

Abstract
Choline is an essential nutrient for humans. Metabolically choline is used for the synthesis of membrane phospholipids (e.g. phosphatidylcholine), as a precursor of the neurotransmitter acetylcholine, and, following oxidation to betaine, choline functions as a methyl group donor in a pathway that produces S-adenosylmethionine. As a methyl donor choline influences DNA and histone methylation – two central epigenomic processes that regulate gene expression. Because the fetus and neonate have high demands for choline, its dietary intake during pregnancy and lactation is particularly important for normal development of the offspring. Studies in rodents have shown that high choline intake during gestation improves cognitive function in adulthood and prevents memory decline associated with old age. These behavioral changes are accompanied by electrophysiological, neuroanatomical, and neurochemical changes and by altered patterns of expression of multiple cortical and hippocampal genes including those encoding key proteins that contribute to the biochemical mechanisms of learning and memory. These actions of choline are observed long after the exposure to the nutrient ended (months) and correlate with fetal hepatic and cerebral cortical choline-evoked changes in global- and gene-specific DNA cytosine methylation and with dramatic changes of the methylation pattern of lysine residues 4, 9 and 27 of histone H3. Moreover, gestational choline modulates the expression of DNA (Dnmt1, Dnmt3a) and histone (G9a/Ehmt2/Kmt1c, Suv39h1/Kmt1a) methyltransferases. In addition to the central role of DNA and histone methylation in brain development, these processes are highly dynamic in adult brain, modulate the expression of genes critical for synaptic plasticity, and are involved in mechanisms of learning and memory. A recent study documented that in a cohort of normal elderly people, verbal and visual memory function correlated positively with the amount of dietary choline consumption. It will be important to determine if these actions of choline on human cognition are mediated by epigenomic mechanisms or by its influence on acetylcholine or phospholipid synthesis.
Keywords: brain, choline, DNA, histone, memory, methylation, nutrition, pregnancy
Choline was first described in 1862 by Strecker [1], who found it in bile, and used the Greek word for bile to name the compound. Determination of the chemical structure of choline and its first chemical synthesis was performed shortly thereafter [2] and in 1868 Strecker found that choline was a part of the molecule of phosphatidylcholine. We now know that phosphatidylcholine is the most abundant structural component of biological membranes. Choline also serves as the precursor of the neurotransmitter, acetylcholine (ACh). The fascinating history of discovery of physiological functions of ACh spans many decades and includes the first demonstration of chemical neurotransmission [see refs. [3,4] for the 1936 Nobel lectures by Otto Loewi and Sir Henry Dale]. In 1937 Mann and Quastel described the process of enzymatic oxidation of choline to betaine aldehyde and betaine [5] and in 1946 du Vigneaud et al showed that betaine serves as a donor of metabolic methyl groups, and suggested that the process of methyl group transfer is indirect and occurs via a methionine intermediate [6]. Indeed, in 1953 Cantoni determined the structure of such an intermediate as S-adenosylmethionine (AdoMet) [7]. Most methylating enzymes use AdoMet as the donor of methyl groups [8,9] generating a methylated product (e.g. 5-methylcytosine in DNA [10] or N-methylated lysine in a histone protein [11]) and S-adenosylhomocysteine (AdoHcy) [8,9]. This metabolic relationship between choline and DNA methylation is illustrated in Figure 1. The essential function of dietary choline in preserving normal health in mammals was recognized already in the 1930s initially in the context of studies on pancreas and insulin. It was noted that dogs with surgically removed pancreas, but treated with insulin, developed fatty liver and later died. Feeding with raw pancreas prevented liver disease [12] and the active ingredient of this treatment was subsequently identified as choline [13–15] by, among others, one of the discoverers of insulin, Charles Herbert Best [14,15].
Figure 1.
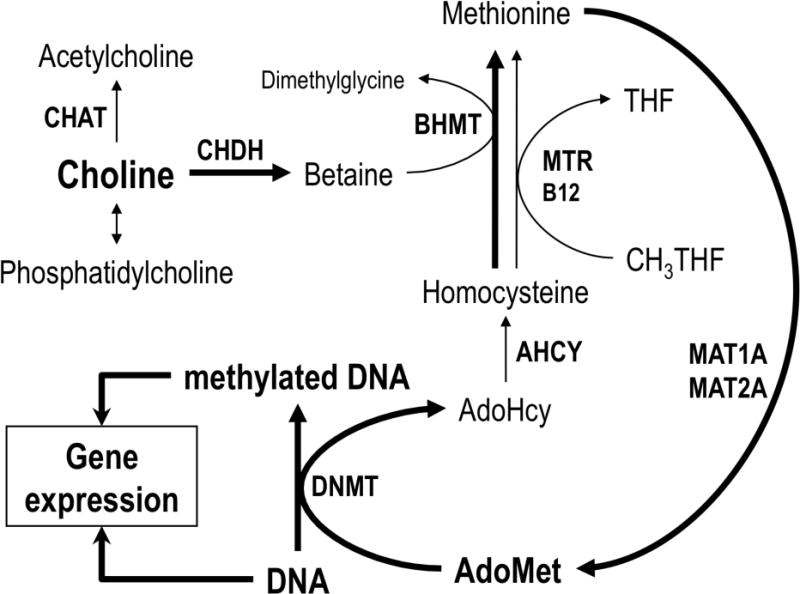
Choline and methyl group metabolism and its relation to DNA methylation and influence on regulation of gene expression: a simplified diagram. Choline is used as a precursor of phosphatidylcholine, acetylcholine [in a reaction catalyzed by choline acetyltransferase (CHAT)], or betaine [in a reaction catalyzed by choline dehydrogenase (CHDH)]. The methyl groups of betaine are used by betaine:homocysteine S-methyltransferase (BHMT) to regenerate methionine from homocysteine. In an alternative pathway, catalyzed by vit. B12-requiring 5-methyltetrahydrofolate-homocysteine S-methyltransferase (MTR), methyltetrahydrofolate (CH3THF) is used as a methyl donor. Methionine is used as a precursor of S-adenosylmethionine (AdoMet) in a reaction catalyzed by methionine adenosyltransferase(s) (MAT1A, MAT2A or MAT2B). AdoMet is used by multiple methylating enzymes including DNA methyltransferases (DNMT1, DNMT3A, DNMT3B) that use AdoMet as a donor of methyl groups to methylate DNA at the 5-position of cytosine residues within the CpG sequences. The DNA methylation state and pattern exerts a modulatory influence on expression of multiple genes (see text). The second product of this, and all other AdoMet-requiring methylation reactions, S-adenosylhomocysteine (AdoHcy) is hydrolyzed to free homocysteine by AdoHcy hydrolase (AHCY). The metabolic pathway linking choline to DNA methylation is indicated by thick arrows.
1. Choline, an essential nutrient for humans
Remarkably, we had to wait another half a century for the definitive proof that choline was an essential nutrient for humans. In a 1991 rigorous clinical study, Zeisel et al placed a group of healthy men on a low choline diet and found that they developed liver damage that could be reversed with a standard dose of dietary choline [16]. This landmark discovery served as the initial basis for the classification of choline as an essential nutrient in 1998 when the Food and Nutrition Board (FNB) of the Institute of Medicine of the National Academy of Sciences of the United States of America issued guidelines on its daily intake [17]. Because at that time, and perhaps to this day, there were insufficient data to generate Recommended Daily Allowance values, the FNB issued Adequate Intake (AI) recommendations (Table 1). The AI calls for the average intake of 7.5 mg of choline daily per kg of body weight. Notably, the AI is increased for pregnant and breast feeding women in order to satisfy the needs of the fetus and baby whose choline is supplied via placenta [18] and milk [19,20], respectively. The AI values were established primarily to ensure that dietary choline is sufficient to prevent liver dysfunction associated with low choline consumption observed in adult men [16] (see above). Subsequent studies have shown that choline deficiency also causes muscle damage [21] and induces apoptotic death of lymphocytes [22].
Table 1.
Choline Adequate Intake (AI) (mg/day)
| Life stage | Age | Females | Males |
|---|---|---|---|
| Infants | 0–6 months | 125 | 125 |
| Infants | 7–12 months | 150 | 150 |
| Children | 1–3 years | 200 | 200 |
| Children | 4–8 years | 250 | 250 |
| Children | 9–13 years | 375 | 375 |
| Adolescents | 14–18 years | 400 | 550 |
| Adults | 19 years and older | 425 | 550 |
| Pregnancy | All ages | 450 | – |
| Breast-feeding | All ages | 550 | – |
2. Nutrigenetics and nutrigenomics of choline in humans
Since the publication of the FNB report, new research data indicate that even in a developed country like the United States, choline nutrition may be low among surprisingly large segments of population [23] and that additional factors including sex, age, and genotype influence the individual’s requirements for choline. These factors constitute the purview of three relatively new sub disciplines in the science of nutrition and genetics, namely: 1) nutrigenetics – how the genome of an organism governs its response to a nutrient; 2) nutrigenomics – how nutrients interact with the genome to modulate gene expression patterns; and 3) nutriepigenomics – how a nutrient influences the epigenome and thus modulates gene expression patterns. Recent observations in the area of choline nutrigenetics showed that non-pregnant, non-nursing, women of childbearing age are relatively resistant to choline deficiency as compared to men and older, postmenopausal women [24]. This is explained by the apparently upregulated endogenous biosynthesis of choline (in the form of phosphatidylcholine) in younger women catalyzed by the hepatic enzyme phosphatidylethanolamine N-methyltransferase (PEMT). PEMT activity constitutes the sole mechanism whereby mammals can synthesize choline de novo. Note, that the reaction proceeds as follows:
Thus, AdoMet can be produced from choline via betaine and methionine, as described above (Figure 1), but it can also be consumed to generate new choline molecules. Interestingly, it was the presence of PEMT activity in humans that served as the key argument against including choline on the list of essential nutrients. We now know from studies in mice that PEMT is vitally important but its activity is insufficient to create adequate amounts of choline for the maintenance of normal health [25–27]. Remarkably, expression of the PEMT gene is increased by estrogens. In younger women, high levels of circulating estrogen are responsible for inducing and maintaining high PEMT activity, and thus rendering the women relatively insensitive to low-choline diets [28,29]. This, apparently adaptive genomic mechanism, may buffer both the mother and her offspring during gestation and lactation from the detrimental effects of short supply of dietary choline. In contrast, postmenopausal women are sensitive to low-choline diets and deplete their choline pools in a similar fashion to men.
There are several polymorphic genes that affect humans’ requirement for choline, including certain alleles of PEMT [29,30]. One of those alleles (polymorphism rs12325817) apparently renders the PEMT gene unresponsive to estrogen conferring high risk of development of choline deficiency-associated liver dysfunction in women carriers on normal diets [31,32]. Additional genetic polymorphisms that modify choline requirements include alleles of 5,10-methylenetetrahydrofolate dehydrogenase (MTHFD1) [33], methyltetrahydrofolate reductase (MTHFR) [34–36] and choline dehydrogenase (CHDH) [29]. Moreover, it is possible to use metabolomic “signatures” in plasma to identify prospectively individuals at risk of developing liver or muscle dysfunction caused by choline deficiency [37]. Taken together, these findings point to a significant heterogeneity of choline metabolism, and thus dietary needs for choline, among humans.
The establishment of the United States Department of Agriculture (USDA) Database for the Choline Content of Common Foods [38] – one of the most valuable resources for epidemiological studies on choline nutrition – has helped to understand this heterogeneity in the metabolism of choline. Recent studies using this tool found that high intake of choline and betaine during pregnancy was associated with lowered risk of orofacial clefts in infants [39]. Moreover, polymorphisms in genes encoding betaine:homocysteine methyltransferase (BHMT) (Figure 1) and CTP:phosphocholine cytidylyltransferase (PCYT1) (the rate-limiting enzyme in phosphatidylcholine synthesis from choline) were associated with altered risk of orofacial clefts [40], further supporting the role of choline metabolism and nutrition in this developmental disorder. The use of the USDA database also revealed that women in the highest quintile of choline and betaine consumption as adults had reduced risk of breast cancer [41] and that high betaine intake lowers the risk of colorectal adenoma in women [42] and of esophageal cancer in both men and women [43]. Moreover, high choline consumption during pregnancy reduced the risk of neural tube defects in offspring [44,45]. These studies are the first to provide evidence for a preventive action of choline in human carcinogenesis and for the significance of choline nutrition during pregnancy for normal development of the human central nervous system. The latter conclusion is supported by the findings that polymorphisms in genes encoding two enzymes of phosphatidylcholine synthesis, PCYT1A and choline kinase A (CHKA) modify the risk of neural tube defects [46].
3. Choline nutrition and programming of brain development and cognitive function in animal models
In rodent models, maternal choline consumption has profound and long-term effects on brain development and cognition in offspring. Choline deficiency during pregnancy inhibits fetal cell proliferation in the hippocampus and stimulates apoptosis in this brain region [47,48]. In contrast, gestational choline supplementation stimulates hippocampal cell division [49]. Moreover, in a commonly used model that employs the offspring of pregnant mice or rats consuming diets of varying choline content during only a one week period of the second half of gestation (embryonic days 11–17; pregnancy in mice and rats lasts 19–21 and 20–22 days, respectively), choline deficiency causes impairments in certain memory tasks [50], whereas choline supplementation improves memory and attention [50–57] and, remarkably, prevents age-related memory decline [50,57]. The two salient conclusions from these studies are: 1. choline nutrition during gestation programs brain development and cognitive ability, and 2. cognitive decline is not an inevitable outcome of old age, but rather can be prevented by increased supply of choline during a critical period of prenatal development.
