

Abstract
Wound healing is a complex process marked by highly coordinated immune fluxes into an area of tissue injury; these are required for re-establishment of normal tissue integrity. Along with this cascade of cellular players, wound healing also requires coordinated flux through a number of biochemical pathways, leading to synthesis of collagen and recycling or removal of damaged tissues. The availability of nutrients, especially amino acids, is critical for wound healing, and enteral supplementation has been intensely studied as a potential mechanism to augment wound healing—either by increasing tensile strength, decreasing healing time, or both. From a practical standpoint, although enteral nutrient supplementation may seem like a reasonable strategy to augment healing, a number of biochemical and physiologic barriers exist that limit this strategy. In this critical review, the physiology of enteral amino acid metabolism and supplementation and challenges therein are discussed in the context of splanchnic physiology and biochemistry. Additionally, a review of studies examining various methods of amino acid supplementation and the associated effects on wound outcomes are discussed.
Keywords: wound healing, nutrient supplementation, wound strength, amino acid supplementation, metabolism, surgical metabolism, collagen metabolism
Introduction
The phases of wound healing include coagulation, inflammation, proliferation, and maturation, and each phase involves a complex but well-orchestrated cascade of biochemical and immunologic events (1). The synthesis, breakdown, and resynthesis of collagen represents a critical factor in the re-establishment of tissue integrity. With the exception of superficial injuries affecting only the epidermis, most wounds heal through the formation of scar tissue, which is predominately composed of collagen. The remodeling and synthesis of collagen is the final, longest, and most critical step in the process of wound healing (2). Metabolically, the process requires ATP and amino acid precursors, as well as various trace minerals, vitamins, and oxygen as cofactors. Therefore, sufficient energy and protein substrate availability is essential in ensuring successful wound fibroplasia.
Wound healing and nutrient intake are intimately associated; this relation has been appreciated for decades. In fact, this observation has been commonly stated as “wound nutrition is in fact whole-body nutrition” (3). Nutritional status, which is a major determinant for wound outcome, can be measured by any number of factors, including successfully completed healing and lack of infection or other complications (4). However, the question of whether the administration of certain nutrients can specifically enhance collagen synthesis or lead to improved outcomes continues to be unanswered. As a result, the administration of various amino acids has been studied extensively. The ability to augment wound healing with nutrient supplementation could hypothetically lead to improved wound strength and potentially lower rates of short- and long-term wound complications (e.g., infection or hernia formation). Improving these factors would not only have important clinical benefits and lead to lower health care costs but also improve quality of life. In the following, we review the biochemical mechanisms of collagen synthesis and the potential pathways that could be exploited to potentially augment wound healing. The physiologic challenges and studies examining the putative effectiveness of nutritional therapies are discussed herein.
Biochemical Considerations of Collagen Synthesis
Biochemically, collagen is approximately one-third glycine (Figure 1), such that every third amino acid is a glycine molecule, according to the formula GLY-X-Y. The next most prevalent amino acid is proline or its derivative, hydroxyproline; these frequently occupy the X or Y positions. The X and Y positions can be filled by any number of amino acids, which are diagrammed in terms of typical abundance in Figure 1. Together, proline and hydroxyproline comprise ∼23% of the amino acid content of the collagen molecule (6, 7). Of the body’s stores of hydroxyproline, ∼99.8% are found in collagen, rendering assays of this amino acid useful as a marker for the total amount of collagen. The hydroxylation of proline occurs posttranslationally and is carried out by the enzyme prolyl hydroxylase, which requires oxygen, iron, and ascorbate (i.e., vitamin C) as cofactors for successful activity. In fact, vitamin C is critical to wound healing; in addition to its importance in serious illness, patients can benefit substantially from vitamin C administration even after elective operations (8). Although vitamin and mineral requirements may be increasingly important to wound healing and other biologic processes, we have limited our current discussion to amino acid biochemistry. Both proline and hydroxyproline are vital for collagen biosynthesis, structure, and strength. Their cyclic structure constrains the rotation of the polypeptide collagen chain, creating and strengthening the helical characteristic of the molecule (7).
FIGURE 1.

Schematic representation of the amino acid composition of collagen. Composite from various sources; reproduced from reference 5 with permission.
Plasma proline concentration can be variable, but its importance to human nutrition is not clear. Bona fide proline deficiency likely occurs only in prolidase deficiency, an autosomal recessive inborn metabolic error in which a lack of prolidase prevents degradation of (proline-containing) iminodipeptides liberated during collagen breakdown; excretion of these dipeptides can be detected in the urine. Patients with prolidase deficiency have low proline levels and, as might be expected, a variety of wound-healing deficits (9).
The availability of proline is critical to the synthesis of collagen as well as other proteins, and the body maintains strict control over proline bioavailability. Proline handling is at least partially dependent on the route of administration; the small intestine takes up considerable loads of dietary proline without selectively clearing arterial proline in large quantities (10). Excessive dietary proline can be metabolized within small intestinal enterocytes via proline oxidase (also known as proline dehydrogenase) in the reaction:
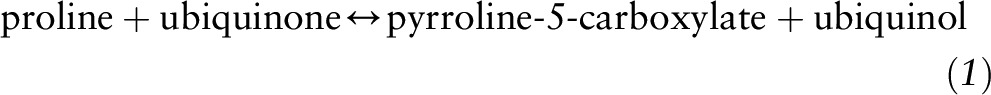 |
This reaction is the major pathway for proline degradation and is ubiquitous in mammals (11). It also produces pyrroline-5-carboxylate (P5C), an important intermediate that links the citric acid and urea cycles (Figure 2). P5C can be converted back to proline via P5C reductase in the presence of either NAD+ or NADP+ in a reaction that is favored when collagen production is increased:
FIGURE 2.

