Fujikawa S, Park EY, Okabe M. Image analysis of morphological change during arachidonic acid production by Mortierella alpina 1S-4. J Biosci Bioeng. 1999;87:489–494. doi: 10.1016/S1389-1723(99)80098-X. [DOI] [PubMed] [Google Scholar]
- 8.Zhu H, Sun S, Zhang S. Enhanced production of total flavones and exopolysaccharides via Vitreoscilla hemoglobin biosynthesis in Phellinus igniarius. Bioresour Technol. 2011;102:1747–1751. doi: 10.1016/j.biortech.2010.08.085. [DOI] [PubMed] [Google Scholar]
- 9.Li M, Wu J, Lin J, Wei D. Expression of Vitreoscilla hemoglobin enhances cell growth and Dihydroxyacetone production in Gluconobacter oxydans. Curr Microbiol. 2010;61:370–375. doi: 10.1007/s00284-010-9621-6. [DOI] [PubMed] [Google Scholar]
- 10.Suen YL, Tang H, Huang J, Chen F. Enhanced production of fatty acids and astaxanthin in Aurantiochytrium sp. by the expression of Vitreoscilla hemoglobin. J Agric Food Chem. 2014;62:12392–12398. doi: 10.1021/jf5048578. [DOI] [PubMed] [Google Scholar]
- 11.Tao L, Sedkova N, Yao H, Ye RW, Sharpe PL, Cheng Q. Expression of bacterial hemoglobin genes to improve astaxanthin production in a methanotrophic bacterium Methylomonas sp. Appl Microbiol Biotechnol. 2006;74:625–633. doi: 10.1007/s00253-006-0708-8. [DOI] [PubMed] [Google Scholar]
- 12.Stark BC, Pagilla KR, Dikshit KL. Recent applications of Vitreoscilla hemoglobin technology in bioproduct synthesis and bioremediation. Appl Microbiol Biotechnol. 2015;99:1627–1636. doi: 10.1007/s00253-014-6350-y. [DOI] [PubMed] [Google Scholar]
- 13.Frey AD, Kallio PT. Bacterial hemoglobins and flavohemoglobins: versatile proteins and their impact on microbiology and biotechnology. FEMS Microbiol Rev. 2003;27:525–545. doi: 10.1016/S0168-6445(03)00056-1. [DOI] [PubMed] [Google Scholar]
- 14.Zhang S, Wang J, Wei Y, Tang Q, Ali MK, He J. Heterologous expression of VHb can improve the yield and quality of biocontrol fungus Paecilomyces lilacinus, during submerged fermentation. J Biotechnol. 2014;187:147–153. doi: 10.1016/j.jbiotec.2014.07.438. [DOI] [PubMed] [Google Scholar]
- 15.Bhave SL, Chattoo BB. Expression ofvitreoscilla hemoglobin improves growth and levels of extracellular enzyme in Yarrowia lipolytica. Biotechnol Bioeng. 2003;84:658–666. doi: 10.1002/bit.10817. [DOI] [PubMed] [Google Scholar]
- 16.Wang X, Sun Y, Shen X, Ke F, Zhao H, Liu Y, et al. Intracellular expression of Vitreoscilla hemoglobin improves production of Yarrowia lipolytica lipase LIP2 in a recombinant Pichia pastoris. Enzym Microb Technol. 2012;50:22–28. doi: 10.1016/j.enzmictec.2011.09.003. [DOI] [PubMed] [Google Scholar]
- 17.Hao G, Chen H, Gu Z, Zhang H, Chen W, Chen YQ, Cullen D. Metabolic engineering of Mortierella alpina for enhanced Arachidonic acid production through the NADPH-supplying strategy. Appl Environ Microbiol. 2016;82(11):3280–3288. doi: 10.1128/AEM.00572-16. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 18.Takeno S, Sakuradani E, Tomi A, Inohara-Ochiai M, Kawashima H, Ashikari T, Shimizu S. Improvement of the fatty acid composition of an oil-producing filamentous fungus, Mortierella alpina 1S-4, through RNA interference with delta12-desaturase gene expression. Appl Environ Microbiol. 2005;71(9):5124–5128. doi: 10.1128/AEM.71.9.5124-5128.2005. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 19.Ando A, Sumida Y, Negoro H, Suroto DA, Ogawa J, Sakuradani E, et al. Establishment of Agrobacterium tumefaciens-mediated transformation of an oleaginous fungus, Mortierella alpina 1S-4, and its application for eicosapentaenoic acid producer breeding. Appl Environ Microbiol. 2009;75:5529–5535. doi: 10.1128/AEM.00648-09. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 20.Mackenzie D, Wongwathanarat P, Carte A, Archer D. Isolation and use of a homologous Histone H4 promoter and a ribosomal DNA region in a transformation vector for the oil-producing fungus Mortierella alpina. Appl Environ Microbiol. 2000;66:4655–4661. doi: 10.1128/AEM.66.11.4655-4661.2000. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 21.Ando A, Sakuradani E, Horinaka K, Ogawa J, Shimizu S. Transformation of an oleaginous zygomycete Mortierella alpina 1S-4 with the carboxin resistance gene conferred by mutation of the iron-sulfur subunit of succinate dehydrogenase. Curr Genet. 2009;55:349–356. doi: 10.1007/s00294-009-0250-1. [DOI] [PubMed] [Google Scholar]