The above-noted structural changes in prenatal brain subsequently translate into neuroanatomical, neurochemical, electrophysiological, and molecular differences in the adult and aged animal. Certain aspects of learning and memory require continual production of new neurons that occurs in the dentate gyrus of the hippocampus throughout the lifetime [58,59]. Prenatal choline supplementation enhances adult neurogenesis in the dentate gyrus while prenatal choline deficiency impairs this process [60–62]. Moreover, the effect of choline supplementation was seen even in aged rats and correlated with a highly trophic microenvironment within the hippocampus of the prenatally choline supplemented rats that included elevated concentrations of brain-derived neurotrophic factor (BDNF), nerve growth factor (NGF), insulin-like growth factor I (IGF1), IGF2, and vascular endothelial growth factor (VEGF) in these animals as compared to controls [60–64]. Prenatal choline supplementation increases the size of the basal forebrain cholinergic neurons [65] that are critical for the processes of learning and memory [66,67], and augments ACh synthesis and release from these neurons [57,68]. In addition, prenatal choline availability alters the activation levels of essential molecular components of memory processing [69], such that phosphorylation of hippocampal mitogen-activated protein kinase (MAPK) and cAMP response element binding protein (CREB) in response to stimulation by glutamate, N-methyl-D-aspartate (NMDA), or depolarizing concentrations of potassium is increased by prenatal choline supplementation and reduced by prenatal choline deficiency [51]. Concordant changes were observed in hippocampal electrophysiological synaptic plasticity measures termed long-term potentiation (LTP). Prenatal choline supplementation enhanced hippocampal LTP in the CA1 region by decreasing the stimulus intensity required for LTP induction [70,71], possibly as a result of an augmented NMDA receptor-mediated neurotransmission [72]. Mellott et al [73] analyzed gene expression patterns in brains of prenatally choline-deficient, choline-supplemented, and control rats using microarrays and found 530 hippocampal and 815 cerebral cortical mRNA species whose levels were modulated by prenatal choline status. The protein products of several of these genes are known to participate in signaling pathways implicated in memory processes and thus may mediate the observed choline-induced changes in LTP and behavior. The list of these genes includes Igf2, protein kinases Prkcb2/PKCβ2, Camk1, and Camk2b/CaMKIIβ, a receptor for the neurotransmitter γ-aminobutyric acid (Gabbr1/GABABR1), and a transcription factor Zif268 (Egr1) [73]. These long-term actions on brain structure and function of altered availability of choline during gestation suggest that they may be mediated by epigenetic mechanisms that include DNA- and histone methylation.
4. DNA methylation
Recent advances in the field of epigenetics have provided the conceptual and experimental framework that helps to explain how some phenotypes can be transmitted following cell mitosis or across generations in complex organisms, including mammals, without a change in primary DNA sequence. The central molecular mechanism that permits this type of inheritance is methylation of DNA at the 5-position of cytosine residues within CpG sequences to form 5-methylcytosine (5mC) (Figure 2). The transcription of genes whose regulatory elements are methylated tends to be different than when the same regions are not methylated due to a concerted change in the interaction of those elements with a complex network of proteins, including transcription factors (see below). This change results in altered phenotype governed by DNA methylation. The pattern of DNA methylation can be propagated through cell divisions because, after DNA replication, the unmethylated daughter strand in hemimethylated DNA becomes symmetrically methylated by the enzyme DNA methyltransferase 1 (DNMT1). The process of DNA methylation is dynamic [74] and responds to the environment, including the availability of nutrients. In particular, DNA methylation is modulated by the availability of nutrients that serve as methyl group donors and cofactors, such as choline, betaine, methionine, folic acid and vitamin B12 (Figure 1). This effect is explained by the direct relationship between dietary intake of choline (and/or other methyl groups) and tissue levels of AdoMet that is frequently observed [75–82]. In proof-of-concept studies it has been shown that very high dietary intake of these compounds in pregnant mice can modify methylation and expression of certain genes and result in modified phenotypes of the offspring [83–88]. Moreover, some of these effects can be transmitted to successive generations [88,89]. These relatively recent discoveries, however, are based on a long history of investigations of the only nutritional deficiency that causes cancer, namely liver tumors (hepatocarcinoma) in rats fed a choline-deficient diet [90–96]. While many hypotheses for the mechanism of this effect had been advanced, one included the idea that dietary choline (or methyl group) deficiency may result in altered methylation of DNA and thus altered expression of proto oncogenes and tumor suppressor genes because dysregulated DNA methylation is a common pathophysiological feature of carcinogenesis [97–100]. Indeed, abnormalities in DNA methylation have been observed in the preneoplastic lesions and in liver tumors of choline- and methyl-group deficient rats [101–114].
Figure 2.
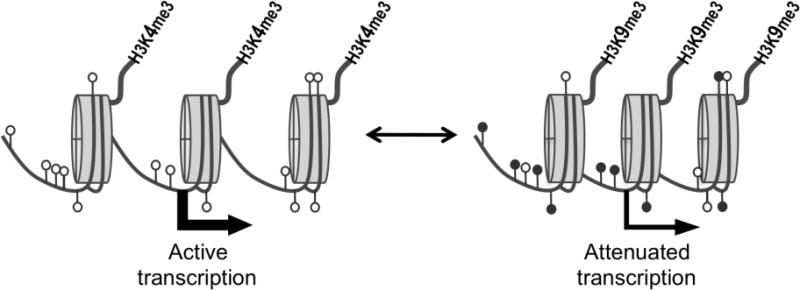
Methylation of DNA and histone H3 modifies chromatin state and regulates transcription. DNA is wrapped around nucleosomes that are composed of a histone octamer containing two molecules of H3. On the left cytosines within CpGs of DNA are unmethylated (white lollipops) and lysine 4 of the H3 N-terminal tail (thick line above the nucleosomes) is methylated (H3K4me3). Under these conditions transcription tends to be active (thick arrow). On the right, DNA has become more methylated (black lollipops) – initially due to the de novo process catalyzed by the DNMT3 enzymes and subsequently maintained in the cell lineage by DNMT1, and H3 is methylated on lysine 9 (H3K9me3). Under these conditions transcription is attenuated (thin arrow). Additional processes, including histone deacetylation, attachment of methylated DNA binding proteins (e.g. MECP2) also contribute to this transcriptional repression and chromatin compaction (illustrated on the right by closer packing of the nucleosomes). The two states are reversible.
DNA methylation is also important for such regulatory mechanisms of gene expression as X chromosome inactivation, cell differentiation and chromatin structure, the expression changes that occur during aging and genomic imprinting [115–119]. There is some evidence that the latter process may be modified by choline/methyl group availability [83,120]. In general, CpG methylation causes transcriptional repression [121–129] by recruiting specific proteins (MECP2, MBD1, MBD2, and MBD3) that bind to the methylated DNA via their methylated–CpG binding domains (MBD) [130]. Upon binding to methylated CpG sites, they further recruit transcriptional corepressors and histone deacetylases. Deacetylated nucleosomes are tightly packed which prevents the access of transcriptional activators to their binding sites in the DNA [reviewed in [122,125,131]]. CpG methylation also contributes to transcriptional repression by specifically preventing transcriptional regulators from binding to their target gene promoters. For example cMyb [132], E2F [133], CREB [134], AP2 [135], NF-κB [136], and STAT3 [129], cannot bind to methylated forms of their DNA recognition sequences. However, DNA methylation may also upregulate transcription by preventing the binding of a transcriptional repressor to a genomic silencer element, as is the case for Igf2 gene [137,138]. DNA methylation is catalyzed by a family of DNA methyltransferase enzymes. The mammalian brain has high levels of expression of DNA methyltransferase 1 (DNMT1) both during development and in adulthood [139–142] and the degree of DNA methylation is higher in the adult brain than in other tissues [143–145]. Moreover, DNA methylation levels in mouse brain undergo dynamic changes perinatally [143], suggesting that DNA methylation is necessary for the differentiation process of the brain. In human brain DNA methylation for most genomic loci tends to increase with age with some loci becoming hypermethylated during childhood, while others after the age of 50, and still others becoming monotonically hypermethylated throughout life until advanced age [146,147]. Human neurons have a distinct DNA methylation pattern as compared to non-neuronal brain cells and this neuronal DNA methylation pattern is characterized by a large individual variation [148]. Interestingly the expression of genes encoding enzymes of methyl group metabolism and DNA and protein methylation undergoes dramatic changes during brain development as illustrated in Figure 3, depicting the developmental pattern of their mRNA expression in rat frontal cortex. Note, that the expression of DNA methyltransferase, Dnmt1, and of protein methyltransferases, G9a/Ehmt2/Kmt1c (that dimethylates histone H3 on lysine 9; H3K9 to H3K9me2 and trimethylates H3K27 to H3K27me3 [149],) and Prmt1 (that methylates arginine residues in multiple protein substrates [150]) are high in mid-gestation and decline rapidly during the latter part of fetal development. A similar pattern of Dnmt3a and Dnmt3b expression in mouse brain has been reported [151]. The maturation of mRNA expression of two genes encoding enzymes that provide metabolic support for the processes of DNA and protein methylation by synthesizing AdoMet (i.e. Mat2a) and the hydrolysis of its demethylated metabolite, AdoHcy (i.e. Ahcy), follows a similar pattern.
Figure 3.
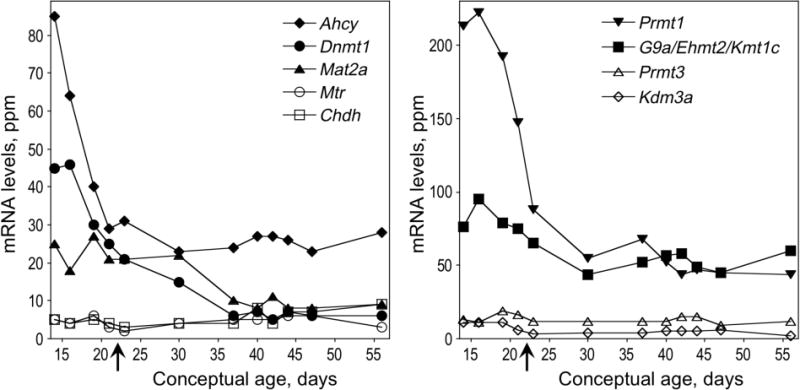
Developmental pattern of expression of selected choline- and methylation-related genes (see Figure 1) in rat frontal cortex. Cortical RNA was analyzed using Affymetrix RG_U34A microarrays as described [73]. Left panel: Chdh, choline dehydrogenase; Mtr, 5-methyltetrahydrofolate-homocysteine S-methyltransferase; Mat2a, methionine adenosyltransferase 2a; Dnmt1, DNA methyltransferase 1; Ahcy, S-adenosylhomocysteine hydrolase. Right panel: Kdm3a/Jmjd1a, lysine (K)-specific demethylase 3A; Prmt3, protein arginine N-methyltransferase 3; G9a/Ehmt2/Kmt1c, euchromatic histone-lysine N-methyltransferase 2; Prmt1, protein arginine N-methyltransferase 1. X axis on the graph corresponds to days post-conception. The day of birth is indicated by an arrow. Postnatal days 1, 8, 15, 18, 22, and 34 are conceptual ages 23, 30, 37, 40, 44, and 56, respectively.
As noted above, DNA methylation patterns acquired during development may be inherited through the cell divisions in a process catalyzed by DNMT1 that methylates hemimethylated CpG sites and thus restores the parental methylation pattern on the daughter DNA strand following DNA replication [152]. In addition, DNMT3A and DNMT3B generate the DNA methylation patterns de novo during development and in adulthood [153,154]. The observed effects of choline- or methyl group deficiency on global- and gene-specific DNA methylation in multiple in vivo and in vitro models vary: several studies reported hypomethylation [101,108,155–163], some no change [164], and some hypermethylation [165–168]. It is noteworthy, however that only some CpG sites respond to the altered availability of methyl groups, suggesting a degree of specificity of this response, that heretofore remains unexplained. Together the results indicate that DNA methylation may respond to the supply of methyl groups in a complex fashion that includes alterations in the activities of DNA methylating and/or demethylating enzymes. Indeed, diet can influence the expression of DNMTs. For example, Fischer male rats fed a methyl-group deficient diet for at least 3 weeks display global DNA hypomethylation and increased expression and activity of cellular DNMTs, (DNMT1 and DNMT3A) possibly as a compensatory mechanism that offsets methyl deficiency [169].
