


Abstract
The so-called upstream binding factor (UBF) is required for the initial step in formation of an RNA polymerase I initiation complex. This function of UBF correlates with its ability to induce the ribosomal enhancesome, a structure which resembles in its mass and DNA content the nucleosome of chromatin. DNA looping in the enhancesome is probably the result of six in-phase bends induced by the HMG boxes of a UBF dimer. Here we show that insertion/deletion mutations in the basic peptide linker lying between the N-terminal dimerisation domain and the first HMG box of Xenopus UBF prevent the DNA looping characteristic of the enhancesome. Using these mutants we demonstrate that (i) the enhancesome structure does not depend on tethering of the entering and exiting DNA duplexes, (ii) UBF monomers induce hemi-enhancesomes, bending the DNA by 175 ± 24° and (iii) two hemi-enhancesomes are precisely phased by UBF dimerisation. We use this and previous data to refine the existing enhancesome model and show that HMG boxes 1 and 2 of UBF lie head-to-head along the DNA.
INTRODUCTION
The ribosomal enhancesome is a highly unusual and surprisingly complex nucleoprotein structure which underlies the function of both the RNA polymerase I (RPI) promoter and the repetitive enhancers of the ribosomal intergenic spacer (IGS) of higher eukaryotes (1–3; for reviews see 4,5). The enhancesome consists of a dimer of the upstream binding factor (UBF) and ∼140 bp of DNA, which is folded into a 360° loop (Fig. 1). UBF binding is the first step in RPI promoter recognition and formation of the pre-initiation complex and is known to promote recruitment of the TBPI complex SL1. This recruitment appears to occur via two distinct mechanisms. Firstly, SL1 specifically recognises and binds the phosphorylated C-terminal acidic tail of UBF (6–9). The second mechanism probably depends on the specific DNA architecture induced by the enhancesome, since the minimal or core region of UBF required for RPI promotion corresponds exactly with that required for the enhancesome (Fig. 1; 10,11). On the basis of these data it has been proposed that the formation of tandem enhancesomes on the UCE and Core promoter elements juxtaposes these elements, allowing their cooperative binding by SL1 (2,3,5).
Figure 1.

(A) Probable DNA contacts made by the first three HMG boxes of Xenopus UBF. A dimer of Xenopus UBF is shown with each HMG box interacting with adjacent 20 bp segments of DNA. (B) A model explaining the role of in-phase DNA bending by the HMG boxes of UBF in formation of the enhancesome. A view face on to the DNA loop and another edge on to the DNA loop are shown. In both (A) and (B) the DNA is indicated in red as a (bent) rod and the ‘core’ region of Xenopus UBF is shown in blue.
The enhancesome is also a regulatory element in RPI transcription. The first two HMG boxes of UBF bind the histone acetyltransferase (HAT) CBP or the retinoblastoma protein Rb in an exclusive or competitive fashion (12). CBP activates transcription by acetylating UBF and possibly the neighbouring chromatin while Rb represses transcription by displacing CBP from UBF and thus enhancing deacetylation (12). It has further been shown that transcription activation by UBF depends upon phosphorylation of its HMG box 5 (of mammalian UBF, equivalent to box 4 of Xenopus UBF) by cyclin-dependent kinase (CDK)-2 and/or CDK-4 (13). An analogous phosphorylation of HMG boxes 1 and 2 by the MAP kinases ERK-1 and/or ERK-2 regulates ribosomal transcription in response to growth factor signalling (Stefanovsky et al., manuscript in preparation).
A low resolution structural model of the enhancesome was deduced by electron spectroscopic imaging (ESI) and DNA footprinting (Fig. 1B; 1–3). The data suggested a complex in which six HMG boxes of a UBF dimer bend 142 ± 30 bp of DNA into a 360°, 17 nm diameter loop. In the model, DNA looping is the result of in-phase bends induced by each of the six HMG boxes. HMG box structures determined to date have shown the boxes to lie in a widened minor groove of the DNA and to induce abrupt bending or kinking towards the major groove. If, as we have proposed, this were also the case for the boxes of UBF, the DNA trajectory within the enhancesome could depend for its stability solely on in-phase DNA bending. As seen in Figure 1B, no protein core exists within the proposed ‘core’ enhancesome structure on which to tether the DNA.