Proline and glutamate metabolism are functional links between the TCA and urea cycles. A number of enzymes and biochemical transformations link the metabolism of proline intermediates; however, many of these reactions are physically separated within the cell. Glutamine and glutamate can be interconverted by the actions of GS and GLN, respectively. The anaplerotic reaction converting glutamate to α-ketoglutarate is catalyzed by GDH and heavily favors the synthesis of α-ketoglutarate. Glutamate can alternatively be metabolized to P5C by P5CS or P5C can be metabolized to glutamate by P5CDH. P5C and GSA spontaneously interconvert with P5C and can be converted to proline via the action of P5CR, whereas the reverse reaction is catalyzed by POX that can also be referred to in some texts as PDH. Additionally, GSA can be metabolized to ornithine via OAT to replenish the urea cycle. Other urea cycle intermediates, including citrulline and arginine, can be given exogenously to produce ornithine that can be subsequently used for proline synthesis. AS, argininosuccinate; ASL, argininosuccinate lyase; CPS-I, carbamoyl phosphate synthetase I; CoA, coenzyme A; GDH, glutamate dehydrogenase; GLN, glutaminase; GS, glutamine synthase; GSA, glutamate-semialdehyde; OAA, oxaloacetic acid; OAT, ornithine aminotransferase; OTC, ornithine transcarbamylase; PDH, proline dehydrogenase; POX, proline oxidase; P5C, pyrroline-5-carboxylate; P5CDH, P5C dehydrogenase; P5CR, P5C reductase; P5CS, P5C synthase; spont., spontaneous; TCA, tricarboxylic acid.
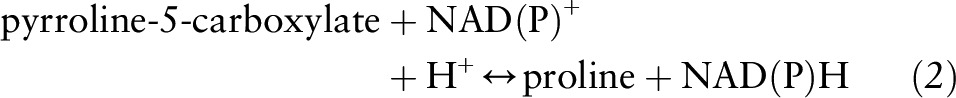 |
Alternatively, P5C and its tautomer, glutamate-5-semialdehyde (GSA), are interchangeable with a number of amino acids and metabolites through a variety of transformations linking the citric acid and urea cycles. Other amino acids that can be interconverted to proline and ultimately incorporated into collagen include glutamine, glutamate, arginine, and ornithine (12). Glutamate and glutamine are biochemically interchangeable and are important sources of P5C for proline synthesis. Glutamine can be deaminated to glutamate via the action of glutaminase with the concomitant production of ammonia:
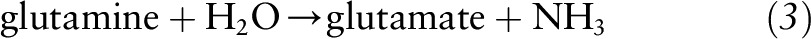 |
The glutamate produced via glutaminase, as well as other sources of glutamate (e.g., dietary), can readily be transformed by enzymes involved in glutamate catabolism. Glutamate dehydrogenase oxidizes glutamate to α-ketoglutarate (anaplerosis) with the concomitant reduction of NADP+ to NADPH in the following reaction:
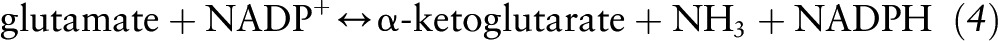 |
Although this reaction is reversible, the conditions favoring the reverse reaction leading to glutamate production from α-ketoglutarate typically do not occur under typical cellular conditions. Conversely, glutamate can be generated through the action of GSA dehydrogenase that catalyzes conversion of GSA to glutamate with concomitant reduction of NADP+ to NADPH:
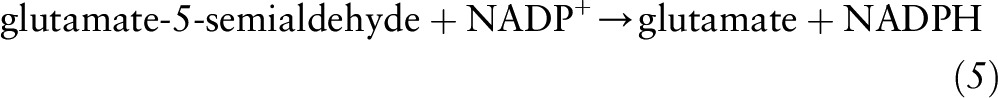 |
Alternatively, glutamate can be generated by the action of P5C dehydrogenase from conversion of P5C to glutamate:
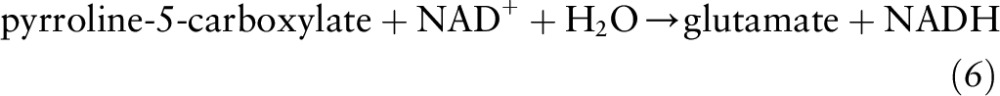 |
Aside from glutamate and glutamine, arginine and ornithine are also readily metabolized to P5C and subsequently converted to other amino acids or intermediates. In most tissues, low levels of arginase expression are available to convert arginine to ornithine through the irreversible final step in the urea cycle:
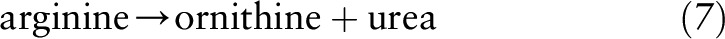 |
Subsequently, ornithine can be converted to P5C by the action of ornithine aminotransferase (OAT):
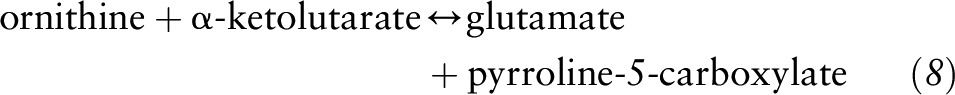 |
The action of OAT is readily reversible and appears to be dependent on the availability of both substrate and products within the cell. Thus, when intracellular ornithine concentration is low, the flux of this pathway is toward ornithine. Conversely, increased ornithine concentration will promote production of P5C, which can be used for proline biosynthesis or conversion to glutamate or glutamine in the previously mentioned reactions (13). Overall, the cell- and tissue-specific expression of these enzymes allows for rapid interconversion of substrates to meet the synthetic demands of optimal wound healing.