The hypothesis that choline intake by pregnant rats might alter DNA methylation and DNMT expression in the fetus was tested in a study by Kovacheva et al [82] who measured these parameters in liver and cerebral cortex on embryonic day 17 in rats following altered dietary supply of choline that had commenced on gestational day 11. The investigators focused on the differentially methylated region 2 (DMR2) of the Igf2 gene because the DMR2 methylation signature changes dramatically during development [170]. Choline-deficient embryos had higher degree of methylation as compared to the control and choline-supplemented rats. One possible mechanism that leads to changes in the global, as well as gene-specific, DNA methylation is via alteration in the activity of DNMTs. It has been shown that DNMT1 is important for maintaining the methylation pattern of the Igf2 gene and Dnmt1 knockout mice have abnormal expression of Igf2 [171]. In liver of choline-deficient embryos, Dnmt1 mRNA was overexpressed by more than 50% as compared to control and choline-supplemented embryos. The data suggested that maternal choline deficiency causes an apparently compensatory induction of Dnmt1 expression in the fetus that prevents the loss of DNA methylation when choline is in short supply. While the focus on the Igf2 gene was driven by the data showing that IGF2 expression in developing and adult brain was governed by prenatal choline intake [64,73,82] and by extensive literature on its regulation by DNA methylation [137,138,172], IGF2 has recently emerged as a critical component of memory consolidation mechanisms [173] suggesting that high IGF2 levels observed in brains of prenatally choline-supplemented rats [64,73] may be part of the mechanism of cognitive enhancement that characterizes these animals.
As noted above, DNA methylation is a dynamic process. Whereas in dividing cells loss of DNA methylation can be accomplished by the failure of DNMT1 to copy the methylation pattern of the mother DNA strand onto the daughter strand, in postmitotic cells, such as neurons, demethylation requires an enzymatic pathway. Recent studies suggest that this process entails initial obligatory conversion of 5mC to 5-hydroxymethylcytosine (5hmC) catalyzed by the Ten-eleven translocation (Tet) family of methyldioxygenases (TET1, TET2, and TET3) that require 2-oxoglutarate and iron Fe- (II) [174–176]. The subsequent enzymatic steps are poorly understood. There is evidence that, in the brain, the activation-induced deaminase (AID)/apolipoprotein B mRNA-editing enzyme complex (APOBEC) family of cytidine deaminases and base excision repair pathway is operational [177]. However, the stepwise oxidation of 5mC to 5-formylcytosine (5fC) and subsequently to 5-carboxylcytosine (5caC) catalyzed by the TET enzymes has also been observed [178,179] and 5caC could be simply decaboxylated to cytosine in a fashion analogous to the conversion of thymine to uracil [178]. Alternatively 5caC could be processed by thymine-DNA glycosylase in base excision repair pathway [179]. Functionally, the formation of 5hmC may be as significant as demethylation because methylated DNA binding proteins including MECP2 [180] MBD1, MBD2 and MBD4 [181] bind poorly to 5hmC. The dynamic DNA methylation-hydroxymethylation-demethylation pathway is only beginning to be explored. It is important to recognize that the current widely-used general sequencing methods (e.g. sequencing of bisulfite-converted DNA) do not distinguish DNA modified by 5mC from 5hmC [182]. Thus, the body of knowledge on DNA methylation, that has accumulated since the invention of the bisulfite-sequencing technique in 1992 [183], has to be considered as inexact. The development of new methods (e.g. [184]) is important in view of recent studies showing a significant amount of 5hmC in brain DNA [182,185–187], its increase during postnatal maturation [188], and dynamic changes in DNA 5hmC content regulated by neuronal activity [177].
5. Histone methylation
The methylation of histones at specific lysine and arginine residues is essential for the epigenetic regulation of transcription, cell division, and the formation of heterochromatin [189–191] because differentially-modified histones within nucleosomes of a genomic region constitute one of the central modes of regulation of transcriptional activity of the region [192]. The addition of methyl groups to histones exerts different effects depending on what residue is methylated and, for a lysine residue, how many methyl groups are added to the epsilon nitrogen atom. Typically, in the case of histone 3 (H3), dior trimethylation of lysine 9 (H3K9me2 and H3K9me3) and trimethylation of lysine 27 (H3K27me3) inhibit transcription [193–199]. In contrast, trimethylation of lysines 4, 36 or 79 (H3K4me3, H3K36me3 and H3K79me3, respectively) is associated with transcriptionally active genes as is monomethylation of histone 4 on lysine 20 and histone 2B on lysine 5 (H4K20me and H2BK5me) [195–202]. There are several enzymes that catalyze these modifications; all use AdoMet as the methyl donor [11]. For example, histone methyltransferase G9a (KMT1C, EHMT2) is responsible for di-methylation of H3K9 to H3K9me2 and for tri-methylation of H3K27 to H3K27me3 [149], and histone methyltransferase Suv39h1 (KMT1A) responsible for tri-methylation at H3K9 to H3K9me3 [203]. Therefore, in addition to modulating DNA methylation, choline supply could affect the methylation of amino acid residues on histone tails [204,205] leading to alterations in the expression of genes involved in growth and development. [Note dramatic changes in G9a and Prmt1 mRNA expression in the developing rat cortex, Figure 3]. Davison et al [206] examined several components of the histone 3 methylating machinery in rat liver and cerebral cortex on embryonic day 17 of rat fetuses derived from mothers consuming varying amounts of choline. The methylation of H3K9 and H3K27 and expression of G9a and Suv39h1 were directly related to the availability of choline. Similar results were obtained in a mouse model [168]. Consistent with the studies of Kovacheva et al [82] described above, DNA methylation of the G9a and Suv39h1 genes was dramatically upregulated by choline deficiency [206]. The latter finding points to the possibility that the expression of these histone methyltransferase is under negative control of methylation of their genes.
6. Conclusions and future directions
Choline nutrition during gestational period in rodents influences brain development and cognitive function throughout lifespan and high choline intake during gestation has emerged as a robust and reliable cognitive enhancing regimen. Moreover, high choline intake during the perinatal period is neuroprotective in a variety of animal models of neuronal damage, including that evoked by seizures [63,207–209], alcohol consumption [210–215] and genetic mutations [216–222]. Data showing that maternal choline supply during pregnancy modifies fetal histone and DNA methylation suggest that a concerted epigenomic mechanism contributes to the long-term developmental effects of varied choline intake in utero [82,159,168,206,223]. In addition to the central role of DNA and histone methylation in brain development, these processes are highly dynamic in adult brain and there is considerable evidence that they modulate the expression of key genes of synaptic plasticity [224–228] and are involved in mechanisms of learning and memory [229–232]. Thus, it is likely that choline nutrition would have modulatory effects on DNA and histone methylation in adult brain and that it could influence cognitive function in adulthood via an epigenetic mechanism as well. At present this idea remains to be tested experimentally. Importantly however, a recent study performed on 1391 normal adult and elderly people (average age 61 years) showed that verbal and visual memory function correlated positively with the amount of dietary choline consumption, with poorest performance in individuals with lowest choline intake and best performance in those who were consuming the highest amounts of choline [233]. These data further support the notion that adequate choline nutrition is essential for the maintenance of cognitive function in people.
Acknowledgments
Some of the studies reviewed here were supported by NIH grants AG009525 and CA120488 to JKB
Abbreviations
- 5caC
5-carboxylcytosine
- 5mC
5-methylcytosine
- 5hmC
5-hydroxymethylcytosine
- 5fC
5-formylcytosine
- ACh
acetylcholine
- AdoHcy
S-adenosylhomocysteine
- AdoMet
S-adenosylmethionine
- AHCY
AdoHcy hydrolase
- AI
adequate intake
- BDNF
brain-derived neurotrophic factor
- BHMT
betaine: homocysteine S-methyltransferase
- CaMKI
calcium/calmodulin-dependent protein kinase I
- CaMKIIβ
calcium/calmodulin-dependent protein kinase II beta
- CHAT
choline acetyltransferase
- CHDH
choline dehydrogenase
- CHKA
choline kinase A
- CREB
cAMP response element binding protein
- DMR
differentially methylated region
- DNMT
DNA methyltransferases
- FNB
Food and Nutrition Board
- G9a
(EHMT2, KMT1C) euchromatic histone lysine N-methyltransferase 2
- GABA
γ-aminobutyric acid
- H3
histone 3
- IGF
insulin-like growth factor
- LTP
termed long-term potentiation
- MAPK
mitogen-activated protein kinase
- MAT
methionine adenosyltransferase
- MECP2
methyl-CpG-binding protein 2
- MBD
methyl-CpG-binding domain protein
- MTHFD1
5,10-methylenetetrahydrofolate dehydrogenase 1
- MTHFR
methyltetrahydrofolate reductase
- MTR
5-methyltetrahydrofolate-homocysteine S-methyltransferase
- NGF
nerve growth factor
- NMDA
N-methyl-D-aspartate
- PCYT1, CTP
phosphocholine cytidylyltransferase 1
- PEMT
phosphatidylethanolamine N-methyltransferase
- PKCβ2
protein kinase C beta-2
- PRMT
protein arginine N-methyltransferase
- Suv39h1
(KMT1A) suppressor of variegation 3–9 homolog 1
- TET
Ten-eleven translocation
- USDA
United States Department of Agriculture
- VEGF
vascular endothelial growth factor
- Zif268 (EGR1)
early growth response
References
- 1.Strecker A. Über einige neue bestandheile der schweingalle. Ann Chem Pharmacie. 1862;123:353–360. [Google Scholar]
- 2.Baeyer A. Synthese des Neurins. Justus Liebigs Annalen der Chemie. 1866;140 [Google Scholar]
- 3.Loewi O. Nobel lecture. Nobelprize.org. 1936 http://www.nobelprize.org/nobel_prizes/medicine/laureates/1936/loewi-lecture.html (Accessed August 15, 2011)
- 4.Dale H. Nobel lecture. Nobelprize.org. 1936 http://www.nobelprize.org/nobel_prizes/medicine/laureates/1936/dale-lecture.html (Accessed August 15, 2011)
- 5.Mann PJ, Quastel JH. The oxidation of choline by rat liver. Biochem J. 1937;31:869–78. doi: 10.1042/bj0310869. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 6.Du Vigneaud V, Simmonds S, Chandler JP, Cohn M. A further investigation of the role of betaine in transmethylation reactions in vivo. J Biol Chem. 1946;165:639–48. [PubMed] [Google Scholar]
- 7.Cantoni GL. S-Adenosylmethionine; a new intermediate formed enzymatically from L-methionine and adenosinetriphosphate. J Biol Chem. 1953;204:403–16. [PubMed] [Google Scholar]
- 8.Chiang PK, Gordon RK, Tal J, Zeng GC, Doctor BP, Pardhasaradhi K, McCann PP. S-adenosylmethionine and methylation. FASEB J. 1996;10:471–480. [PubMed] [Google Scholar]
- 9.Grillo MA, Colombatto S. S-adenosylmethionine and its products. Amino Acids. 2008;34:187–93. doi: 10.1007/s00726-007-0500-9. [DOI] [PubMed] [Google Scholar]
- 10.Jeltsch A. Molecular enzymology of mammalian DNA methyltransferases. Curr Top Microbiol Immunol. 2006;301:203–25. doi: 10.1007/3-540-31390-7_7. [DOI] [PubMed] [Google Scholar]