Despite the strong evidence supporting our present model of the enhancesome, an alternative explanation for DNA looping within UBF–DNA stuctures cannot be completely excluded (14). In this alternative explanation UBF stabilises a looped DNA structure by crosslinking the entering and exiting DNA duplex. It is well documented that HMG boxes bind cruciform structures with high affinity (see for example 15) and such DNA structures do resemble DNA cross-over junctions (16). Given the low resolution of the images we have so far obtained by ESI it has not been possible to resolve this problem definitively.
We have now investigated a potential inter-domain linker element found between the N-terminal dimerisation domain and HMG box 1 of all UBFs and show that this element is important for the diad symmetry of the enhancesome. The data provide strong evidence for an enhancesome in which the DNA is neither tethered to a protein core nor is it tethered by crosslinking of the entering and exiting duplexes. The data also provide a demonstration of the crucial importance the inter-domain linker peptides play in the enhancesome structure and suggest that extreme care is needed in the interpretation of UBF domain swapping experiments.
MATERIALS AND METHODS
Isolation and expression of Xenopus laevis UBF mutants
Mutants were assembled in the vector pGEX-2T, expressed in Escherichia coli and purified as previously described (1,17). Nbox13 (or Core UBF) was produced by fusing amino acids 16–383 of Xenopus UBF2b (18) to GST, which was not cleaved from the final product. Mutant 598 bp NcoI–SphI fragments containing the insertion/deletion region were produced by ligation of two PCR fragments. One of them (NcoI/blunt) was produced using an upstream Xenopus UBF2b primer and the primer 5′-GCCTTTGTATGGATGTC-3′. The second fragment (blunt/SphI), which carried the ‘Plus’ or ‘Minus’ GKK mutations, was obtained using a downstream sequence primer and either primer 5′-AAAAAAGGCAAAAAAGCTTAAGAAAC-3′ (GKK insertion) or primer 5′-AAGAAACACCCCGAGTTTC-3′ (LKK deletion). The mutant NcoI–SphI fragments were formed by flush end ligating the PCR products before introducing them into the pGEX-Xenopus UBF2b vector in place of the wild-type NcoI–SphI fragment.
Dimerisation of free Core UBF protein
Wild-type, Plus and Minus Nbox13/Core UBF coding regions were subcloned (without the GST domain) into pCDNA3.1His C (Invitrogen). The corresponding [35S]methionine-labelled proteins were then produced in vitro using TnT coupled T7 reticulocyte lysate (Promega), affinity purified on M2 anti-FLAG affinity matrix (Sigma) and eluted with 0.5 mg/ml FLAG peptide following the manufacturer’s protocols.
The GST-fused wild-type and mutant Nbox13/Core UBF constructs were also expressed in E.coli JM 109 and bound to G-Sepharose matrix (Amersham Pharmacia Biotech). Aliquots of 30 µl of the G-Sepharose matrices carrying equal amounts of the recombinant proteins or GST alone were incubated with the respective in vitro translated protein. After 1 h incubation at 4°C the matrices were washed with PBS, the bound proteins resolved by 10% SDS–PAGE and the gel subjected to phosphorimaging.
ESI analysis of protein–DNA complexes
Each UBF mutant (1 µg) was incubated in 25 µl of 50 mM HEPES pH 7.6, 5 mM MgCl2, 80 mM KCl, 1 mM DTT with 200 ng X.laevis 1.1 kb BamHI enhancer DNA fragment (19,20). After 15 min at room temperature, the mixture was chromatographed on a 0.5 ml column of Sepharose CL-2B to separate DNA-bound UBF from free protein. The column buffer contained 10 mM HEPES pH 7.2, 5 mM MgCl2, 1% formaldehyde, 0.5% glutaraldehyde. The peak DNA fraction (5 µl) was placed on a 1000 mesh copper electron microscope grid, which had been coated with a 3 nm carbon film, and glow discharged immediately before use (21). After 30 s, excess sample was washed from the grid with H2O and the grid air dried after all but a thin layer of the H2O had been removed.