Physiologic and Biochemical Challenges to Augmenting Collagen Synthesis
Enterocyte amino acid catabolism.
As alluded to previously, dietary nutrient supplementation of proline or its precursor metabolites is hypothetically advantageous and may improve wound healing through various mechanisms. However, this concept is more complicated than it may seem at first glance. In fact, a number of amino acids are highly metabolized in the splanchnic tissues and do not appear in large quantities in the peripheral circulation; these include glutamine (14), glutamate (15–17), and aspartate (18). Previously, it was thought that the liver played a role in regulating peripheral concentrations of enteral amino acids; however, many of these nutrients are actually metabolized within the mucosal enterocytes before they reach the portal circulation or liver (19, 20). Thus, for glutamine, glutamate, and aspartate, dietary supplementation does not necessarily lead to corresponding increases in the peripheral blood.
Unlike glutamine, glutamate, and aspartate, arginine and proline are handled differently in the splanchnic tissues. In the case of proline, although early studies have suggested that enteral proline was not significantly metabolized in the splanchnic tissues (10), later studies have demonstrated the synthesis of arginine, ornithine, and citrulline from enteral proline (21–23). Biochemically, proline oxidase and OAT are highly expressed in the small intestine, allowing for increased proline catabolism despite high dietary loads. There is likely a physiologic limit to the amount of proline that reaches the portal, and eventually the peripheral circulation; these mechanisms have yet to be defined. To add to the complexity of clinical translation, enteral amino acid metabolism likely differs based on the developmental state of the individual (e.g., a neonate compared with an adult) and may change depending on short- and long-term nutrient exposure (24).
Splanchnic-systemic trafficking of metabolites.
Unlike that of the aforementioned amino acids, de novo arginine synthesis requires organ-to-organ shuttling of substrates, referred to as the intestinal-renal axis. Arginine and arginine precursors can be converted to citrulline within the enterocyte; subsequently, this citrulline escapes hepatic clearance in the portal circulation (25). This allows citrulline to bypass the liver and pass into the systemic circulation, where it can then be extracted by the kidney and converted to arginine. This intestinal-renal axis exists because the small intestine lacks expression of 2 urea cycle enzymes (Figure 2), argininosuccinate synthase and argininosuccinate lyase (26), thereby permitting accumulation of citrulline in the intestine. Upon reaching the kidneys, which have minimal arginase expression and markedly higher expression of argininosuccinate synthase and argininosuccinate lyase, citrulline is readily converted to arginine (27). This regulation of arginine and citrulline synthesis is complex and is also affected by dietary protein content that alters the expression of arginine- and citrulline-metabolizing enzymes to fine-tune the hepatic delivery of arginine and citrulline (28).
Mass action and regulation of cellular metabolic pathways.
Even if sufficient collagen-promoting precursors could be delivered to the cells and tissues involved in wound healing, there would still exist further intracellular mechanisms controlling metabolism. Subcellular localization of enzymes and substrates is critically important to cellular synthetic and oxidative function because synthetic and degradative biochemical pathways are physically separated within the cell to prevent futile cycling of metabolites (e.g., mitochondrial FA oxidation compared with cytosolic FA synthesis). Aside from anabolic and catabolic considerations, the arginine paradox is a prime example of the importance of subcellular localization limiting enzymatic activity. The paradox is that the arginine concentration within endothelial cells far exceeds the Michaelis constant (Km) for endothelial NO synthase; however, intravenous infusion of exogenous arginine causes vasodilation in a manner dependent on endothelial NO synthase. Clearly, these effects demonstrate that much remains to be understood about the regulation of the cell. Aside from compartmentalization of metabolic pathways, numerous other layers of regulation exist that must be overcome to promote flux through a particular metabolic pathway. These additional layers of regulation not only include classical negative feedback regulation and allosteric effects of substrate-enzyme interactions, but also substrate-level phosphorylation via cell-signaling mechanisms and transcriptional effects (29). Overall, a strategy to specifically augment proline and, subsequently, collagen synthesis is met with layers of regulation and challenges to be overcome clinically.
Clinical and Preclinical Data Examining Nutrient Supplementation and Wound Healing
Despite the theoretical challenges associated with augmenting collagen synthesis in vivo, a number of approaches have been used to examine various dietary or topical supplements. One approach is to examine the structure of collagen and pursue provision of substrates based on its molecular composition. During the first 10 d of healing, wound proline levels are 30–50% higher than plasma levels, suggesting that the importation of proline into the wound occurs actively or that the biosynthesis of proline takes place in the wound environment (30). Conversely, some of the proline could also be generated from the lysis of collagen, which occurs in parallel to synthesis. The high demand for proline during wound repair could cause a local proline deficiency; theoretically, therefore, enhancing proline bioavailability would be an attractive strategy to optimize collagen biosynthesis.