- 11.Teperino R, Schoonjans K, Auwerx J. Histone methyl transferases and demethylases; can they link metabolism and transcription? Cell Metab. 2010;12:321–7. doi: 10.1016/j.cmet.2010.09.004. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 12.Allan FN, Bowie DJ, Macleod JJR, Robinson WL. Behavior of depancreatized dogs kept alive with insulin. Br J Exp Pathol. 1924;5:75–83. [Google Scholar]
- 13.Hershey JM. Substitution of “lecithin” for raw pancreas in the diet of the depancreatized dog. Am J Physiol. 1930;93:657–658. [Google Scholar]
- 14.Best CH, Huntsman ME. The effects of the components of lecithine upon deposition of fat in the liver. J Physiol. 1932;75:405–12. doi: 10.1113/jphysiol.1932.sp002899. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 15.Best CH, Channon HJ, Ridout JH. Choline and the dietary production of fatty livers. J Physiol. 1934;81:409–21. doi: 10.1113/jphysiol.1934.sp003144. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 16.Zeisel SH, Da Costa KA, Franklin PD, Alexander EA, Lamont JT, Sheard NF, Beiser A. Choline, an essential nutrient for humans. FASEB J. 1991;5:2093–2098. [PubMed] [Google Scholar]
- 17.FNB. Dietary reference intakes for thiamin, riboflavin, niacin, vitamin B6, folate, vitamin B12, panthotenic acid, biotin, and choline. National Academy Press; Washington, DC: 1998. [PubMed] [Google Scholar]
- 18.Garner SC, Mar MH, Zeisel SH. Choline distribution and metabolism in pregnant rats and fetuses are influenced by the choline content of the maternal diet. J Nutr. 1995;125:2851–2858. doi: 10.1093/jn/125.11.2851. [DOI] [PubMed] [Google Scholar]
- 19.Holmes-McNary MQ, Cheng WL, Mar MH, Fussell S, Zeisel SH. Choline and choline esters in human and rat milk and in infant formulas. Am J Clin Nutr. 1996;64:572–576. doi: 10.1093/ajcn/64.4.572. [DOI] [PubMed] [Google Scholar]
- 20.Zeisel SH, Char D, Sheard NF. Choline, phosphatidylcholine and sphingomyelin in human and bovine milk and infant formulas. J Nutr. 1986;116:50–58. doi: 10.1093/jn/116.1.50. [DOI] [PubMed] [Google Scholar]
- 21.da Costa KA, Badea M, Fischer LM, Zeisel SH. Elevated serum creatine phosphokinase in choline-deficient humans: mechanistic studies in C2C12 mouse myoblasts. Am J Clin Nutr. 2004;80:163–70. doi: 10.1093/ajcn/80.1.163. [DOI] [PubMed] [Google Scholar]
- 22.da Costa KA, Niculescu MD, Craciunescu CN, Fischer LM, Zeisel SH. Choline deficiency increases lymphocyte apoptosis and DNA damage in humans. Am J Clin Nutr. 2006;84:88–94. doi: 10.1093/ajcn/84.1.88. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 23.Jensen HH, Batres-Marquez SP, Carriquiry A, Schalinske KL. Choline in the diets of the US population: NHANES, 2003–2004. FASEB J. 2007;21:LB219. [Google Scholar]
- 24.Fischer LM, daCosta KA, Kwock L, Stewart PW, Lu TS, Stabler SP, Allen RH, Zeisel SH. Sex and menopausal status influence human dietary requirements for the nutrient choline. Am J Clin Nutr. 2007;85:1275–85. doi: 10.1093/ajcn/85.5.1275. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 25.Waite KA, Cabilio NR, Vance DE. Choline deficiency-induced liver damage is reversible in Pemt−/− mice. J Nutr. 2002;132:68–71. doi: 10.1093/jn/132.1.68. [DOI] [PubMed] [Google Scholar]
- 26.Walkey CJ, Yu L, Agellon LB, Vance DE. Biochemical and evolutionary significance of phospholipid methylation. J Biol Chem. 1998;273:27043–6. doi: 10.1074/jbc.273.42.27043. [DOI] [PubMed] [Google Scholar]
- 27.Zhu XN, Song JN, Mar MH, Edwards LJ, Zeisel SH. Phosphatidylethanolamine N-methyltransferase (PEMT) knockout mice have hepatic steatosis and abnormal hepatic choline metabolite concentrations despite ingesting a recommended dietary intake of choline. Biochem J. 2003;370:987–993. doi: 10.1042/BJ20021523. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 28.Resseguie M, Song J, Niculescu MD, da Costa KA, Randall TA, Zeisel SH. Phosphatidylethanolamine N-methyltransferase (PEMT) gene expression is induced by estrogen in human and mouse primary hepatocytes. FASEB J. 2007;21:2622–32. doi: 10.1096/fj.07-8227com. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 29.da Costa KA, Kozyreva OG, Song J, Galanko JA, Fischer LM, Zeisel SH. Common genetic polymorphisms affect the human requirement for the nutrient choline. FASEB J. 2006;20:1336–44. doi: 10.1096/fj.06-5734com. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 30.Caudill MA, Dellschaft N, Solis C, Hinkis S, Ivanov AA, Nash-Barboza S, Randall KE, Jackson B, Solomita GN, Vermeylen F. Choline intake, plasma riboflavin, and the phosphatidylethanolamine N-methyltransferase G5465A genotype predict plasma homocysteine in folate-deplete Mexican-American men with the methylenetetrahydrofolate reductase 677TT genotype. J Nutr. 2009;139:727–33. doi: 10.3945/jn.108.100222. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 31.Resseguie ME, da Costa KA, Galanko JA, Patel M, Davis IJ, Zeisel SH. Aberrant estrogen regulation of PEMT results in choline deficiency-associated liver dysfunction. J Biol Chem. 2011;286:1649–58. doi: 10.1074/jbc.M110.106922. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 32.Fischer LM, da Costa KA, Kwock L, Galanko J, Zeisel SH. Dietary choline requirements of women: effects of estrogen and genetic variation. Am J Clin Nutr. 2010;92:1113–9. doi: 10.3945/ajcn.2010.30064. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 33.Kohlmeier M, da Costa KA, Fischer LM, Zeisel SH. Genetic variation of folate-mediated one-carbon transfer pathway predicts susceptibility to choline deficiency in humans. Proc Natl Acad Sci U S A. 2005;102:16025–30. doi: 10.1073/pnas.0504285102. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 34.Holm PI, Hustad S, Ueland PM, Vollset SE, Grotmol T, Schneede J. Modulation of the homocysteine-betaine relationship by methylenetetrahydrofolate reductase 677 C->T genotypes and B-vitamin status in a large scale epidemiological study. J Clin Endocrinol Metab. 2007;92:1535–1541. doi: 10.1210/jc.2006-1471. [DOI] [PubMed] [Google Scholar]
- 35.Abratte CM, Wang W, Li R, Moriarty DJ, Caudill MA. Folate intake and the MTHFR C677T genotype influence choline status in young Mexican American women. J Nutr Biochem. 2008;19:158–65. doi: 10.1016/j.jnutbio.2007.02.004. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 36.Yan J, Wang W, Gregory JF, 3rd, Malysheva O, Brenna JT, Stabler SP, Allen RH, Caudill MA. MTHFR C677T genotype influences the isotopic enrichment of one-carbon metabolites in folate-compromised men consuming d9-choline. Am J Clin Nutr. 2011;93:348–55. doi: 10.3945/ajcn.110.005975. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 37.Sha W, da Costa KA, Fischer LM, Milburn MV, Lawton KA, Berger A, Jia W, Zeisel SH. Metabolomic profiling can predict which humans will develop liver dysfunction when deprived of dietary choline. FASEB J. 2010;24:2962–2975. doi: 10.1096/fj.09-154054. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 38.Patterson KY, Bhagwat AS, Williams JR, Howe JC, Holden JM, Zeisel SH, Da Costa CA, Mar H. USDA database for the choline content of common foods. Release two. 2008 http://www.ars.usda.gov/Services/docs.htm?docid=6232 (Accessed August 15, 2011)
- 39.Shaw GM, Carmichael SL, Laurent C, Rasmussen SA. Maternal nutrient intakes and risk of orofacial clefts. Epidemiology. 2006;17:285–91. doi: 10.1097/01.ede.0000208348.30012.35. [DOI] [PubMed] [Google Scholar]
- 40.Mostowska A, Hozyasz KK, Wojcicki P, Dziegelewska M, Jagodzinski PP. Associations of folate and choline metabolism gene polymorphisms with orofacial clefts. J Med Genet. 2009;47:809–815. doi: 10.1136/jmg.2009.070029. [DOI] [PubMed] [Google Scholar]
- 41.Xu X, Gammon MD, Zeisel SH, Lee YL, Wetmur JG, Teitelbaum SL, Bradshaw PT, Neugut AI, Santella RM, Chen J. Choline metabolism and risk of breast cancer in a population-based study. FASEB J. 2008;22:2045–2052. doi: 10.1096/fj.07-101279. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 42.Cho E, Willett WC, Colditz GA, Fuchs CS, Wu K, Chan AT, Zeisel SH, Giovannucci EL. Dietary choline and betaine and the risk of distal colorectal adenoma in women. J Natl Cancer Inst. 2007;99:1224–31. doi: 10.1093/jnci/djm082. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 43.Ibiebele TI, Hughes MC, Pandeya N, Zhao Z, Montgomery G, Hayward N, Green AC, Whiteman DC, Webb PM. High intake of folate from food sources is associated with reduced risk of esophageal cancer in an Australian population. J Nutr. 2011;141:274–83. doi: 10.3945/jn.110.131235. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 44.Shaw GM, Carmichael SL, Yang W, Selvin S, Schaffer DM. Periconceptional dietary intake of choline and betaine and neural tube defects in offspring. Am J Epidemiol. 2004;160:102–9. doi: 10.1093/aje/kwh187. [DOI] [PubMed] [Google Scholar]
- 45.Shaw GM, Finnell RH, Blom HJ, Carmichael SL, Vollset SE, Yang W, Ueland PM. Choline and Risk of Neural Tube Defects in a Folate-Fortified Population. Epidemiology. 2009;20:714–9. doi: 10.1097/EDE.0b013e3181ac9fe7. [DOI] [PubMed] [Google Scholar]
- 46.Enaw JO, Zhu H, Yang W, Lu W, Shaw GM, Lammer EJ, Finnell RH. CHKA and PCYT1A gene polymorphisms, choline intake and spina bifida risk in a California population. BMC Med. 2006;4:36. doi: 10.1186/1741-7015-4-36. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 47.Albright CD, Friedrich CB, Brown EC, Mar MH, Zeisel SH. Maternal dietary choline availability alters mitosis, apoptosis and the localization of TOAD-64 protein in the developing fetal rat septum. Dev Brain Res. 1999;115:123–129. doi: 10.1016/s0165-3806(99)00057-7. [DOI] [PubMed] [Google Scholar]
- 48.Albright CD, Tsai AY, Friedrich CB, Mar MH, Zeisel SH. Choline availability alters embryonic development of the hippocampus and septum in the rat. Dev Brain Res. 1999;113:13–20. doi: 10.1016/s0165-3806(98)00183-7. [DOI] [PubMed] [Google Scholar]
- 49.Craciunescu CN, Albright CD, Mar MH, Song J, Zeisel SH. Choline availability during embryonic development alters progenitor cell mitosis in developing mouse hippocampus. J Nutr. 2003;133:3614–3618. doi: 10.1093/jn/133.11.3614. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 50.Meck WH, Williams CL. Simultaneous temporal processing is sensitive to prenatal choline availability in mature and aged rats. Neuroreport. 1997;8:3045–3051. doi: 10.1097/00001756-199709290-00009. [DOI] [PubMed] [Google Scholar]
- 51.Mellott TJ, Williams CL, Meck WH, Blusztajn JK. Prenatal choline supplementation advances hippocampal development and enhances MAPK and CREB activation. FASEB J. 2004;18:NIL412–NIL427. doi: 10.1096/fj.03-0877fje. [DOI] [PubMed] [Google Scholar]
- 52.Meck WH, Smith RA, Williams CL. Pre- and postnatal choline supplementation produces long-term facilitation of spatial memory. Dev Psychobiol. 1988;21:339–353. doi: 10.1002/dev.420210405. [DOI] [PubMed] [Google Scholar]
- 53.Meck WH, Smith RA, Williams CL. Organizational changes in cholinergic activity and enhanced visuospatial memory as a function of choline administered prenatally or postnatally or both. Behav Neurosci. 1989;103:1234–1241. doi: 10.1037//0735-7044.103.6.1234. [DOI] [PubMed] [Google Scholar]
- 54.Meck WH, Williams CL. Characterization of the facilitative effects of perinatal choline supplementation on timing and temporal memory. Neuroreport. 1997;8:2831–2835. doi: 10.1097/00001756-199709080-00005. [DOI] [PubMed] [Google Scholar]
- 55.Meck WH, Williams CL. Perinatal choline supplementation increases the threshold for chunking in spatial memory. Neuroreport. 1997;8:3053–3059. doi: 10.1097/00001756-199709290-00010. [DOI] [PubMed] [Google Scholar]