ESI analysis of DNA–protein complexes has been previously described (21,22). A brief description follows. Estimation of the masses of the UBF–DNA complexes was carried out on a reference image recorded at 120 eV in the electron energy loss spectrum. DNA was used as an internal mass standard and the mass of the complex was estimated by comparison of integrated optical density of the complex with the integrated optical density over a defined length of DNA. Net phosphorus images were obtained by subtraction of the 120 eV reference image from a 155 eV energy loss image recorded at the peak of the phosphorus L2,3 ionization edge, after alignment and normalisation. Results were compared quantitatively with a multiple parameter background correction using two pre-edge images recorded at 105 and 120 eV (21).
RESULTS
The UBFs all contain two short, lysine-rich AKK elements, where A stands for aliphatic (Gly or Leu in the known UBFs), linking the N-terminal dimerisation domain to the first HMG box domain (Fig. 2A). Similar basic peptides flanking the HMG boxes of other proteins have been shown to lie in the major groove stabilising the induced DNA bending (see for example 23). If we were correct in suggesting that the enhancesome was formed by in-phase DNA bending, the relative positioning along the DNA of the two UBF monomers should be crucial to maintain the phase of the DNA bends induced by each (Fig. 1). Since this positioning does not depend on the underlying DNA sequence (3) it must be defined by the N-terminal dimerisation domain and the peptide, including the AKK repeat, linking this domain to HMG box 1. It was thus argued that changes in the length of the AKK linker might result in a spacing change displacing one UBF monomer relative to the other along the DNA duplex. If the DNA loop of the enhancesome depended purely on in-phase bending, such a spacing change should disrupt DNA looping in the enhancesome. For example, displacing one of the two UBF molecules shown in Figure 1 by 1.7 nm (a half-helical turn) along the DNA should induce an ‘S’ bend in the DNA rather than a loop. Given that the AKK peptide repeat unit would have an extended length of nearly 1.2 nm, insertion or deletion of copies of this repeat in both UBF monomers of the enhancesome could cause a relative UBF displacement of up to two-thirds of a turn of the DNA duplex.
Figure 2.

(A) The domain structure of Xenopus UBF and the C-terminally truncated ‘core’ UBF, Nbox13. The sequence of the peptide linking the N-terminal dimerisation domain (Dimer.) and HMG box 1 is shown aligned with the N-terminal sequence of NHP6a, known to bind the narrowed major groove on the inside of the HMG box-induced DNA bend. Basic amino acids are shown in black and similarities between the sequences are boxed. (B) The wild-type (WT) and mutant (‘Minus’ or ‘Plus,) linkers used in the present study. (C) Analysis of the protein masses of complexes of Nbox13 (Minus and Plus mutants) bound to the 1.1 kb ribosomal enhancer DNA repeat from X.laevis (19) as determined by ESI. (D) Dimerisation of the free Core UBF mutants. [35S]methionine-labelled FLAG-tagged forms of wild-type and each core UBF mutant (Applied) were allowed to interact with the immobilised equivalent UBF form (Matrix). The protein retained by homodimerisation was analysed by SDS–PAGE followed by phosphorimaging (see Materials and Methods). Relative homodimerisation (Rel. Dimer) is given as recovery of bound 35S-labelled mutant protein relative to wild-type.
Both Plus and Minus linker mutations inhibit UBF dimerisation
Two different core UBF (NBox13) mutants were prepared. In the first one, an AKK linker element was deleted, producing the ‘Minus’ mutant, while in the second an extra AKK motif was added, producing the ‘Plus’ mutant (Fig. 2B). When these two forms of core UBF were complexed with DNA and the complexes quantitatively analysed by ESI, only a few contained an NBox13 dimer (expected mass 150 kDa), while most contained only a monomer of core UBF (Fig. 2C). Our data had suggested that the first three HMG boxes of UBF bound DNA in a colinear fashion (1; Fig. 1). The Minus deletion mutation should then bring the two core UBF molecules of the enhancesome closer together and hence might engender a topological problem. On the other hand, the Plus insertion mutation should not display such a problem. It was, therefore, surprising to find that exactly the opposite in fact occurred. The Plus mutation induced a significantly stronger propensity toward monomer complexes than did the Minus mutation (Fig. 2C). In the absence of DNA, each mutant protein was found to be capable of dimerisation (Fig. 2D). The Plus mutant, which showed poor dimerisation when DNA bound, homodimerised nearly as effectively as did the wild-type (85%). Homodimerisation of the free Minus mutant protein was less efficient than the wild-type (55%), showing that this mutation had affected dimerisation, as was suspected it might. However, 40% of the Minus mutant DNA complexes, but only 10% of the Plus mutant DNA complexes, contained a dimer of core UBF (Fig. 2C). It was, therefore, concluded that lengthening the AKK linker repeat selectively affected dimerisation of DNA-bound but not the free core UBF. This was consistent with this linker having an important topological role in the relative positioning of the two UBF molecules on the DNA.