Although the most straightforward approach to improve wound strength is to provide additional proline in the diet, neither proline nor hydroxyproline increase wound breaking strength (31). Ingested hydroxyproline is readily degraded and synthesis of hydroxyproline occurs only posttranslationally—not de novo—once proline has already been incorporated into collagen. In experiments at our laboratory, male Sprague Dawley rats weighing 250–300 g were given a 1% dietary supplement of proline (proline:total diet, wt:wt) in addition to a complete nonpurified diet adequate for growth and reproduction. All animals underwent dorsal skin incision and subcutaneous implantation of polyvinyl alcohol sponges. After 10 d, we found no difference in wound breaking strength or in wound collagen deposition as assessed by the amount of hydroxyproline present in the subcutaneously implanted sponges (A Barbul, unpublished results, 2008). As mentioned previously, it is not surprising that supplemental dietary proline fails to increase the abundance of wound collagen; this likely reflects the complex metabolic regulation that controls cellular concentrations of proline or other collagen substrates.
Glutamine and glutamate.
The citric acid cycle is the source of a large amount of glutamate, and this production couples the citric acid cycle to proline biosynthesis. However, glutamate supplementation does not lead to increases in proline synthesis, and large amounts of dietary glutamate are not well tolerated. Conversely, glutamine has been extensively studied. Glutamine is the most abundant amino acid in the body, accounting for ∼20% of the total circulating free amino acid pool and 60% of the free intracellular amino acid pool in most tissues. Glutamine is a critical fuel for many cells, including fibroblasts, macrophages, neutrophils, and lymphocytes, all of which participate in wound healing (32). Scientifically, it is well understood that almost all cells require exogenous glutamine for growth and function in vitro.
Given the abundance of glutamine in cells that contribute to wound healing, it is not surprising that there is a rapid decrease in plasma and muscle glutamine concentration following injury (33). This decrease is greater than that of any other amino acid. Although efficacy of supplemental glutamine administration has been shown in some clinically important conditions, it has not proved to have any noticeable effect on wound healing (4). In vitro administration of 0.15 mmol/L glutamine (without glutamate or other glutamine derivatives) in human dermal confluent fibroblast cultures increases collagen synthesis at the mRNA level 1.7–2.3-fold (34). It is postulated that this might occur via a metabolic intermediary, P5C (35). Reports have demonstrated the benefit of glutamine supplementation alone on wound collagen synthesis and accumulation in primary or secondary (36) wounds. Specifically, a study of air leak pressures after deliberate pulmonary laceration in a group of Wistar albino rats indicated that rats receiving glutamine supplementation had higher air leak pressures than control animals, and that these animals also had higher levels of immature collagen, as demonstrated by histopathology (37). A second study similarly demonstrated greater burst strength in colon anastomoses in rats with glutamine supplementation than those with glycine supplementation, and also noted higher levels of mature collagen in the anastomotic colon tissue of the glutamine group (38). An additional study demonstrated a subjective trend toward less edema and lower neutrophil cell counts as well as less bacterial colonization and abscess formation in a rat colotomy model. Although subjective improvements were reported, burst strength was not different between glutamine-supplemented and control animals (39). Most of the benefits of supplemental glutamine appear to involve improved gut permeability and protein synthesis, normalization of serum levels, and decreased length of hospital stay (40). In addition, there is some evidence that preinjury glutamine administration can prevent damage to collagen expression caused by external beam radiation (41). Furthermore, a mixture of BCAAs and glutamine improved the fractional synthesis rate of dermal tropocollagen by a factor of 2.04 ± 0.54 (mean ± SE) in an immunocompetent mouse irradiation model (42). Thus, glutamine supplementation has demonstrated some possible wound-healing benefits that need to be further evaluated in clinical studies for possible translation to clinical practice.
There have also been multiple studies concerning the use of glutamine in concert with other amino acids; however, these studies have demonstrated conflicting results. A study from our laboratory demonstrated that daily administration of a mixture of 14 g arginine, 3 g β-hydroxy-β-methylbutyrate, and 14 g glutamine in 2 divided doses to healthy volunteers resulted in an increase in the collagen deposition in subcutaneously implanted polytetrafluoroethylene catheters as measured by hydroxyproline content. This was not accompanied by increased overall protein deposition as measured by catheter α-amino acid nitrogen abundance (43). Similarly, BCAAs and glutamine supplementation increases the fractional synthesis rate of dermal collagen in rats with protein malnourishment (44). Administration of a mixture of arginine, glutamine, and β-hydroxy-β-methylbutyrate was also reported to decrease wound sizes at 10 and 14 d after injury (45). Administration of these same amino acids also increased hydroxyproline content in diabetic foot ulcers (46), but not in a basic science model of the same injury (47). Regardless, there is considerable variation in supplementation and wound outcomes and definitive studies are needed to determine whether or not glutamine could be used therapeutically, either alone or in combination with other amino acids, to facilitate wound healing in a clinical setting.
Arginine and ornithine.
As mentioned earlier, the urea cycle can also provide proline biosynthetic precursors through conversion of arginine to ornithine via arginase, a key urea cycle enzyme. Ornithine, through the action of OAT, can subsequently be converted to GSA, linking ornithine to proline synthesis. In naturally occurring proteins, arginine is by far more prevalent and available than ornithine.