- 56.Meck WH, Williams CL. Choline supplementation during prenatal development reduces proactive interference in spatial memory. Dev Brain Res. 1999;118:51–9. doi: 10.1016/s0165-3806(99)00105-4. [DOI] [PubMed] [Google Scholar]
- 57.Meck WH, Williams CL, Cermak JM, Blusztajn JK. Developmental periods of choline sensitivity provide an ontogenetic mechanism for regulating memory capacity and age-related dementia. Front Integr Neurosci. 2008;1:7. doi: 10.3389/neuro.07.007.2007. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 58.Clelland CD, Choi M, Romberg C, Clemenson GD, Jr, Fragniere A, Tyers P, Jessberger S, Saksida LM, Barker RA, Gage FH, Bussey TJ. A functional role for adult hippocampal neurogenesis in spatial pattern separation. Science. 2009;325:210–3. doi: 10.1126/science.1173215. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 59.Kitamura T, Saitoh Y, Takashima N, Murayama A, Niibori Y, Ageta H, Sekiguchi M, Sugiyama H, Inokuchi K. Adult neurogenesis modulates the hippocampus-dependent period of associative fear memory. Cell. 2009;139:814–27. doi: 10.1016/j.cell.2009.10.020. [DOI] [PubMed] [Google Scholar]
- 60.Glenn MJ, Gibson EM, Kirby ED, Mellott TJ, Blusztajn JK, Williams CL. Prenatal choline availability modulates hippocampal neurogenesis and neurogenic responses to enriching experiences in adult female rats. Eur J Neurosci. 2007;25:2473–2482. doi: 10.1111/j.1460-9568.2007.05505.x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 61.Glenn MJ, Kirby ED, Gibson EM, Wong-Goodrich SJ, Mellott TJ, Blusztajn JK, Williams CL. Age-related declines in exploratory behavior and markers of hippocampal plasticity are attenuated by prenatal choline supplementation in rats. Brain Res. 2008;1237:110–23. doi: 10.1016/j.brainres.2008.08.049. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 62.Wong-Goodrich SJ, Glenn MJ, Mellott TJ, Blusztajn JK, Meck WH, Williams CL. Spatial memory and hippocampal plasticity are differentially sensitive to the availability of choline in adulthood as a function of choline supply in utero. Brain Res. 2008;1237:153–66. doi: 10.1016/j.brainres.2008.08.074. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 63.Wong-Goodrich SJ, Mellott TJ, Glenn MJ, Blusztajn JK, Williams CL. Prenatal choline supplementation attenuates neuropathological response to status epilepticus in the adult rat hippocampus. Neurobiol Dis. 2008;30:255–69. doi: 10.1016/j.nbd.2008.01.008. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 64.Napoli I, Blusztajn JK, Mellott TJ. Prenatal choline supplementation in rats increases the expression of IGF2 and its receptor IGF2R and enhances IGF2-induced acetylcholine release in hippocampus and frontal cortex. Brain Res. 2008;1237:124–35. doi: 10.1016/j.brainres.2008.08.046. [DOI] [PubMed] [Google Scholar]
- 65.Williams CL, Meck WH, Heyer D, Loy R. Hypertrophy of basal forebrain neurons and enhanced visuospatial memory in perinatally choline-supplemented rats. Brain Res. 1998;794:225–238. doi: 10.1016/s0006-8993(98)00229-7. [DOI] [PubMed] [Google Scholar]
- 66.Fibiger HC. Cholinergic mechanisms in learning, memory and dementia: A review of recent evidence. TINS. 1991;14:220–223. doi: 10.1016/0166-2236(91)90117-d. [DOI] [PubMed] [Google Scholar]
- 67.Sarter M, Parikh V. Choline transporters, cholinergic transmission and cognition. Nat Rev Neurosci. 2005;6:48–56. doi: 10.1038/nrn1588. [DOI] [PubMed] [Google Scholar]
- 68.Cermak JM, Holler T, Jackson DA, Blusztajn JK. Prenatal availability of choline modifies development of the hippocampal cholinergic system. FASEB J. 1998;12:349–357. doi: 10.1096/fasebj.12.3.349. [DOI] [PubMed] [Google Scholar]
- 69.Sweatt JD. The neuronal MAP kinase cascade: a biochemical signal integration system subserving synaptic plasticity and memory. J Neurochem. 2001;76:1–10. doi: 10.1046/j.1471-4159.2001.00054.x. [DOI] [PubMed] [Google Scholar]
- 70.Jones JP, H Meck W, Williams CL, Wilson WA, Swartzwelder HS. Choline availability to the developing rat fetus alters adult hippocampal long-term potentiation. Dev Brain Res. 1999;118:159–67. doi: 10.1016/s0165-3806(99)00103-0. [DOI] [PubMed] [Google Scholar]
- 71.Pyapali GK, Turner DA, Williams CL, Meck WH, Swartzwelder HS. Prenatal dietary choline supplementation decreases the threshold for induction of long-term potentiation in young adult rats. J Neurophysiol. 1998;79:1790–1796. doi: 10.1152/jn.1998.79.4.1790. [DOI] [PubMed] [Google Scholar]
- 72.Montoya D, Swartzwelder HS. Prenatal choline supplementation alters hippocampal N-methyl-D-aspartate receptor-mediated neurotransmission in adult rats. Neurosci Lett. 2000;296:85–88. doi: 10.1016/s0304-3940(00)01660-8. [DOI] [PubMed] [Google Scholar]
- 73.Mellott TJ, Follettie MT, Diesl V, Hill AA, Lopez-Coviella I, Blusztajn JK. Prenatal choline availability modulates hippocampal and cerebral cortical gene expression. FASEB J. 2007;21:1311–1323. doi: 10.1096/fj.06-6597com. [DOI] [PubMed] [Google Scholar]
- 74.Chen ZX, Riggs AD. DNA methylation and demethylation in mammals. J Biol Chem. 2011;286:18347–53. doi: 10.1074/jbc.R110.205286. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 75.Hoffman DR, Haning JA, Cornatzer WE. Effect of a methyl-deficient diet on rat liver phosphatidylcholine biosyntheis. Can J Biochem. 1981;59:543–550. doi: 10.1139/o81-075. [DOI] [PubMed] [Google Scholar]
- 76.Shivapurkar N, Poirier LA. Tissue levels of S-adenosylmethionine and S-adenosylhomocysteine in rats fed methyl-deficient, amino acid-defined diets for one to five weeks. Carcinogenesis. 1983;4:1051–7. doi: 10.1093/carcin/4.8.1051. [DOI] [PubMed] [Google Scholar]
- 77.Shivapurkar N, Wilson MJ, Hoover KL, Mikol YB, Creasia D, Poirier LA. Hepatic DNA methylation and liver tumor formation in male C3H mice fed methionine- and choline-deficient diets. J Natl Cancer Inst. 1986;77:213–7. [PubMed] [Google Scholar]
- 78.Svardal AM, Ueland PM, Berge RK, Aarsland A, Aarsaether N, Lonning PE, Refsum H. Effect of methotrexate on homocysteine and other sulfur compounds in tissues of rats fed a normal or a defined, choline-deficient diet. Cancer Chemother Pharmacol. 1988;21:313–8. doi: 10.1007/BF00264197. [DOI] [PubMed] [Google Scholar]
- 79.Zeisel SH, Zola T, DaCosta K-A, Pomfret EA. Effect of choline deficiency on S-adenosylmethionine and methionine concentrations in rat liver. Biochem J. 1989;259:725–729. doi: 10.1042/bj2590725. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 80.Pomfret EA, da Costa K, Zeisel SH. Effects of choline deficiency and methotrexate treatment upon rat liver. J Nutr Biochem. 1990;1:533–541. doi: 10.1016/0955-2863(90)90039-n. [DOI] [PubMed] [Google Scholar]
- 81.Innis SM, Davidson AG, Melynk S, James SJ. Choline-related supplements improve abnormal plasma methionine-homocysteine metabolites and glutathione status in children with cystic fibrosis. Am J Clin Nutr. 2007;85:702–8. doi: 10.1093/ajcn/85.3.702. [DOI] [PubMed] [Google Scholar]
- 82.Kovacheva VP, Mellott TJ, Davison JM, Wagner N, Lopez-Coviella I, Schnitzler AC, Blusztajn JK. Gestational choline deficiency causes global and Igf2 gene DNA hypermethylation by up-regulation of Dnmt1 expression. J Biol Chem. 2007;282:31777–88. doi: 10.1074/jbc.M705539200. [DOI] [PubMed] [Google Scholar]
- 83.Wolff GL, Kodell RL, Moore SR, Cooney CA. Maternal epigenetics and methyl supplements affect agouti gene expression in Avy/a mice. FASEB J. 1998;12:949–957. [PubMed] [Google Scholar]
- 84.Cooney CA, Dave AA, Wolff GL. Maternal methyl supplements in mice affect epigenetic variation and DNA methylation of offspring. J Nutr. 2002;132:2393S–2400S. doi: 10.1093/jn/132.8.2393S. [DOI] [PubMed] [Google Scholar]
- 85.Waterland RA, Jirtle RL. Transposable elements: targets for early nutritional effects on epigenetic gene regulation. Mol Cell Biol. 2003;23:5293–300. doi: 10.1128/MCB.23.15.5293-5300.2003. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 86.Waterland RA, Dolinoy DC, Lin JR, Smith CA, Shi X, Tahiliani KG. Maternal methyl supplements increase offspring DNA methylation at Axin Fused. Genesis. 2006;44:401–6. doi: 10.1002/dvg.20230. [DOI] [PubMed] [Google Scholar]
- 87.Dolinoy DC, Huang D, Jirtle RL. Maternal nutrient supplementation counteracts bisphenol A-induced DNA hypomethylation in early development. Proc Natl Acad Sci U S A. 2007;104:13056–61. doi: 10.1073/pnas.0703739104. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 88.Waterland RA, Travisano M, Tahiliani KG, Rached MT, Mirza S. Methyl donor supplementation prevents transgenerational amplification of obesity. Int J Obes (Lond) 2008:1373–1379. doi: 10.1038/ijo.2008.100. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 89.Cropley JE, Suter CM, Beckman KB, Martin DI. Germ-line epigenetic modification of the murine A vy allele by nutritional supplementation. Proc Natl Acad Sci U S A. 2006;103:17308–12. doi: 10.1073/pnas.0607090103. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 90.Copeland DH, Salmon WD. The Occurrence of Neoplasms in the Liver, Lungs, and Other Tissues of Rats as a Result of Prolonged Choline Deficiency. Am J Pathol. 1946;22:1059–79. [PubMed] [Google Scholar]
- 91.Engel RW, Copeland DH, Salmon WD. Carcinogenic effects associated with diets deficient in choline and related nutrients. Ann N Y Acad Sci. 1947;49:49–67. [Google Scholar]
- 92.Salmon WD, Copeland DH. Liver carcinoma and related lesions in chronic choline deficiency. Ann N Y Acad Sci. 1954;57:665–77. doi: 10.1111/j.1749-6632.1954.tb36443.x. [DOI] [PubMed] [Google Scholar]
- 93.Shinozuka H, Katyal SL, Perera MI. Choline deficiency and chemical carcinogenesis. Adv Exp Med Biol. 1986;206:253–67. doi: 10.1007/978-1-4613-1835-4_19. [DOI] [PubMed] [Google Scholar]
- 94.Lombardi B, Chandar N, Locker J. Nutritional model of hepatocarcinogenesis. Rats fed choline-devoid diet. Dig Dis Sci. 1991;36:979–984. doi: 10.1007/BF01297151. [DOI] [PubMed] [Google Scholar]
- 95.Newberne PM, Rogers AE. Labile methyl groups and the promotion of cancer. Annu Rev Nutr. 1986;6:407–32. doi: 10.1146/annurev.nu.06.070186.002203. [DOI] [PubMed] [Google Scholar]
- 96.Newberne PM. Lipotropic factors and oncogenesis. Adv Exp Med Biol. 1986;206:223–51. doi: 10.1007/978-1-4613-1835-4_18. [DOI] [PubMed] [Google Scholar]
- 97.Feinberg AP, Vogelstein B. Hypomethylation distinguishes genes of some human cancers from their normal counterparts. Nature. 1983;301:89–92. doi: 10.1038/301089a0. [DOI] [PubMed] [Google Scholar]
- 98.Feinberg AP, Tycko B. The history of cancer epigenetics. Nat Rev Cancer. 2004;4:143–53. doi: 10.1038/nrc1279. [DOI] [PubMed] [Google Scholar]
- 99.Irizarry RA, Ladd-Acosta C, Wen B, Wu Z, Montano C, Onyango P, Cui H, Gabo K, Rongione M, Webster M, Ji H, Potash JB, Sabunciyan S, Feinberg AP. The human colon cancer methylome shows similar hypo- and hypermethylation at conserved tissue-specific CpG island shores. Nat Genet. 2009;41:178–86. doi: 10.1038/ng.298. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 100.Rodriguez-Paredes M, Esteller M. Cancer epigenetics reaches mainstream oncology. Nat Med. 2011;17:330–9. doi: 10.1038/nm.2305. [DOI] [PubMed] [Google Scholar]
- 101.Bhave MR, Wilson MJ, Poirier LA. c-H-ras and c-K-ras gene hypomethylation in the livers and hepatomas of rats fed methyl-deficient, amino acid-defined diets. Carcinogenesis. 1988;9:343–8. doi: 10.1093/carcin/9.3.343. [DOI] [PubMed] [Google Scholar]
- 102.Zapisek WF, Cronin GM, Lyn-Cook BD, Poirier LA. The onset of oncogene hypomethylation in the livers of rats fed methyl- deficient, amino acid-defined diets. Carcinogenesis. 1992;13:1869–1872. doi: 10.1093/carcin/13.10.1869. [DOI] [PubMed] [Google Scholar]