Mutant UBF dimers bend but do not loop DNA
ESI micrographs of the complexes containing dimers of the AKK mutant UBFs showed that while each complex was associated with a DNA bend, none formed the near 360° DNA loop characteristic of the enhancesome (Fig. 3A). No crossing-over of incoming and outgoing DNA duplexes was evident in these complexes nor were the trajectories of these duplexes tangential with the periphery of the complexes, both criteria of the complete DNA loop that is observed in the wild-type enhancesome (Fig. 3B). (A much larger number of complexes were scanned visually without any showing a looped DNA structure.) This was consistent with the mutation having induced a phasing change between the DNA bends induced by each UBF monomer or in some other way having prevented correct DNA bending by each monomer. Though in no case was an S bend apparent, some complexes did show two distinct DNA bends (Fig. 3A, iv and v), consistent with the AKK mutations having induced a dephasing of the hemi-enhancesomes.
Figure 3.

Minus and Plus mutations of core UBF prevent the formation of the 360° DNA loop characteristic of the wild-type enhancesome. (A) Dimers of Minus and Plus core UBF mutants complexed with DNA. (B) Typical enhancesome complexes containing dimers of wild-type core UBF complexed to DNA. (A) and (B) show superimposed phosphorus (DNA in red) and total mass (grey) ESI images of wild-type and core UBF mutants bound to the 1.1 kb Xenopus ribosomal enhancer DNA. The calculated protein mass for each complex is given.
Complexes containing mutant UBF monomers resemble hemi-enhancesomes
Complexes containing monomers of the AKK UBF mutants were also associated with a clear DNA bend (Fig. 4A and B). In the case of both Minus and Plus mutants the complexes were often associated with a near perfect 180° DNA bend. Analysis of the DNA bend angles (Fig. 5) gave 179 ± 37° for the Minus mutant and 172 ± 16° for the Plus mutant, very close to the bend angle expected for a half enhancesome (175 ± 8°) (3). Thus, each UBF monomer of the enhancesome was able to independently bend DNA into a near 180° bend. This clearly showed that inter-molecular UBF interactions were unnecessary for the DNA looping of the enhancesome. It also showed that the enhancesome is formed by two completely independent hemi-enhancesome in-phase bends induced by UBF monomers.
Figure 4.

Monomer complexes of Minus and Plus core UBF mutants bound to the 1.1 kb Xenopus ribosomal enhancer DNA. (A) Monomer complexes formed by the Minus core UBF mutant. (B) Monomer Plus core UBF mutant complexes. Phosphorus and total mass images are shown superimposed as in Figure 3. The calculated protein mass for each complex is given.
Figure 5.
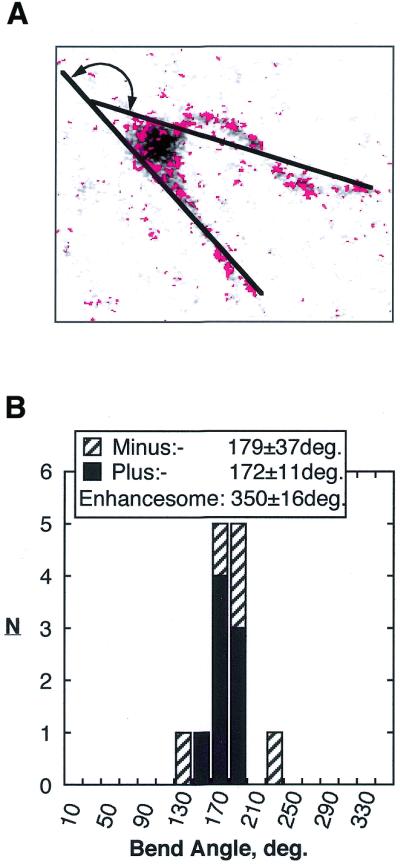
Monomer Minus and Plus core UBF mutant complexes were analysed for DNA bend angle associated with the complex. (A) A typical bend angle measurement on a monomer complex. (B) Histogram of bend angles observed for both Minus and Plus mutants. The angle expected for the wild-type enhancesome is given.
