
Fig. 1. In vivo and in organello analysis of the CL dependence of the Tim23-Tim50 interaction.
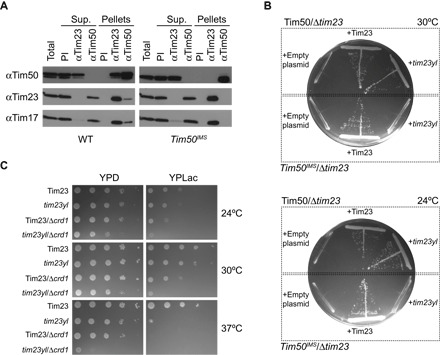
(A) Coimmunoprecipitation analysis of the Tim23-Tim50 interaction. Digitonin lysates of mitochondria from yeast strains containing full-length Tim50 (left) or Tim50IMS (right) were immunoprecipitated with affinity-purified antibodies to Tim23 (αTim23) or Tim50 (αTim50) or preimmune immunoglobulins (PI). Unbound (Sup.) and bound (Pellets) fractions were analyzed by SDS-PAGE and immunodecoration with antibodies against Tim50, Tim23, or Tim17, as indicated. Total and Sup. lanes indicate 20% of starting material; all other lanes represent undiluted samples. (B) Synthetic lethality of tim23yl and Tim50IMS. Yeast cells containing a chromosomal deletion of TIM23 complemented with a TIM23-containing URA3 plasmid were generated in a Tim50 or Tim50IMS background (delineated by dotted boxes). Cells were transformed with centromeric plasmids carrying either Tim23 or tim23yl, as indicated, and subjected to URA3 plasmid loss by plating on glucose-containing medium with 5-fluoroorotic acid (5-FOA). Cells were incubated at 30°C (upper plate) or 24°C (lower plate). (C) Genetic interaction of Δcrd1 and tim23yl. Tenfold serial dilutions were prepared from yeast cells containing WT Tim23 or the tim23yl mutant in a background of normal CL synthesis or CL synthase knockouts (Δcrd1). Cells were spotted onto YPD or YPLac medium as shown and grown at the indicated temperatures.