- 103.Dizik M, Christman JK, Wainfan E. Alterations in expression and methylation of specific genes in livers of rats fed a cancer promoting methyl-deficient diet. Carcinogenesis. 1991;12:1307–1312. doi: 10.1093/carcin/12.7.1307. [DOI] [PubMed] [Google Scholar]
- 104.Smith ML, Yeleswarapu L, Scalamogna P, Locker J, Lombardi B. p53 mutations in hepatocellular carcinomas induced by a choline-devoid diet in male Fischer 344 rats. Carcinogenesis. 1993;14:503–510. doi: 10.1093/carcin/14.3.503. [DOI] [PubMed] [Google Scholar]
- 105.Pogribny IP, Miller BJ, James SJ. Alterations in hepatic p53 gene methylation patterns during tumor progression with folate/methyl deficiency in the rat. Cancer Lett. 1997;115:31–8. doi: 10.1016/s0304-3835(97)04708-3. [DOI] [PubMed] [Google Scholar]
- 106.Tsujiuchi T, Tsutsumi M, Sasaki Y, Takahama M, Konishi Y. Hypomethylation of CpG sites and c-myc gene overexpression in hepatocellular carcinomas, but not hyperplastic nodules, induced by a choline-deficient L-amino acid-defined diet in rats. Jpn J Cancer Res. 1999;90:909–913. doi: 10.1111/j.1349-7006.1999.tb00834.x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 107.Pogribny IP, James SJ. De novo methylation of the p16INK4A gene in early preneoplastic liver and tumors induced by folate/methyl deficiency in rats. Cancer Lett. 2002;187:69–75. doi: 10.1016/s0304-3835(02)00408-1. [DOI] [PubMed] [Google Scholar]
- 108.James SJ, Pogribny IP, Pogribna M, Miller BJ, Jernigan S, Melnyk S. Mechanisms of DNA damage, DNA hypomethylation, and tumor progression in the folate/methyl-deficient rat model of hepatocarcinogenesis. J Nutr. 2003;133:3740S–3747S. doi: 10.1093/jn/133.11.3740S. [DOI] [PubMed] [Google Scholar]
- 109.Pogribny IP, James SJ, Jernigan S, Pogribna M. Genomic hypomethylation is specific for preneoplastic liver in folate/methyl deficient rats and does not occur in non-target tissues. Mutat Res. 2004;548:53–9. doi: 10.1016/j.mrfmmm.2003.12.014. [DOI] [PubMed] [Google Scholar]
- 110.Pogribny IP, Ross SA, Wise C, Pogribna M, Jones EA, Tryndyak VP, James SJ, Dragan YP, Poirier LA. Irreversible global DNA hypomethylation as a key step in hepatocarcinogenesis induced by dietary methyl deficiency. Mutat Res. 2006;593:80–7. doi: 10.1016/j.mrfmmm.2005.06.028. [DOI] [PubMed] [Google Scholar]
- 111.Asada K, Kotake Y, Asada R, Saunders D, Broyles RH, Towner RA, Fukui H, Floyd RA. LINE-1 hypomethylation in a choline-deficiency-induced liver cancer in rats: dependence on feeding period. J Biomed Biotechnol. 2006;2006:17142. doi: 10.1155/JBB/2006/17142. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 112.Tsujiuchi T, Shimizu K, Itsuzaki Y, Onishi M, Sugata E, Fujii H, Honoki K. CpG site hypermethylation of E-cadherin and Connexin26 genes in hepatocellular carcinomas induced by a choline-deficient L-Amino Acid-defined diet in rats. Mol Carcinog. 2007;46:269–274. doi: 10.1002/mc.20268. [DOI] [PubMed] [Google Scholar]
- 113.Shimizu K, Onishi M, Sugata E, Sokuza Y, Mori C, Nishikawa T, Honoki K, Tsujiuchi T. Disturbance of DNA methylation patterns in the early phase of hepatocarcinogenesis induced by a choline-deficient L-amino acid-defined diet in rats. Cancer Sci. 2007;98:1318–22. doi: 10.1111/j.1349-7006.2007.00564.x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 114.Pogribny IP, Shpyleva SI, Muskhelishvili L, Bagnyukova TV, James SJ, Beland FA. Role of DNA damage and alterations in cytosine DNA methylation in rat liver carcinogenesis induced by a methyl-deficient diet. Mutat Res. 2009;669:56–62. doi: 10.1016/j.mrfmmm.2009.05.003. [DOI] [PubMed] [Google Scholar]
- 115.Robertson KD, Jones PA. DNA methylation: past, present and future directions. Carcinogenesis. 2000;21:461–467. doi: 10.1093/carcin/21.3.461. [DOI] [PubMed] [Google Scholar]
- 116.Bird A. DNA methylation patterns and epigenetic memory. Genes Dev. 2002;16:6–21. doi: 10.1101/gad.947102. [DOI] [PubMed] [Google Scholar]
- 117.Bernstein BE, Meissner A, Lander ES. The mammalian epigenome. Cell. 2007;128:669–81. doi: 10.1016/j.cell.2007.01.033. [DOI] [PubMed] [Google Scholar]
- 118.Meissner A. Epigenetic modifications in pluripotent and differentiated cells. Nat Biotechnol. 2010;28:1079–88. doi: 10.1038/nbt.1684. [DOI] [PubMed] [Google Scholar]
- 119.Portela A, Esteller M. Epigenetic modifications and human disease. Nat Biotechnol. 2010;28:1057–68. doi: 10.1038/nbt.1685. [DOI] [PubMed] [Google Scholar]
- 120.Waterland RA, Lin JR, Smith CA, Jirtle RL. Post-weaning diet affects genomic imprinting at the insulin-like growth factor 2 (Igf2) locus. Hum Mol Genet. 2006;15:705–16. doi: 10.1093/hmg/ddi484. [DOI] [PubMed] [Google Scholar]
- 121.Paulsen M, Ferguson-Smith AC. DNA methylation in genomic imprinting, development, and disease. J Pathol. 2001;195:97–110. doi: 10.1002/path.890. [DOI] [PubMed] [Google Scholar]
- 122.Li E. Chromatin modification and epigenetic reprogramming in mammalian development. Nat Rev Genet. 2002;3:662–73. doi: 10.1038/nrg887. [DOI] [PubMed] [Google Scholar]
- 123.Reinhart B, Eljanne M, Chaillet JR. Shared role for differentially methylated domains of imprinted genes. Mol Cell Biol. 2002;22:2089–2098. doi: 10.1128/MCB.22.7.2089-2098.2002. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 124.Tucker KL. Methylated cytosine and the brain: a new base for neuroscience. Neuron. 2001;30:649–652. doi: 10.1016/s0896-6273(01)00325-7. [DOI] [PubMed] [Google Scholar]
- 125.Robertson KD, Wolffe AP. DNA methylation in health and disease. Nat Rev Genet. 2000;1:11–19. doi: 10.1038/35049533. [DOI] [PubMed] [Google Scholar]
- 126.Nan X, Ng HH, Johnson CA, Laherty CD, Turner BM, Eisenman RN, Bird A. Transcriptional repression by the methyl-CpG-binding protein MeCP2 involves a histone deacetylase complex. Nature. 1998;393:386–9. doi: 10.1038/30764. [DOI] [PubMed] [Google Scholar]
- 127.Jones PL, Veenstra GJ, Wade PA, Vermaak D, Kass SU, Landsberger N, Strouboulis J, Wolffe AP. Methylated DNA and MeCP2 recruit histone deacetylase to repress transcription. Nat Genet. 1998;19:187–91. doi: 10.1038/561. [DOI] [PubMed] [Google Scholar]
- 128.Eden S, Hashimshony T, Keshet I, Cedar H, Thorne AW. DNA methylation models histone acetylation. Nature. 1998;394:842. doi: 10.1038/29680. [DOI] [PubMed] [Google Scholar]
- 129.Takizawa T, Nakashima K, Namihira M, Ochiai W, Uemura A, Yanagisawa M, Fujita N, Nakao M, Taga T. DNA methylation is a critical cell-intrinsic determinant of astrocyte differentiation in the fetal brain. Dev Cell. 2001;1:749–758. doi: 10.1016/s1534-5807(01)00101-0. [DOI] [PubMed] [Google Scholar]
- 130.Klose RJ, Bird AP. Genomic DNA methylation: the mark and its mediators. Trends Biochem Sci. 2006;31:89–97. doi: 10.1016/j.tibs.2005.12.008. [DOI] [PubMed] [Google Scholar]
- 131.Barrett RM, Wood MA. Beyond transcription factors: the role of chromatin modifying enzymes in regulating transcription required for memory. Learn Mem. 2008;15:460–7. doi: 10.1101/lm.917508. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 132.Klempnauer KH. Methylation-sensitive DNA binding by v-myb and c-myb proteins. Oncogene. 1993;8:111–115. [PubMed] [Google Scholar]
- 133.Campanero MR, Armstrong MI, Flemington EK. CpG methylation as a mechanism for the regulation of E2F activity. Proc Natl Acad Sci U S A. 2000;97:6481–6486. doi: 10.1073/pnas.100340697. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 134.Weih F, Nitsch D, Reik A, Schutz G, Becker PB. Analysis of CpG methylation and genomic footprinting at the tyrosine aminotransferase gene: DNA methylation alone is not sufficient to prevent protein binding in vivo. EMBO J. 1991;10:2559–2567. doi: 10.1002/j.1460-2075.1991.tb07796.x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 135.Comb M, Goodman HM. CpG methylation inhibits proenkephalin gene expression and binding of the transcription factor AP-2. Nucleic Acids Res. 1990;18:3975–3982. doi: 10.1093/nar/18.13.3975. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 136.Kirillov A, Kistler B, Mostoslavsky R, Cedar H, Wirth T, Bergman Y. A role for nuclear NF-kappaB in B-cell-specific demethylation of the Igkappa locus. Nat Genet. 1996;13:435–441. doi: 10.1038/ng0895-435. [DOI] [PubMed] [Google Scholar]
- 137.Eden S, Constancia M, Hashimshony T, Dean W, Goldstein B, Johnson AC, Keshet I, Reik W, Cedar H. An upstream repressor element plays a role in Igf2 imprinting. EMBO J. 2001;20:3518–3525. doi: 10.1093/emboj/20.13.3518. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 138.Murrell A, Heeson S, Bowden L, Constancia M, Dean W, Kelsey G, Reik W. An intragenic methylated region in the imprinted Igf2 gene augments transcription. EMBO Rep. 2001;2:1101–1106. doi: 10.1093/embo-reports/kve248. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 139.Inano K, Suetake I, Ueda T, Miyake Y, Nakamura M, Okada M, Tajima S. Maintenance-type DNA methyltransferase is highly expressed in post- mitotic neurons and localized in the cytoplasmic compartment. J Biochem (Tokyo) 2000;128:315–321. doi: 10.1093/oxfordjournals.jbchem.a022755. [DOI] [PubMed] [Google Scholar]
- 140.Trasler JM, Trasler DG, Bestor TH, Li E, Ghibu F. DNA methyltransferase in normal and Dnmtn/Dnmtn mouse embryos. Dev Dyn. 1996;206:239–247. doi: 10.1002/(SICI)1097-0177(199607)206:3<239::AID-AJA2>3.0.CO;2-J. [DOI] [PubMed] [Google Scholar]
- 141.Goto K, Numata M, Komura JI, Ono T, Bestor TH, Kondo H. Expression of DNA methyltransferase gene in mature and immature neurons as well as proliferating cells in mice. Differentiation. 1994;56:39–44. doi: 10.1046/j.1432-0436.1994.56120039.x. [DOI] [PubMed] [Google Scholar]
- 142.Brooks PJ, Marietta C, Goldman D. DNA mismatch repair and DNA methylation in adult brain neurons. J Neurosci. 1996;16:939–945. doi: 10.1523/JNEUROSCI.16-03-00939.1996. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 143.Tawa R, Ono T, Kurishita A, Okada S, Hirose S. Changes of DNA methylation level during pre- and postnatal periods in mice. Differentiation. 1990;45:44–48. doi: 10.1111/j.1432-0436.1990.tb00455.x. [DOI] [PubMed] [Google Scholar]
- 144.Wilson VL, Smith RA, Ma S, Cutler RG. Genomic 5-methyldeoxycytidine decreases with age. J Biol Chem. 1987;262:9948–9951. [PubMed] [Google Scholar]
- 145.Ono T, Uehara Y, Kurishita A, Tawa R, Sakurai H. Biological significance of DNA methylation in the ageing process. Age Ageing. 1993;22:S34–S43. doi: 10.1093/ageing/22.suppl_1.s34. [DOI] [PubMed] [Google Scholar]
- 146.Siegmund KD, Connor CM, Campan M, Long TI, Weisenberger DJ, Biniszkiewicz D, Jaenisch R, Laird PW, Akbarian S. DNA Methylation in the Human Cerebral Cortex Is Dynamically Regulated throughout the Life Span and Involves Differentiated Neurons. PLoS ONE. 2007;2:e895. doi: 10.1371/journal.pone.0000895. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 147.Hernandez DG, Nalls MA, Gibbs JR, Arepalli S, van der Brug M, Chong S, Moore M, Longo DL, Cookson MR, Traynor BJ, Singleton AB. Distinct DNA methylation changes highly correlated with chronological age in the human brain. Hum Mol Genet. 2011;20:1164–72. doi: 10.1093/hmg/ddq561. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 148.Iwamoto K, Bundo M, Ueda J, Oldham MC, Ukai W, Hashimoto E, Saito T, Geschwind DH, Kato T. Neurons show distinctive DNA methylation profile and higher interindividual variations compared with non-neurons. Genome Res. 2011;21:688–96. doi: 10.1101/gr.112755.110. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 149.Tachibana M, Sugimoto K, Nozaki M, Ueda J, Ohta T, Ohki M, Fukuda M, Takeda N, Niida H, Kato H, Shinkai Y. G9a histone methyltransferase plays a dominant role in euchromatic histone H3 lysine 9 methylation and is essential for early embryogenesis. Genes Dev. 2002;16:1779–91. doi: 10.1101/gad.989402. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 150.Nicholson TB, Chen T, Richard S. The physiological and pathophysiological role of PRMT1-mediated protein arginine methylation. Pharmacol Res. 2009;60:466–74. doi: 10.1016/j.phrs.2009.07.006. [DOI] [PubMed] [Google Scholar]