Constraints on relative HMG box positioning in the enhancesome
The spacing of the UBF monomers along the DNA is of key importance to the correct phasing of the bends they induce. Our present data suggest that this spacing is in part a function of the length of the linker between the dimerisation domain and HMG box 1. However, since each HMG box of UBF kinks the DNA independently of its neighbouring boxes (3), it is also essential that the boxes within each UBF molecule are exactly spaced along the DNA in order to maintain the relative phasing of their bends. We have previously shown that an UBF dimer induces a near 360° loop in ∼140 bp of DNA regardless of the underlying DNA sequence (3). Thus the information for exact spacing of the HMG boxes along the DNA and hence the phasing of the kinks they induce resides solely within the UBF molecule. Given this, we conclude that inter-box interactions and/or the inter-box linker peptides ensure relative box positioning and hence the phasing of DNA kinks. Thus, the inter-box linkers resemble in some aspects the dimerisation domain–box 1 AKK linker, which, as we have shown above, ensures hemi-enhancesome phasing. In support of this view, the inter-HMG box sequences of UBF from mammals to Xenopus are at least as highly, if not more highly, conserved than the HMG boxes themselves. This is especially evident for the 7 amino acid box 1–box 2 linker peptide (two conservative changes) and the 25 amino acid box 2–box 3 linker peptide (two conservative changes).
To maintain in-phase DNA bending the HMG boxes must bind the same face of the DNA. Structural analyses have shown that HMG box domains usually contact less than one turn of DNA. However, probably by steric hindrance, they protect 18–20 bp, i.e. up to two duplex turns, against DNase I attack (1,3,11,24). Thus, binding of an HMG box every turn of the DNA duplex may not be possible. In the core enhancesome we have shown that six HMG boxes of a UBF dimer protect and bend nearly 140 bp of DNA, strongly suggesting that the boxes bind every two DNA turns, (6 boxes × 2 turns × 10–10.6 bp/turn = 120–127 bp) (1,3). However, the length of the box 1–box 2 linker is insufficient to permit tandem binding of HMG boxes 1 and 2 to sites spaced by two DNA turns. The N- and C-termini of all HMG boxes lie proximal in the fold (Fig. 6A). Thus, the 7 amino acid box 1–box 2 linker (less than 3 nm fully extended) is too short for boxes 1 and 2 to position themselves tandemly along the DNA duplex with a two turn (∼6.8 nm) spacing, i.e. as in Figure 1. The most likely solution seems that HMG box 1 in fact lies head-to-head with box 2 when on DNA (Fig. 6B). Though to suggest such an inverted configuration is novel, it is not completely without precedent. The HMG box of NHP6a is able to bind the SRY DNA recognition sequence in the reverse orientation to the HMG box of SRY (23). The 25 amino acid linker between boxes 2 and 3 of UBF is then more than adequate to allow tandem binding of these boxes on DNA. Inverting HMG box 1 has some important implications. The N-terminal dimerisation domain of UBF could no longer lie at one end of an array of colinear HMG boxes but would link into this array between box 1 and box 2. However, by analogy with NHP6a, the AKK basic linker immediately N-terminal of box 1 might lie in the major groove, both stabilising the box 1-induced DNA bend and placing the dimerisation domains of a UBF dimer in closer proximity.
Figure 6.

Rethinking the enhancesome model to account for the length of interdomain peptide linkers. (A) The structure of a typical sequence non-specific HMG box fold, that of HMG-D (32). The three helices and the N- and C-termini (NH and COOH) are indicated. Note that the polypeptide chain enters and exits the HMG box fold proximally. (B) A schematic of the core enhancesome indicating the probable positions and orientations of the HMG boxes. The length of peptide linkers is given in amino acids (aa), the linkers themselves are indicated by broad arrows N- to C-terminally. Previous data shows that boxes 1 and 2 bind adjacent DNA sequences, enforcing an inversion of box 1 relative to box 2 and, hence, a head-to-head topology for these two boxes. The DNA is shown as a bent red ribbon and the HMG boxes are shown in blue using the HMG-D structure.