In humans, arginine is synthesized in adequate quantities to sustain muscle and connective tissue mass but likely insufficient quantities for optimal collagen biosynthesis and healing following injury. Serious physiologic stress or injury rapidly decreases total body arginine stores. During periods of stress, endogenous arginine synthesis is insufficient to meet the demands of increased protein turnover. Thus, arginine becomes conditionally essential and necessary for wound healing and the maintenance of a positive nitrogen balance (48).
This conditionally essential status of arginine has been well documented and studied in many model systems. The role of arginine in wound healing was first shown in animals that were fed an arginine-deficient diet for 4–6 wk and were subjected to the minor trauma of a dorsal skin incision and closure. In this model, animals manifested increased postoperative weight loss, increased mortality (in a model system that typically has a mortality rate of 0), and a notable decrease in wound breaking strength and wound collagen accumulation compared with animals fed a diet containing arginine (49). Follow-up experiments using stock diet–fed, arginine-replete rats that were then fed a diet containing an additional 1% arginine (wt:wt) demonstrated improved wound healing (48). Similar findings were observed in parenterally fed rats supplemented with high doses (7.5 g/L) of arginine (50).
Clinically, 2 studies in healthy human volunteers examined the effects of arginine supplementation. These studies have demonstrated that arginine (in dosages of 17–24.8 g/d) enhances collagen deposition in healthy adult and elderly human volunteers (51, 52). Arginine supplementation did not affect the rate of epithelialization of a superficial skin defect, suggesting that the predominate effect of arginine supplementation is wound collagen deposition (52).
Several mechanisms have been postulated to explain the positive effect of arginine on wound healing, but many questions still exist. Although arginine comprises a small amount of the collagen molecule (<5%) (Figure 1), it is feasible that supplemental arginine provides substrate precursors for conversion of arginine → ornithine → GSA → proline pathway (Figure 2). Support for this hypothesis is demonstrated by the observation that arginine concentrations within the wound are essentially undetectable during later phases of healing—a time when fibroplasia predominates—which indicates a local wound arginine-deficient state (53). Further support for this hypothetical pathway has been demonstrated in experiments actively supplementing ornithine. Stock diet–fed mice that were supplemented with 1% ornithine demonstrated enhanced wound breaking strength and collagen deposition, similar in magnitude to the effect in mice supplemented with 1% arginine (54) (Figure 3A, B). Moreover, ornithine concentrations in wound fluid are typically higher than plasma concentrations, but there is also higher arginase activity. Ornithine supplementation further elevates plasma and wound fluid ornithine levels. The rate of ornithine conversion to proline has been estimated in vivo at <5%; in vitro studies using whole cartilage and cell preparations estimate a 20% conversion of ornithine into the total proline incorporated into protein (53, 56, 57). This marked discrepancy might reflect methodical differences between tracer studies and incorporation experiments.
FIGURE 3.

Effect of 14-d dietary supplementation with 1% arginine (A) or ornithine (B) on wound breaking strength and wound collagen deposition in implantable dermal sponges from wild-type and iNOS KO male mice. *,**Significant differences from control, P < 0.05 and P < 0.01, respectively (Student’s t test). Values are means ± SEs; n = 10–12/group. iNOS, inducible NO synthase; KO, knockout. Composite adapted from references 54 and 55 with permission.
Aside from arginine and ornithine, another urea cycle intermediate of potential importance is citrulline. Citrulline is a precursor for arginine and is able to nutritionally replace arginine in species requiring arginine for growth (58). Additionally, dietary citrulline elevates plasma arginine concentrations in short-gut animal models (59). On the other hand, citrulline does not share any of the other biological or pharmacologic effects of arginine, nor does it have any effect on wound healing or collagen synthesis (48). Interestingly, ornithine is unable to replace arginine for growth requirement but does continue to share similar effects on wound healing as mentioned previously.
Arginine is also the substrate for the generation of NO, a critical player in wound healing (60). Consistent with this role in wound healing, functional loss of inducible NO synthase (iNOS) abrogates the benefits of arginine on wound healing (55) (Figure 3A). On the other hand, ornithine increases wound breaking strength and collagen accumulation in iNOS-knockout mice to the same extent as in wild-type animals (54) (Figure 3B), suggesting that iNOS is at least partially responsible for the improved wound healing response secondary to arginine, but not that of ornithine (55). Nevertheless, these observations do not invalidate the roles played by both arginine and ornithine as precursors of proline synthesis.
In a series of experiments, we examined whether simultaneous dietary administration of arginine and proline administration results in enhanced collagen deposition. In male adult Sprague Dawley rats undergoing dorsal skin incision and subcutaneous placement of polyvinyl alcohol sponges, no additive effect of proline was noted beyond the effect of arginine alone, suggesting that arginine provides sufficient precursor for proline biosynthesis (A Barbul, unpublished results, 2005).
In addition to effects on wound healing, arginine and ornithine are also strong secretagogues of growth hormone, which might also underlie some of their effects on wound collagen deposition. Moreover, both are also stimulators of T cell function and T lymphocytes, which are necessary for normal wound healing (1). How much these effects contribute to the positive effects of arginine and ornithine on wound healing is unknown; future studies could address these areas to understand how these precursors affect endocrine function and wound healing.