- 151.Feng J, Chang H, Li E, Fan G. Dynamic expression of de novo DNA methyltransferases Dnmt3a and Dnmt3b in the central nervous system. J Neurosci Res. 2005;79:734–46. doi: 10.1002/jnr.20404. [DOI] [PubMed] [Google Scholar]
- 152.Hsu DW, Lin MJ, Lee TL, Wen SC, Chen X, Shen CK. Two major forms of DNA (cytosine-5) methyltransferase in human somatic tissues. Proc Natl Acad Sci U S A. 1999;96:9751–6. doi: 10.1073/pnas.96.17.9751. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 153.Okano M, Li E. Genetic analyses of DNA methyltransferase genes in mouse model system. J Nutr. 2002;132:2462S–2465S. doi: 10.1093/jn/132.8.2462S. [DOI] [PubMed] [Google Scholar]
- 154.Singal R, vanWert JN. De novo methylation of an embryonic globin gene during normal development is strand specific and spreads from the proximal transcribed region. Blood. 2001;98:3441–3446. doi: 10.1182/blood.v98.12.3441. [DOI] [PubMed] [Google Scholar]
- 155.Alonso-Aperte E, Varela-Moreiras G. Brain folates and DNA methylation in rats fed a choline deficient diet or treated with low doses of methotrexate. Int J Vitam Nutr Res. 1996;66:232–236. [PubMed] [Google Scholar]
- 156.Wainfan E, Dizik M, Stender M, Christman JK. Rapid appearance of hypomethylated DNA in livers of rats fed cancer- promoting, methyl-deficient diets. Cancer Res. 1989;49:4094–4097. [PubMed] [Google Scholar]
- 157.Wilson MJ, Shivapurkar N, Poirier LA. Hypomethylation of hepatic nuclear DNA in rats fed with a carcinogenic methyl-deficient diet. Biochem J. 1984;218:987–90. doi: 10.1042/bj2180987. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 158.Christman JK, Sheikhnejad G, Dizik M, Abileah S, Wainfan E. Reversibility of changes in nucleic acid methylation and gene expression induced in rat liver by severe dietary methyl deficiency. Carcinogenesis. 1993;14:551–557. doi: 10.1093/carcin/14.4.551. [DOI] [PubMed] [Google Scholar]
- 159.Niculescu MD, Craciunescu CN, Zeisel SH. Dietary choline deficiency alters global and gene-specific DNA methylation in the developing hippocampus of mouse fetal brains. FASEB J. 2006;20:43–9. doi: 10.1096/fj.05-4707com. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 160.Kim YI, Pogribny IP, Basnakian AG, Miller JW, Selhub J, James SJ, Mason JB. Folate deficiency in rats induces DNA strand breaks and hypomethylation within the p53 tumor suppressor gene. Am J Clin Nutr. 1997;65:46–52. doi: 10.1093/ajcn/65.1.46. [DOI] [PubMed] [Google Scholar]
- 161.Niculescu MD, Yamamuro Y, Zeisel SH. Choline availability modulates human neuroblastoma cell proliferation and alters the methylation of the promoter region of the cyclin-dependent kinase inhibitor 3 gene. J Neurochem. 2004;89:1252–9. doi: 10.1111/j.1471-4159.2004.02414.x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 162.Mehedint MG, Craciunescu CN, Zeisel SH. Maternal dietary choline deficiency decreases angiogenesis in fetal mouse hippocampus. Proc Natl Acad Sci U S A. 2010;107:12834–12839. doi: 10.1073/pnas.0914328107. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 163.Shin W, Yan J, Abratte CM, Vermeylen F, Caudill MA. Choline intake exceeding current dietary recommendations preserves markers of cellular methylation in a genetic subgroup of folate-compromised men. J Nutr. 2010;140:975–80. doi: 10.3945/jn.110.121186. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 164.Kim YI, Christman JK, Fleet JC, Cravo ML, Salomon RN, Smith D, Ordovas J, Selhub J, Mason JB. Moderate folate deficiency does not cause global hypomethylation of hepatic and colonic DNA or c-myc-specific hypomethylation of colonic DNA in rats. Am J Clin Nutr. 1995;61:1083–90. doi: 10.1093/ajcn/61.4.1083. [DOI] [PubMed] [Google Scholar]
- 165.Song J, Sohn KJ, Medline A, Ash C, Gallinger S, Kim YI. Chemopreventive effects of dietary folate on intestinal polyps in Apc+/−Msh2−/− mice. Cancer Res. 2000;60:3191–9. [PubMed] [Google Scholar]
- 166.Sohn KJ, Stempak JM, Reid S, Shirwadkar S, Mason JB, Kim YI. The effect of dietary folate on genomic and p53-specific DNA methylation in rat colon. Carcinogenesis. 2003;24:81–90. doi: 10.1093/carcin/24.1.81. [DOI] [PubMed] [Google Scholar]
- 167.Pogribny IP, Karpf AR, James SR, Melnyk S, Han T, Tryndyak VP. Epigenetic alterations in the brains of Fisher 344 rats induced by long-term administration of folate/methyl-deficient diet. Brain Res. 2008;1237:25–34. doi: 10.1016/j.brainres.2008.07.077. [DOI] [PubMed] [Google Scholar]
- 168.Mehedint MG, Niculescu MD, Craciunescu CN, Zeisel SH. Choline deficiency alters global histone methylation and epigenetic marking at the Re1 site of the calbindin 1 gene. FASEB J. 2010;24:184–195. doi: 10.1096/fj.09-140145. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 169.Ghoshal K, Li X, Datta J, Bai S, Pogribny I, Pogribny M, Huang Y, Young D, Jacob ST. A folate- and methyl-deficient diet alters the expression of DNA methyltransferases and methyl CpG binding proteins involved in epigenetic gene silencing in livers of F344 rats. J Nutr. 2006;136:1522–7. doi: 10.1093/jn/136.6.1522. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 170.Lopes S, Lewis A, Hajkova P, Dean W, Oswald J, Forne T, Murrell A, Constancia M, Bartolomei M, Walter J, Reik W. Epigenetic modifications in an imprinting cluster are controlled by a hierarchy of DMRs suggesting long-range chromatin interactions. Hum Mol Genet. 2003;12:295–305. doi: 10.1093/hmg/ddg022. [DOI] [PubMed] [Google Scholar]
- 171.Li E, Beard C, Jaenisch R. Role for DNA methylation in genomic imprinting. Nature. 1993;366:362–365. doi: 10.1038/366362a0. [DOI] [PubMed] [Google Scholar]
- 172.Bell AC, Felsenfeld G. Methylation of a CTCF-dependent boundary controls imprinted expression of the Igf2 gene. Nature. 2000;405:482–485. doi: 10.1038/35013100. [DOI] [PubMed] [Google Scholar]
- 173.Chen DY, Stern SA, Garcia-Osta A, Saunier-Rebori B, Pollonini G, Bambah-Mukku D, Blitzer RD, Alberini CM. A critical role for IGF-II in memory consolidation and enhancement. Nature. 2011;469:491–7. doi: 10.1038/nature09667. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 174.Tahiliani M, Koh KP, Shen Y, Pastor WA, Bandukwala H, Brudno Y, Agarwal S, Iyer LM, Liu DR, Aravind L, Rao A. Conversion of 5-methylcytosine to 5-hydroxymethylcytosine in mammalian DNA by MLL partner TET1. Science. 2009;324:930–5. doi: 10.1126/science.1170116. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 175.Ito S, D’Alessio AC, Taranova OV, Hong K, Sowers LC, Zhang Y. Role of Tet proteins in 5mC to 5hmC conversion, ES-cell self-renewal and inner cell mass specification. Nature. 2010;466:1129–33. doi: 10.1038/nature09303. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 176.Ficz G, Branco MR, Seisenberger S, Santos F, Krueger F, Hore TA, Marques CJ, Andrews S, Reik W. Dynamic regulation of 5-hydroxymethylcytosine in mouse ES cells and during differentiation. Nature. 2011;473:398–402. doi: 10.1038/nature10008. [DOI] [PubMed] [Google Scholar]
- 177.Guo JU, Su Y, Zhong C, Ming GL, Song H. Hydroxylation of 5-Methylcytosine by TET1 Promotes Active DNA Demethylation in the Adult Brain. Cell. 2011;145:423–34. doi: 10.1016/j.cell.2011.03.022. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 178.Ito S, Shen L, Dai Q, Wu SC, Collins LB, Swenberg JA, He C, Zhang Y. Tet Proteins Can Convert 5-Methylcytosine to 5-Formylcytosine and 5-Carboxylcytosine. Science. 2011 doi: 10.1126/science.1210597. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 179.He YF, Li BZ, Li Z, Liu P, Wang Y, Tang Q, Ding J, Jia Y, Chen Z, Li L, Sun Y, Li X, Dai Q, Song CX, Zhang K, He C, Xu GL. Tet-Mediated Formation of 5-Carboxylcytosine and Its Excision by TDG in Mammalian DNA. Science. 2011;333:1303–1307. doi: 10.1126/science.1210944. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 180.Valinluck V, Tsai HH, Rogstad DK, Burdzy A, Bird A, Sowers LC. Oxidative damage to methyl-CpG sequences inhibits the binding of the methyl-CpG binding domain (MBD) of methyl-CpG binding protein 2 (MeCP2) Nucleic Acids Res. 2004;32:4100–8. doi: 10.1093/nar/gkh739. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 181.Jin SG, Kadam S, Pfeifer GP. Examination of the specificity of DNA methylation profiling techniques towards 5-methylcytosine and 5-hydroxymethylcytosine. Nucleic Acids Res. 2010;38:e125. doi: 10.1093/nar/gkq223. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 182.Kinney SM, Chin HG, Vaisvila R, Bitinaite J, Zheng Y, Esteve PO, Feng S, Stroud H, Jacobsen SE, Pradhan S. Tissue specific distribution and dynamic changes of 5-hydroxymethylcytosine in mammalian genome. J Biol Chem. 2011;286:24685–24693. doi: 10.1074/jbc.M110.217083. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 183.Frommer M, McDonald LE, Millar DS, Collis CM, Watt F, Grigg GW, Molloy PL, Paul CL. A genomic sequencing protocol that yields a positive display of 5- methylcytosine residues in individual DNA strands. Proc Natl Acad Sci U S A. 1992;89:1827–1831. doi: 10.1073/pnas.89.5.1827. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 184.Flusberg BA, Webster DR, Lee JH, Travers KJ, Olivares EC, Clark TA, Korlach J, Turner SW. Direct detection of DNA methylation during single-molecule, real-time sequencing. Nat Methods. 2010;7:461–165. doi: 10.1038/nmeth.1459. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 185.Jin SG, Wu X, Li AX, Pfeifer GP. Genomic mapping of 5-hydroxymethylcytosine in the human brain. Nucleic Acids Res. 2011;39:5015–5024. doi: 10.1093/nar/gkr120. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 186.Song CX, Szulwach KE, Fu Y, Dai Q, Yi C, Li X, Li Y, Chen CH, Zhang W, Jian X, Wang J, Zhang L, Looney TJ, Zhang B, Godley LA, Hicks LM, Lahn BT, Jin P, He C. Selective chemical labeling reveals the genome-wide distribution of 5-hydroxymethylcytosine. Nat Biotechnol. 2011;29:68–72. doi: 10.1038/nbt.1732. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 187.Kriaucionis S, Heintz N. The nuclear DNA base 5-hydroxymethylcytosine is present in Purkinje neurons and the brain. Science. 2009;324:929–30. doi: 10.1126/science.1169786. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 188.Szulwach KE, Li X, Li Y, Song CX, Wu H, Dai Q, Irier H, Upadhyay AK, Gearing M, Levey AI, Vasanthakumar A, Godley LA, Chang Q, Cheng X, He C, Jin P. 5-hmC-mediated epigenetic dynamics during postnatal neurodevelopment and aging. Nat Neurosci. 2011;14:1607–1616. doi: 10.1038/nn.2959. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 189.Jenuwein T, Allis CD. Translating the histone code. Science. 2001;293:1074–1080. doi: 10.1126/science.1063127. [DOI] [PubMed] [Google Scholar]
- 190.Jenuwein T. The epigenetic magic of histone lysine methylation. Febs J. 2006;273:3121–35. doi: 10.1111/j.1742-4658.2006.05343.x. [DOI] [PubMed] [Google Scholar]
- 191.Strahl BD, Allis CD. The language of covalent histone modifications. Nature. 2000;403:41–5. doi: 10.1038/47412. [DOI] [PubMed] [Google Scholar]
- 192.Li B, Carey M, Workman JL. The role of chromatin during transcription. Cell. 2007;128:707–19. doi: 10.1016/j.cell.2007.01.015. [DOI] [PubMed] [Google Scholar]
- 193.Cao R, Wang LJ, Wang HB, Xia L, Erdjument-Bromage H, Tempst P, Jones RS, Zhang Y. Role of histone H3 lysine 27 methylation in polycomb-group silencing. Science. 2002;298:1039–1043. doi: 10.1126/science.1076997. [DOI] [PubMed] [Google Scholar]