DISCUSSION
The enhancesome is a nucleoprotein formed when a dimer of the architectural HMG box transcription factor UBF interacts with 142 ± 30 bp of ribosomal DNA. It is similar in mass to the core nucleosome of chromatin, however, it loops the ribosomal DNA into a single superhelical turn of ∼17 nm diameter. Ribosomal transcription is regulated by many factors that determine cell growth and proliferation (12,13,25–29) and MAP kinase (V.Y.Stefanovsky, in preparation). Since each of these factors induces post-translational modification of UBF, they have the potential to affect enhancesome structure. Determination of this structure is, therefore, of key importance to our future understanding of ribosomal transcription regulation.
We have identified a basic tripeptide linker repeat (AKK, where A is an aliphatic residue) lying between the N-terminal dimerisation domain and the first HMG box DNA-binding domain of UBF. Modulating the length of this tripeptide repeat allowed us to observe hemi-enhancesomes. The data significantly advance our understanding of the enhancesome structure. Firstly, they show unequivocally that each UBF monomer can act non-cooperatively to induce a near 180° hemi-enhancesomal DNA bend. The enhancesome is therefore a modular structure consisting of two hemi-enhancesomes linked in-phase by the dimerisation domain of UBF. Secondly, the entering and exiting DNA duplexes need not be linked at their cross-over point in order to maintain the 360° enhancesomal DNA loop, as was previously suggested (14). Thirdly, the length of the basic inter-domain linker between the dimerisation domain and HMG box 1 is critical to enhancesome topology. The data suggest that the length of this linker peptide is critical for the relative spacing of the two UBF monomers in the enhancesome and hence determines the phasing of the 180° DNA bends they induce. By analogy, inter-HMG box linkers will most probably be critical for correct phasing of the DNA bends induced by the individual boxes. Consistent with this, UBF was inactivated when HMG boxes and inter-domain linkers were interchanged (30).
Consideration of the inter-HMG box domain peptides of the UBFs showed that HMG boxes 1 and 2 almost certainly do not bind tandemly along the enhancesomal DNA, but head-to-head (Fig. 6B). Initially this possibility seemed rather surprising, since it would appear to position the N-terminal dimerisation domain somewhat distal from the junction of the two UBF molecules. However, it is fully consistent with the symmetry of the DNA duplex and in fact HMG boxes have sometimes been observed to bind the same DNA duplex in opposite orientations (23). Several atomic resolution HMG box structures have also suggested that lysine-rich peptides, similar to the AKK repeats of UBF, lying immediately N- or C-terminal to an HMG box can cross the DNA backbone to lie in the narrowed major groove on the inner surface of the induced DNA bend (see for example 23,31). In UBF such a major groove interaction could serve to determine the position and topology of the dimerisation domains within the enhancesome (Fig. 6B). Folding of the AKK repeat peptide into the major groove on the inside of the DNA bend induced by HMG box 1 potentially places the dimerisation domain within the enhancesome loop, potentially explaining the central protein mass which we have consistently observed by ESI (2,3).
In the absence of a model of the enhancesome at atomic resolution, our molecular modelling from ESI data provides the sole basis for interpretation of the rapidly accumulating functional data on the role of UBF in ribosomal transcription regulation. It may also help to guide the design of new experiments.
Acknowledgments
ACKNOWLEDGEMENTS
This work was supported in Québec by an operating grant from the Medical Research Council of Canada (MRC-C, now the CIHR), a FCAR-FRSQ Santé (Québec) scholarship to G.P. and a MRC-C Scientist award to T.M and in Calgary by an operating grant from the Natural Sciences and Engineering Research Council to D.P.B.-J. The Centre de Recherche en Cancerologie de l’Université Laval is supported by the Fonds de Recherche sur la Santé de Québec (FRSQ).