Aside from citric acid and urea cycle precursors that could be used for proline biosynthesis, ornithine-α-ketoglutarate (OKG) could also be used as a potential supplement. OKG has been studied extensively and is noted to have beneficial effects on nitrogen balance, protein metabolism, and recovery from trauma and sepsis. It has also been shown to reduce healing time following burn injury in both animals and humans. OKG administration also elevates plasma proline levels in both healthy volunteers and burn patients (61). Unfortunately, the possible effect of OKG on collagen synthesis has, to our knowledge, never been studied (L Cynober, Paris Descartes University Hospital, personal communication, 2008).
Overall, there are a number of hypotheses regarding the augmentation of wound healing; however, the best method of supporting collagen biosynthesis is by providing adequate host nutrition. Arginine and ornithine supplementation are most effective in increasing collagen deposition, but whether this is accomplished by conversion to proline or other biochemical pathways remains to be confirmed. The failure of citric acid precursors and amino acids to improve collagen synthesis remains puzzling and deserves further study, but the lack of effect is consistent with the complex regulation of cellular metabolism.
Acknowledgments
The authors’ responsibilities were as follows—VLA and KM: drafted the first version of the manuscript; and all authors: contributed critical revisions, edited the manuscript, and read and approved the final version of the manuscript.
Footnotes
Abbreviations used: GSA, glutamate-5-semialdehyde; OAT, ornithine aminotransferase; OKG, ornithine-α-ketoglutarate; P5C, pyrroline-5-carboxylate.
References
- 1.Park JE, Barbul A. Understanding the role of immune regulation in wound healing. Am J Surg 2004;187:11S–16S. [DOI] [PubMed] [Google Scholar]
- 2.Witte MB, Barbul A. General principles of wound healing. Surg Clin North Am 1997;77:509–28. [DOI] [PubMed] [Google Scholar]
- 3.Thomas DR. Improving outcome of pressure ulcers with nutritional interventions: a review of the evidence. Nutrition 2001;17:121–5. [DOI] [PubMed] [Google Scholar]
- 4.Arnold M, Barbul A. Nutrition and wound healing. Plast Reconstr Surg 2006;117:42S–58S. [DOI] [PubMed] [Google Scholar]
- 5.Barbul A. Proline precursors to sustain mammalian collagen synthesis. J Nutr 2008;138:2021S–4S. [DOI] [PubMed] [Google Scholar]
- 6.Krane SM. The importance of proline residues in the structure, stability and susceptibility to proteolytic degradation of collagens. Amino Acids 2008;35:703–10. [DOI] [PubMed] [Google Scholar]
- 7.Grant ME, Prockop DJ. The biosynthesis of collagen. 3. N Engl J Med 1972;286:291–300. [DOI] [PubMed] [Google Scholar]
- 8.Fukushima R, Yamazaki E. Vitamin C requirement in surgical patients. Curr Opin Clin Nutr Metab Care 2010;13:669–76. [DOI] [PubMed] [Google Scholar]
- 9.Trent JT, Kirsner RS. Leg ulcers secondary to prolidase deficiency. Adv Skin Wound Care 2004;17:468–72. [DOI] [PubMed] [Google Scholar]
- 10.Wu G. Intestinal mucosal amino acid catabolism. J Nutr 1998;128:1249–52. [DOI] [PubMed] [Google Scholar]
- 11.Wu G, Bazer FW, Datta S, Johnson GA, Li P, Satterfield MC, Spencer TE. Proline metabolism in the conceptus: implications for fetal growth and development. Amino Acids 2008;35:691–702. [DOI] [PubMed] [Google Scholar]
- 12.Wu G, Bazer FW, Davis TA, Kim SW, Li P, Rhoads JM, Satterfield MC, Smith SB, Spencer TE, Yin Y. Arginine metabolism and nutrition in growth, health and disease. Amino Acids 2009;37:153–68. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 13.Ginguay A, Cynober L, Curis E, Nicolis I. Ornithine aminotransferase, an important glutamate-metabolizing enzyme at the crossroads of multiple metabolic pathways. Biology (Basel) 2017;6:18. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 14.Windmueller HG, Spaeth AE. Uptake and metabolism of plasma glutamine by the small intestine. J Biol Chem 1974;249:5070–9. [PubMed] [Google Scholar]
- 15.Reeds PJ, Burrin DG, Stoll B, Jahoor F. Intestinal glutamate metabolism. J Nutr 2000;130:978S–82S. [DOI] [PubMed] [Google Scholar]
- 16.Matthews DE, Marano MA, Campbell RG. Splanchnic bed utilization of glutamine and glutamic acid in humans. Am J Physiol 1993;264:E848–54. [DOI] [PubMed] [Google Scholar]
- 17.Windmueller HG, Spaeth AE. Intestinal metabolism of glutamine and glutamate from the lumen as compared to glutamine from blood. Arch Biochem Biophys 1975;171:662–72. [DOI] [PubMed] [Google Scholar]