- 194.Rea S, Eisenhaber F, O’Carroll D, Strahl BD, Sun ZW, Schmid M, Opravil S, Mechtler K, Ponting CP, Allis CD, Jenuwein T. Regulation of chromatin structure by site-specific histone H3 methyltransferases. Nature. 2000;406:593–9. doi: 10.1038/35020506. [DOI] [PubMed] [Google Scholar]
- 195.Bernstein BE, Mikkelsen TS, Xie X, Kamal M, Huebert DJ, Cuff J, Fry B, Meissner A, Wernig M, Plath K, Jaenisch R, Wagschal A, Feil R, Schreiber SL, Lander ES. A bivalent chromatin structure marks key developmental genes in embryonic stem cells. Cell. 2006;125:315–26. doi: 10.1016/j.cell.2006.02.041. [DOI] [PubMed] [Google Scholar]
- 196.Barski A, Cuddapah S, Cui K, Roh TY, Schones DE, Wang Z, Wei G, Chepelev I, Zhao K. High-resolution profiling of histone methylations in the human genome. Cell. 2007;129:823–37. doi: 10.1016/j.cell.2007.05.009. [DOI] [PubMed] [Google Scholar]
- 197.Mikkelsen TS, Ku M, Jaffe DB, Issac B, Lieberman E, Giannoukos G, Alvarez P, Brockman W, Kim TK, Koche RP, Lee W, Mendenhall E, O’Donovan A, Presser A, Russ C, Xie X, Meissner A, Wernig M, Jaenisch R, Nusbaum C, Lander ES, Bernstein BE. Genome-wide maps of chromatin state in pluripotent and lineage-committed cells. Nature. 2007;448:553–60. doi: 10.1038/nature06008. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 198.Heintzman ND, Hon GC, Hawkins RD, Kheradpour P, Stark A, Harp LF, Ye Z, Lee LK, Stuart RK, Ching CW, Ching KA, Antosiewicz-Bourget JE, Liu H, Zhang X, Green RD, Lobanenkov VV, Stewart R, Thomson JA, Crawford GE, Kellis M, Ren B. Histone modifications at human enhancers reflect global cell-type-specific gene expression. Nature. 2009;459:108–112. doi: 10.1038/nature07829. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 199.Hawkins RD, Hon GC, Lee LK, Ngo Q, Lister R, Pelizzola M, Edsall LE, Kuan S, Luu Y, Klugman S, Antosiewicz-Bourget J, Ye Z, Espinoza C, Agarwahl S, Shen L, Ruotti V, Wang W, Stewart R, Thomson JA, Ecker JR, Ren B. Distinct epigenomic landscapes of pluripotent and lineage-committed human cells. Cell Stem Cell. 2010;6:479–91. doi: 10.1016/j.stem.2010.03.018. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 200.Noma K, Allis CD, Grewal SI. Transitions in distinct histone H3 methylation patterns at the heterochromatin domain boundaries. Science. 2001;293:1150–5. doi: 10.1126/science.1064150. [DOI] [PubMed] [Google Scholar]
- 201.Schneider R, Bannister AJ, Myers FA, Thorne AW, Crane-Robinson C, Kouzarides T. Histone H3 lysine 4 methylation patterns in higher eukaryotic genes. Nat Cell Biol. 2004;6:73–7. doi: 10.1038/ncb1076. [DOI] [PubMed] [Google Scholar]
- 202.Strahl BD, Ohba R, Cook RG, Allis CD. Methylation of histone H3 at lysine 4 is highly conserved and correlates with transcriptionally active nuclei in Tetrahymena. Proc Natl Acad Sci U S A. 1999;96:14967–72. doi: 10.1073/pnas.96.26.14967. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 203.Aagaard L, Laible G, Selenko P, Schmid M, Dorn R, Schotta G, Kuhfittig S, Wolf A, Lebersorger A, Singh PB, Reuter G, Jenuwein T. Functional mammalian homologues of the Drosophila PEV-modifier Su(var)3–9 encode centromere-associated proteins which complex with the heterochromatin component M31. EMBO J. 1999;18:1923–38. doi: 10.1093/emboj/18.7.1923. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 204.Zhu XN, Mar MH, Song JN, Zeisel SH. Deletion of the Pemt gene increases progenitor cell mitosis, DNA and protein methylation and decreases calretinin expression in embryonic day 17 mouse hippocampus. Dev Brain Res. 2004;149:121–129. doi: 10.1016/j.devbrainres.2004.01.004. [DOI] [PubMed] [Google Scholar]
- 205.Dobosy JR, Fu VX, Desotelle JA, Srinivasan R, Kenowski ML, Almassi N, Weindruch R, Svaren J, Jarrard DF. A methyl-deficient diet modifies histone methylation and alters Igf2 and H19 repression in the prostate. Prostate. 2008;68:1187–95. doi: 10.1002/pros.20782. [DOI] [PubMed] [Google Scholar]
- 206.Davison JM, Mellott TJ, Kovacheva VP, Blusztajn JK. Gestational choline supply regulates methylation of histone H3, expression of histone methyltransferases G9a (Kmt1c) and Suv39h1 (Kmt1a) and DNA methylation of their genes in rat fetal liver and brain. J Biol Chem. 2009;284:1982–1989. doi: 10.1074/jbc.M807651200. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 207.Yang Y, Liu Z, Cermak JM, Tandon P, Sarkisian MR, Stafstrom CF, Neill JC, Blusztajn JK, Holmes GL. Protective effects of prenatal choline supplementation on seizure-induced memory impairment. J Neurosci. 2000;20:RC109. doi: 10.1523/JNEUROSCI.20-22-j0006.2000. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 208.Holmes GL, Yang Y, Liu Z, Cermak JM, Sarkisian MR, Stafstrom CE, Neill JC, Blusztajn JK. Seizure-induced memory impairment is reduced by choline supplementation before or after status epilepticus. Epilepsy Res. 2002;48:3–13. doi: 10.1016/s0920-1211(01)00321-7. [DOI] [PubMed] [Google Scholar]
- 209.Wong-Goodrich SJ, Mellott TJ, Liu B, Blusztajn JK, Williams CL. Water maze experience and prenatal choline supplementation differentially promote long-term hippocampal recovery from seizures in adulthood. Hippocampus. 2011;21:584–608. doi: 10.1002/hipo.20783. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 210.Thomas JD, La Fiette MH, Quinn VR, Riley EP. Neonatal choline supplementation ameliorates the effects of prenatal alcohol exposure on a discrimination learning task in rats. Neurotoxicol Teratol. 2000;22:703–711. doi: 10.1016/s0892-0362(00)00097-0. [DOI] [PubMed] [Google Scholar]
- 211.Thomas JD, Biane JS, O’Bryan KA, O’Neill TM, Dominguez HD. Choline supplementation following third-trimester-equivalent alcohol exposure attenuates behavioral alterations in rats. Behav Neurosci. 2007;121:120–30. doi: 10.1037/0735-7044.121.1.120. [DOI] [PubMed] [Google Scholar]
- 212.Ryan SH, Williams JK, Thomas JD. Choline supplementation attenuates learning deficits associated with neonatal alcohol exposure in the rat: effects of varying the timing of choline administration. Brain Res. 2008;1237:91–100. doi: 10.1016/j.brainres.2008.08.048. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 213.Thomas JD, Abou EJ, Dominguez HD. Prenatal choline supplementation mitigates the adverse effects of prenatal alcohol exposure on development in rats. Neurotoxicol Teratol. 2009;31:303–11. doi: 10.1016/j.ntt.2009.07.002. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 214.Thomas JD, Idrus NM, Monk BR, Dominguez HD. Prenatal choline supplementation mitigates behavioral alterations associated with prenatal alcohol exposure in rats. Birth Defects Res A Clin Mol Teratol. 2010;88:827–37. doi: 10.1002/bdra.20713. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 215.Thomas JD, Tran TD. Choline supplementation mitigates trace, but not delay, eyeblink conditioning deficits in rats exposed to alcohol during development. Hippocampus. 2011;22:619–630. doi: 10.1002/hipo.20925. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 216.Ricceri L, Berger-Sweeney J. Postnatal choline supplementation in preweanling mice: sexually dimorphic behavioral and neurochemical effects. Behav Neurosci. 1998;112:1387–1392. [PubMed] [Google Scholar]
- 217.Nag N, Berger-Sweeney JE. Postnatal dietary choline supplementation alters behavior in a mouse model of Rett syndrome. Neurobiol Dis. 2007;26:473–80. doi: 10.1016/j.nbd.2007.02.003. [DOI] [PubMed] [Google Scholar]
- 218.Stevens KE, Adams CE, Yonchek J, Hickel C, Danielson J, Kisley MA. Permanent improvement in deficient sensory inhibition in DBA/2 mice with increased perinatal choline. Psychopharmacology (Berl) 2008;198:413–420. doi: 10.1007/s00213-008-1170-3. [DOI] [PubMed] [Google Scholar]
- 219.Nag N, Mellott TJ, Berger-Sweeney JE. Effects of postnatal dietary choline supplementation on motor regional brain volume and growth factor expression in a mouse model of Rett syndrome. Brain Res. 2008;1237:101–9. doi: 10.1016/j.brainres.2008.08.042. [DOI] [PubMed] [Google Scholar]
- 220.Ward BC, Agarwal S, Wang K, Berger-Sweeney J, Kolodny NH. Longitudinal brain MRI study in a mouse model of Rett Syndrome and the effects of choline. Neurobiol Dis. 2008;31:110–9. doi: 10.1016/j.nbd.2008.03.009. [DOI] [PubMed] [Google Scholar]
- 221.Ward BC, Kolodny NH, Nag N, Berger-Sweeney JE. Neurochemical changes in a mouse model of Rett syndrome: changes over time and in response to perinatal choline nutritional supplementation. J Neurochem. 2009;108:361–71. doi: 10.1111/j.1471-4159.2008.05768.x. [DOI] [PubMed] [Google Scholar]
- 222.Moon J, Chen M, Gandhy SU, Strawderman M, Levitsky DA, Maclean KN, Strupp BJ. Perinatal choline supplementation improves cognitive functioning and emotion regulation in the Ts65Dn mouse model of Down syndrome. Behav Neurosci. 2010;124:346–61. doi: 10.1037/a0019590. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 223.Kovacheva VP, Davison JM, Mellott TJ, Rogers AE, Yang S, O’Brien MJ, Blusztajn JK. Raising gestational choline intake alters gene expression in DMBA-evoked mammary tumors and prolongs survival. FASEB J. 2009;23:1054–63. doi: 10.1096/fj.08-122168. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 224.Chen WG, Chang Q, Lin YX, Meissner A, West AE, Griffith EC, Jaenisch R, Greenberg ME. Derepression of BDNF transcription involves calcium-dependent phosphorylation of MeCP2. Science. 2003;302:885–889. doi: 10.1126/science.1086446. [DOI] [PubMed] [Google Scholar]
- 225.Martinowich K, Hattori D, Wu H, Fouse S, He F, Hu Y, Fan GP, Sun YE. DNA methylation-related chromatin remodeling in activity-dependent Bdnf gene regulation. Science. 2003;302:890–893. doi: 10.1126/science.1090842. [DOI] [PubMed] [Google Scholar]
- 226.Levenson JM, Roth TL, Lubin FD, Miller CA, Huang IC, Desai P, Malone LM, Sweatt JD. Evidence that DNA (cytosine-5) methyltransferase regulates synaptic plasticity in the hippocampus. J Biol Chem. 2006;281:15763–73. doi: 10.1074/jbc.M511767200. [DOI] [PubMed] [Google Scholar]
- 227.Nelson ED, Kavalali ET, Monteggia LM. Activity-dependent suppression of miniature neurotransmission through the regulation of DNA methylation. J Neurosci. 2008;28:395–406. doi: 10.1523/JNEUROSCI.3796-07.2008. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 228.Yossifoff M, Kisliouk T, Meiri N. Dynamic changes in DNA methylation during thermal control establishment affect CREB binding to the brain-derived neurotrophic factor promoter. Eur J Neurosci. 2008;28:2267–77. doi: 10.1111/j.1460-9568.2008.06532.x. [DOI] [PubMed] [Google Scholar]
- 229.Miller CA, Sweatt JD. Covalent modification of DNA regulates memory formation. Neuron. 2007;53:857–69. doi: 10.1016/j.neuron.2007.02.022. [DOI] [PubMed] [Google Scholar]
- 230.Lubin FD, Roth TL, Sweatt JD. Epigenetic regulation of BDNF gene transcription in the consolidation of fear memory. J Neurosci. 2008;28:10576–86. doi: 10.1523/JNEUROSCI.1786-08.2008. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 231.Miller CA, Gavin CF, White JA, Parrish RR, Honasoge A, Yancey CR, Rivera IM, Rubio MD, Rumbaugh G, Sweatt JD. Cortical DNA methylation maintains remote memory. Nat Neurosci. 2010;13:664–6. doi: 10.1038/nn.2560. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 232.Gupta S, Kim SY, Artis S, Molfese DL, Schumacher A, Sweatt JD, Paylor RE, Lubin FD. Histone methylation regulates memory formation. J Neurosci. 2010;30:3589–99. doi: 10.1523/JNEUROSCI.3732-09.2010. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 233.Poly C, Massaro JM, Seshadri S, Wolf PA, Cho E, Krall E, Jacques PF, Au R. The relation of dietary choline to cognitive performance and white-matter hyperintensity in the Framingham Offspring Cohort. Am J Clin Nutr. 2011;94:1584–91. doi: 10.3945/ajcn.110.008938. [DOI] [PMC free article] [PubMed] [Google Scholar]