References
- 1.Leblanc B., Read,C. and Moss,T. (1993) Recognition of the Xenopus ribosomal core promoter by the transcription factor xUBF involves multiple HMG box domains and leads to an xUBF interdomain interaction. EMBO J., 12, 513–525. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 2.Bazett-Jones D.P., Leblanc,B., Herfort,M. and Moss,T. (1994) Short-range DNA looping by the Xenopus HMG-box transcription factor, xUBF. Science, 264, 1134–1137. [DOI] [PubMed] [Google Scholar]
- 3.Stefanovsky V.Y., Bazett-Jones,D.P., Pelletier,G. and Moss,T. (1996) The DNA supercoiling architecture induced by the transcription factor xUBF requires three of its five HMG-boxes. Nucleic Acids Res., 24, 3208–3215. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 4.Moss T. and Stefanovsky,V.Y. (1995) Promotion and regulation of ribosomal transcription in eukaryotes by RNA Polymerase I. In Cohn,W.E. and Moldave,K. (eds), Progress in Nucleic Acids and Molecular Biology. Academic Press, San Diego, CA, pp. 25–66. [DOI] [PubMed]
- 5.Moss T., Stefanovsky,V.Y. and Pelletier,G. (1998) The structural and architectural role of Upstream Binding Factor, UBF. In Paule,M.R. (ed.), Transcription of Ribosomal Genes by Eukaryotic RNA Polymerase I. Landes Bioscience, Austin, TX, pp. 75–94.
- 6.Hempel W.M., Cavanaugh,A.H., Hannan,R.D., Taylor,L. and Rothblum,L.I. (1996) The species-specific RNA polymerase I transcription factor SL-1 binds to upstream binding factor. Mol. Cell. Biol., 16, 557–563. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 7.Kihm A.J., Hershey,J.C., Haystead,T.A.J., Madsen,C.S. and Owens,G.K. (1998) Phosphorylation of the rRNA transcription factor upstream binding factor promotes its association with TATA binding protein. Proc. Natl Acad. Sci. USA, 95, 14816–14820. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 8.Sullivan G.J. and McStay,B. (1998) Dimerization and HMG box domains 1–3 present in Xenopus UBF are sufficient for its role in transcriptional enhancement. Nucleic Acids Res., 26, 3555–3561. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 9.Tuan J.C., Zhai,W.G. and Comai,L. (1999) Recruitment of TATA-binding protein-TAF complex SL1 to the human ribosomal DNA promoter is mediated by the carboxy-terminal activation domain of upstream binding factor (UBF) and is regulated by UBF phosphorylation. Mol. Cell. Biol., 19, 2872–2879. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 10.McStay B., Frazier,M.W. and Reeder,R.H. (1991) xUBF contains a novel dimerization domain essential for RNA polymerase I transcription. Genes Dev., 5, 1957–1968. [DOI] [PubMed] [Google Scholar]
- 11.Jantzen H.M., Chow,A.M., King,D.S. and Tjian,R. (1992) Multiple domains of the RNA polymerase I activator hUBF interact with the TATA-binding protein complex hSL1 to mediate transcription. Genes Dev., 6, 1950–1963. [DOI] [PubMed] [Google Scholar]
- 12.Pelletier G., Stefanovsky,V.Y., Faubladier,M., Hirschler-Laszkiewicz,I., Savard,J., Rothblum,L.I., Côté,J. and Moss,T. (2000) Competitive recruitment of CBP and Rb-HDAC regulates UBF acetylation and ribosomal transcription. Mol. Cell, 6, 1059–1066. [DOI] [PubMed] [Google Scholar]
- 13.Voit R., Hoffmann,M. and Grummt,I. (1999) Phosphorylation by G1-specific cdk-cyclin complexes activates the nucleolar transcription factor UBF. EMBO J., 18, 1891–1899. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 14.Hu C.H., McStay,B., Jeong,S.W. and Reeder,R.H. (1994) xUBF, an RNA polymerase I transcription factor, binds crossover DNA with low sequence specificity. Mol. Cell. Biol., 14, 2871–2882. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 15.Pöhler J.R.G., Norman,D.G., Bramham,J., Bianchi,M.E. and Lilley,D.M.J. (1998) HMG box proteins bind to four-way DNA junctions in their open conformation. EMBO J., 17, 817–826. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 16.Lilley D.M. and Clegg,R.M. (1993) The structure of the four-way junction in DNA. Annu. Rev. Biophys. Biomol. Struct., 22, 299–328. [DOI] [PubMed] [Google Scholar]
- 17.Smith D.B. and Corcoran,L.M. (1991) Protein expression. In Ausubel,F.M., Brent,R., Kingston,R.E., Moore,D.D., Seidman,J.G., Smith,J.A. and Struhl,K. (eds), Current Protocols in Molecular Biology. Greene Publishing Associates & Wiley-Interscience, New York, NY.