- 18.Windmueller HG, Spaeth AE. Metabolism of absorbed aspartate, asparagine, and arginine by rat small intestine in vivo. Arch Biochem Biophys 1976;175:670–6. [DOI] [PubMed] [Google Scholar]
- 19.Stoll B, Burrin DG, Henry J, Yu H, Jahoor F, Reeds PJ. Substrate oxidation by the portal drained viscera of fed piglets. Am J Physiol 1999;277:E168–75. [DOI] [PubMed] [Google Scholar]
- 20.Stoll B, Henry J, Reeds PJ, Yu H, Jahoor F, Burrin DG. Catabolism dominates the first-pass intestinal metabolism of dietary essential amino acids in milk protein-fed piglets. J Nutr 1998;128:606–14. [DOI] [PubMed] [Google Scholar]
- 21.Murphy JM, Murch SJ, Ball RO. Proline is synthesized from glutamate during intragastric infusion but not during intravenous infusion in neonatal piglets. J Nutr 1996;126:878–86. [DOI] [PubMed] [Google Scholar]
- 22.Tomlinson C, Rafii M, Sgro M, Ball RO, Pencharz P. Arginine is synthesized from proline, not glutamate, in enterally fed human preterm neonates. Pediatr Res 2011;69:46–50. [DOI] [PubMed] [Google Scholar]
- 23.Tomlinson C, Rafii M, Ball RO, Pencharz PB. Arginine can be synthesized from enteral proline in healthy adult humans. J Nutr 2011;141:1432–6. [DOI] [PubMed] [Google Scholar]
- 24.Köhler ES, Sankaranarayanan S, van Ginneken CJ, van Dijk P, Vermeulen JL, Ruijter JM, Lamers WH, Bruder E. The human neonatal small intestine has the potential for arginine synthesis; developmental changes in the expression of arginine-synthesizing and -catabolizing enzymes. BMC Dev Biol 2008;8:107. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 25.Windmueller HG, Spaeth AE. Source and fate of circulating citrulline. Am J Physiol 1981;241:E473–80. [DOI] [PubMed] [Google Scholar]
- 26.Bertolo RF, Burrin DG. Comparative aspects of tissue glutamine and proline metabolism. J Nutr 2008;138:2032S–9S. [DOI] [PubMed] [Google Scholar]
- 27.Dhanakoti SN, Brosnan JT, Herzberg GR, Brosnan ME. Renal arginine synthesis: studies in vitro and in vivo. Am J Physiol 1990;259:E437–42. [DOI] [PubMed] [Google Scholar]
- 28.Cynober L, Le Boucher J, Vasson M-P. Arginine metabolism in mammals. J Nutr Biochem 1995;6:402–13. [Google Scholar]
- 29.Lorendeau D, Christen S, Rinaldi G, Fendt SM. Metabolic control of signalling pathways and metabolic auto‐regulation. Biol Cell 2015;107:251–72. [DOI] [PubMed] [Google Scholar]
- 30.Caldwell MD, Mastrofrancesco B, Shearer J, Bereiter D. The temporal change in amino acid concentration within wound fluid—a putative rationale. Prog Clin Biol Res 1991;365:205–22. [PubMed] [Google Scholar]
- 31.Peacock EE., Jr Effect of dietary proline and hydroxyproline on tensile strength of healing wounds. Proc Soc Exp Biol Med 1960;105:380–3. [DOI] [PubMed] [Google Scholar]
- 32.Tapiero H, Mathé G, Couvreur P, Tew KD. II. Glutamine and glutamate. Biomed Pharmacother 2002;56:446–57. [DOI] [PubMed] [Google Scholar]
- 33.Askanazi J, Carpentier YA, Michelsen CB, Elwyn DH, Furst P, Kantrowitz LR, Gump FE, Kinney JM. Muscle and plasma amino acids following injury. Influence of intercurrent infection. Ann Surg 1980;192:78–85. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 34.Bellon G, Chaqour B, Wegrowski Y, Monboisse J-C, Borel J-P. Glutamine increases collagen gene transcription in cultured human fibroblasts. Biochim Biophys Acta 1995;1268:311–23. [DOI] [PubMed] [Google Scholar]
- 35.Karna E, Miltyk W, Wołczyński S, Pałka JA. The potential mechanism for glutamine-induced collagen biosynthesis in cultured human skin fibroblasts. Comp Biochem Physiol B Biochem Mol Biol 2001;130:23–32. [DOI] [PubMed] [Google Scholar]
- 36.Bozkırlı BO, Gündoğdu RH, Ersoy E, Lortlar N, Yıldırım Z, Temel H, Oduncu M, Karakaya J. Pilot experimental study on the effect of arginine, glutamine, and β-hydroxy β-methylbutyrate on secondary wound healing. JPEN J Parenter Enteral Nutr 2015;39:591–7. [DOI] [PubMed] [Google Scholar]
- 37.Sanli A, Onen A, Sarioglu S, Sis B, Guneli E, Gokcen B, Karapolat S, Acikel U. Glutamine administration enhances the healing of lung parenchymal injuries and reduces air leakage in rats. Tohoku J Exp Med 2006;210:239–45. [DOI] [PubMed] [Google Scholar]
- 38.da Costa MA, Campos AC, Coelho JC, de Barros AM, Matsumoto HM. Oral glutamine and the healing of colonic anastomoses in rats. JPEN J Parenter Enteral Nutr 2003;27:182–5. [DOI] [PubMed] [Google Scholar]
- 39.Alecrim HM, Duarte SAC, Amaral MEB, Diógenis F, Carneiro FP, de Sousa JB. Effect of glutamine supplementation on left colon healing in rats with extrahepatic biliary obstruction. Acta Cir Bras 2015;30:73–9. [DOI] [PubMed] [Google Scholar]