- 22.Hao G, Chen H, Wang L, Gu Z, Song Y, Zhang H, et al. Role of malic enzyme during fatty acid synthesis in the oleaginous fungus Mortierella alpina. Appl Environ Microbiol. 2014;80:2672–2678. doi: 10.1128/AEM.00140-14. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 23.Takeno S, Sakuradani E, Tomi A, Inohara-Ochiai M, Kawashima H, Shimizu S. Transformation of oil-producing fungus, Mortierella alpina 1S-4, using Zeocin, and application to arachidonic acid production. J Biosci Bioeng. 2005;100:617–622. doi: 10.1263/jbb.100.617. [DOI] [PubMed] [Google Scholar]
- 24.Cheng YR, Sun ZJ, Cui GZ, Song XJ, Cui Q. A new strategy for strain improvement of Aurantiochytrium sp. based on heavy-ions mutagenesis and synergistic effects of cold stress and inhibitors of enoyl-ACP reductase. Enzym Microb Technol. 2016;93-94:182–190. doi: 10.1016/j.enzmictec.2016.08.019. [DOI] [PubMed] [Google Scholar]
- 25.Kikukawa H, Sakuradani E, Ando A, Okuda T, Ochiai M, Shimizu S, et al. Disruption of lig4 improves gene targeting efficiency in the oleaginous fungus Mortierella alpina 1S-4. J Biotechnol. 2015;208:63–69. doi: 10.1016/j.jbiotec.2015.05.020. [DOI] [PubMed] [Google Scholar]
- 26.Ji XJ, Zhang AH, Nie ZK, Wu WJ, Ren LJ, Huang H. Efficient arachidonic acid-rich oil production by Mortierella alpina through a repeated fed-batch fermentation strategy. Bioresour Technol. 2014;170:356–360. doi: 10.1016/j.biortech.2014.07.098. [DOI] [PubMed] [Google Scholar]
- 27.Li X, Lin Y, Chang M, Jin Q, Wang X. Efficient production of arachidonic acid by Mortierella alpina through integrating fed-batch culture with a two-stage pH control strategy. Bioresour Technol. 2015;181:275–282. doi: 10.1016/j.biortech.2015.01.009. [DOI] [PubMed] [Google Scholar]
- 28.Suthar DH, Chattoo BB. Expression of Vitreoscilla hemoglobin enhances growth and levels of alpha-amylase in Schwanniomyces occidentalis. Appl Microbiol Biotechnol. 2006;72:94–102. doi: 10.1007/s00253-005-0237-x. [DOI] [PubMed] [Google Scholar]
- 29.Li MH, Wu JA, Lin JP, Wei DZ. Expression of Vitreoscilla hemoglobin enhances cell growth and Dihydroxyacetone production in Gluconobacter oxydans. Curr Microbiol. 2010;61:370–375. doi: 10.1007/s00284-010-9621-6. [DOI] [PubMed] [Google Scholar]
- 30.Koike Y, Cai HJ, Higashiyama K, Fujikawa S, Park EY. Effect of consumed carbon to nitrogen ratio of mycelial morphology and arachidonic acid production in cultures of Mortierella alpina. J Biosci Bioeng. 2001;91:382–389. doi: 10.1016/S1389-1723(01)80156-0. [DOI] [PubMed] [Google Scholar]
- 31.Higashiyama KM, K Tsujimura H. Effects of dissolved oxygen on the morphology of an Arachidonic acid production by Mortierella alpina 1S-4. Biotechnol Bioeng. 1998;63:442–448. doi: 10.1002/(SICI)1097-0290(19990520)63:4<442::AID-BIT7>3.0.CO;2-9. [DOI] [PubMed] [Google Scholar]
- 32.Kallio PT, Kim DJ, Tsai PS, Bailey JE. Intracellular expression of Vitreoscilla hemoglobin alters Escherichia-Coli energy-metabolism under oxygen-limited conditions. Eur J Biochem. 1994;219:201–208. doi: 10.1111/j.1432-1033.1994.tb19931.x. [DOI] [PubMed] [Google Scholar]
- 33.Ramandeep HKW, Raje M, Kim KJ, Stark BC, Dikshit KL, et al. Vitreoscilla hemoglobin - intracellular localization and binding to membranes. J Biol Chem. 2001;276:24781–24789. doi: 10.1074/jbc.M009808200. [DOI] [PubMed] [Google Scholar]
- 34.Zhang L, Li Y, Wang Z, Xia Y, Chen W, Tang K. Recent developments and future prospects of Vitreoscilla hemoglobin application in metabolic engineering. Biotechnol Adv. 2007;25:123–136. doi: 10.1016/j.biotechadv.2006.11.001. [DOI] [PubMed] [Google Scholar]
- 35.Li W, Zhang Y, Xu H, Wu L, Cao Y, Zhao H, et al. pH-induced quaternary assembly of Vitreoscilla hemoglobin: the monomer exhibits better peroxidase activity. Biochim Biophys Acta (BBA) - Proteins and Proteomics. 2013;1834:2124–2132. doi: 10.1016/j.bbapap.2013.07.005. [DOI] [PubMed] [Google Scholar]
- 36.Sakuradani E. Advances in the production of various polyunsaturated fatty acids through oleaginous fungus Mortierella alpina breeding. Biosci Biotechnol Biochem. 2010;74:908–917. doi: 10.1271/bbb.100001. [DOI] [PubMed] [Google Scholar]
- 37.Vongsangnak W, Ruenwai R, Tang X, Hu X, Zhang H, Shen B, et al. Genome-scale analysis of the metabolic networks of oleaginous Zygomycete fungi. Gene. 2013;521:180–190. doi: 10.1016/j.gene.2013.03.012. [DOI] [PubMed] [Google Scholar]
Associated Data
This section collects any data citations, data availability statements, or supplementary materials included in this article.
Data Availability Statement
The strains, plasmids, primers, optimized vgb Sequence and Schematic representation of the plasmid construction are supplied as supporting information.