- 18.Bachvarov D. and Moss,T. (1991) The RNA polymerase I transcription factor xUBF contains 5 tandemly repeated HMG homology boxes. Nucleic Acids Res., 19, 2331–2335. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 19.Moss T., Boseley,P.G. and Birnstiel,M.L. (1980) More ribosomal spacer sequences from Xenopus laevis. Nucleic Acids Res., 8, 467–485. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 20.De Winter R.F.J. and Moss,T. (1987) A complex array of sequences enhances ribosomal transcription in Xenopus laevis. J. Mol. Biol., 196, 813–827. [DOI] [PubMed] [Google Scholar]
- 21.Bazett-Jones D.P. (1993) Empirical basis for phosphorous mapping and structure determination of DNA:protein complexes by electron spectroscopic imaging. Microbeam Anal., 2, 69–79. [Google Scholar]
- 22.Bazett-Jones D.P. and Brown,M.L. (1988) Electron microscopy shows that TFIIIA bends DNA. Mol. Cell. Biol., 9, 336–341. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 23.Allain F.H., Yen,Y.M., Masse,J.E., Schultze,P., Dieckmann,T., Johnson,R.C. and Feigon,J. (1999) Solution structure of the HMG protein NHP6A and its interaction with DNA reveals the structural determinants for non-sequence-specific binding. EMBO J., 18, 2563–2579. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 24.Giese K., Amsterdam,A. and Grosschedl,R. (1991) DNA-binding properties of the HMG domain of the lymphoid-specific transcriptional regulator LEF-1. Genes Dev., 5, 2567–2578. [DOI] [PubMed] [Google Scholar]
- 25.Cavanaugh A.H., Hempel,W.M., Taylor,L.J., Rogalsky,V., Todorov,G. and Rothblum,L.I. (1995) Activity of RNA polymerase I transcription factor UBF blocked by Rb gene product. Nature, 374, 177–180. [DOI] [PubMed] [Google Scholar]
- 26.Voit R., Schäfer,K. and Grummt,I. (1997) Mechanism of repression of RNA polymerase I transcription by the retinoblastoma protein. Mol. Cell. Biol., 17, 4230–4237. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 27.Voit R., Schnapp,A., Kuhn,A., Rosenbauer,H., Hirschmann,P., Stunnenberg,H.G. and Grummt,I. (1992) The nucleolar transcription factor mUBF is phosphorylated by casein kinase II in the C-terminal hyperacidic tail which is essential for transactivation. EMBO J., 11, 2211–2218. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 28.O’Mahony D.J., Xie,W., Smith,S.D., Singer,H.A. and Rothblum,L.I. (1992) Differential phosphorylation and localisation of the transcription factor UBF in vivo in response to serum deprivation. J. Biol. Chem., 267, 35–38. [PubMed] [Google Scholar]
- 29.O’Mahony D.J., Smith,S.D., Xie,W. and Rothblum,L.I. (1992) Analysis of the phosphorylation, DNA-binding and dimerization properties of the RNA polymerase I transcription factors UBF1 and UBF2. Nucleic Acids Res., 20, 1301–1308. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 30.Cairns C. and McStay,B. (1995) HMG box 4 is the principal determinant of species specificity in the RNA polymerase I transcription factor UBF. Nucleic Acids Res., 23, 4583–4590. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 31.Lnenicek-Allen M., Read,C.M. and Crane-Robinson,C. (1996) The DNA bend angle and binding affinity of an HMG box increased by the presence of short terminal arms. Nucleic Acids Res., 24, 1047–1051. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 32.Murphy F.V., Sweet,R.M. and Churchill,M.E. (1999) The structure of a chromosomal high mobility group protein–DNA complex reveals sequence-neutral mechanisms important for non-sequence-specific DNA recognition. EMBO J., 18, 6610–6618. [DOI] [PMC free article] [PubMed] [Google Scholar]