- 40.Zhou YP, Jiang ZM, Sun YH, Wang XR, Ma EL, Wilmore D. The effect of supplemental enteral glutamine on plasma levels, gut function, and outcome in severe burns: a randomized, double-blind, controlled clinical trial. JPEN J Parenter Enteral Nutr 2003;27:241–5. [DOI] [PubMed] [Google Scholar]
- 41.Rocha BR, Gombar FM, Barcellos LM, Costa WS, Barcellos Sampaio FJ, Ramos CF. Glutamine supplementation prevents collagen expression damage in healthy urinary bladder caused by radiotherapy. Nutrition 2011;27:809–15. [DOI] [PubMed] [Google Scholar]
- 42.Murakami H, Shimbo K, Inoue Y, Takino Y, Kobayashi H. Importance of amino acid composition to improve skin collagen protein synthesis rates in UV-irradiated mice. Amino Acids 2012;42:2481–9. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 43.Williams JZ, Abumrad N, Barbul A. Effect of a specialized amino acid mixture on human collagen deposition. Ann Surg 2002;236:369–75. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 44.Murakami H, Shimbo K, Takino Y, Kobayashi H. Combination of BCAAs and glutamine enhances dermal collagen protein synthesis in protein-malnourished rats. Amino Acids 2013;44:969–76. [DOI] [PubMed] [Google Scholar]
- 45.Gündoğdu RH, Temel H, Bozkırlı BO, Ersoy E, Yazgan A, Yıldırım Z. The mixture of arginine, glutamine, and β-hydroxy-β-methyl butyrate enhances the healing of ischemic wounds in rats. JPEN J Parenter Enteral Nutr 2016. Jan 29 (Epub ahead of print; DOI: 10.1177/0148607115625221). [DOI] [PubMed] [Google Scholar]
- 46.Jones MS, Rivera M, Puccinelli CL, Wang MY, Williams SJ, Barber AE. Targeted amino acid supplementation in diabetic foot wounds: pilot data and a review of the literature. Surg Infect (Larchmt) 2014;15:708–12. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 47.Kesici U, Kesici S, Ulusoy H, Yucesan F, Turkmen AU, Besir A, Tuna V. Effects of glutamine on wound healing. Int Wound J 2015;12:280–4. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 48.Barbul A. Arginine: biochemistry, physiology, and therapeutic implications. JPEN J Parenter Enteral Nutr 1986;10:227–38. [DOI] [PubMed] [Google Scholar]
- 49.Seifter E, Rettura G, Barbul A, Levenson SM. Arginine: an essential amino acid for injured rats. Surgery 1978;84:224–30. [PubMed] [Google Scholar]
- 50.Barbul A, Sisto DA, Wasserkrug HL, Yoshimura NN, Efron G. Metabolic and immune effects of arginine in postinjury hyperalimentation. J Trauma 1981;21:970–4. [DOI] [PubMed] [Google Scholar]
- 51.Barbul A, Lazarou SA, Efron DT, Wasserkrug HL, Efron G. Arginine enhances wound healing and lymphocyte immune responses in humans. Surgery 1990;108:331–7. [PubMed] [Google Scholar]
- 52.Kirk SJ, Hurson M, Regan MC, Holt DR, Wasserkrug HL, Barbul A. Arginine stimulates wound healing and immune function in elderly human beings. Surgery 1993;114:155–60. [PubMed] [Google Scholar]
- 53.Albina JE, Mills CD, Barbul A, Thirkill CE, Henry WL, Mastrofrancesco B, Caldwell MD. Arginine metabolism in wounds. Am J Physiol 1988;254:E459–67. [DOI] [PubMed] [Google Scholar]
- 54.Shi HP, Fishel RS, Efron DT, Williams JZ, Fishel MH, Barbul A. Effect of supplemental ornithine on wound healing. J Surg Res 2002;106:299–302. [DOI] [PubMed] [Google Scholar]
- 55.Shi HP, Efron DT, Most D, Tantry US, Barbul A. Supplemental dietary arginine enhances wound healing in normal but not inducible nitric oxide synthase knockout mice. Surgery 2000;128:374–8. [DOI] [PubMed] [Google Scholar]
- 56.Smith RJ, Phang JM. Proline metabolism in cartilage: the importance of proline biosynthesis. Metabolism 1978;27:685–94. [DOI] [PubMed] [Google Scholar]
- 57.Smith RJ, Phang JM. The importance of ornithine as a precursor for proline in mammalian cells. J Cell Physiol 1979;98:475–81. [DOI] [PubMed] [Google Scholar]
- 58.Curis E, Nicolis I, Moinard C, Osowska S, Zerrouk N, Bénazeth S, Cynober L. Almost all about citrulline in mammals. Amino Acids 2005;29:177–205. [DOI] [PubMed] [Google Scholar]
- 59.Osowska S, Moinard C, Loï C, Neveux N, Cynober L. Citrulline increases arginine pools and restores nitrogen balance after massive intestinal resection. Gut 2004;53:1781–6. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 60.Rizk M, Witte MB, Barbul A. Nitric oxide and wound healing. World J Surg 2004;28:301–6. [DOI] [PubMed] [Google Scholar]
- 61.Cynober LA. Metabolic & therapeutic aspects of amino acids in clinical nutrition. 2nd ed. Boca Raton, FL: CRC Press; 2003. [Google Scholar]